
Abstract
This study aims to explore the mechanism underlying the role of ubiquitin-specific protease 14 (USP14) in regulating P53 expression and influencing the development of hepatitis B. The animal and cell models of hepatitis B were constructed. The mRNA and protein expression of USP14, mouse double minute 2 (MDM2), and P53 were detected by western blot and qPCR. The USP14 overexpression vector was constructed. The pathological changes of liver tissue were detected by HE and Masson staining. Protein immunoprecipitation was used to detect the interaction between MDM2 and P53, as well as between MDM2 and USP14. The ubiquitination levels of P53 after USP14 overexpression were detected. qPCR and western blot were used to detect the expression of MDM2, Bcl-2, P53, Bax, and Caspase-1 in vivo and in vitro. Compared with the control group, the model group showed increased cell proliferation, increased expression of MDM2 and Bcl-2 in cells and liver tissue, and decreased expression of P53, Bax, and Caspase-1. Compared with the model group, overexpression of USP14 resulted in a decrease in MDM2 expression and an increase in P53 expression. After transfection with the USP14 overexpression plasmid, cell proliferation was inhibited, and the expression of MDM2 and Bcl-2 was decreased in cells and liver tissue, while the expression of P53, Bax, and Caspase-1 was increased. In the model of hepatitis B, USP14 upregulation downregulated MDM2 and promoted P53 deubiquitination to delay the invasion of hepatitis B virus and the development of hepatitis.
Background
As a small DNA virus, hepatitis B virus (HBV) is closely related to diverse liver disorders, such as liver failure, hepatocellular carcinoma (HCC), and cirrhosis (Seeger and Mason, 2015). Although most adults could spontaneously clear HBV after acute infection, about 5% of people develop chronic infection (Zou et al., 2021). Therefore, the development of new rapid screening methods for patients with hepatitis B and the exploration of the mechanism underlying HBV’s role in cirrhosis and liver cancer development have important social and economic benefits for the prevention and treatment of hepatitis B.
HBV is associated with most pathogen-related tumors (Bouvard et al., 2009), and the tumor viruses exert their carcinogenic effects primarily by disrupting the P53 signaling pathway (Tornesello et al., 2015). They can hinder the expression of tumor suppressors, such as p130/Rb, p107, pRb, and P53, and enable cells to evade apoptosis (Sáenz-Robles et al., 2001). HBx, consisting of 154 amino acids, is a protein that contributes to viral replication and HBV transcription (McClain et al., 2007). Extensive research has shown complicated interactions between P53 and HBx. HBx can bind to the c-terminus of the P53 protein and hinder the function of P53 to regulate downstream gene expression (inhibiting P53 activity) (Soria et al., 2010). Mouse double minute 2 (MDM2) is an oncogenic gene that facilitates cancer cells to proliferate and migrate. Nonphosphorylated P53 binds to MDM2, which inhibits P53. In addition, MDM2 has E3 ligase activity, which can cause ubiquitination and degradation of P53 protein. When P53 phosphorylates, it dissociates from the MDM2/MDX complex, activates and regulates the transcription of downstream Bax and other apoptotic-related genes, and promotes cancer cell apoptosis (Brooks et al., 2007). Xian et al. found that overexpression of P53 leads to the degradation of HBx in HBV-infected liver cancer cells, while overexpression of MDM2 promotes the accumulation of HBx in cells (Xian et al., 2010).
P53 degradation is primarily controlled by the ubiquitin-proteasome pathway (Lee and Gu, 2010). USP7, USP5, USP13, USP10, and other deubiquitinating enzymes enhance P53 stability through removing ubiquitin molecules (Dayal et al., 2009; Li et al., 2002; Liu et al., 2011). Ubiquitin-specific protease 14 (USP14), belonging to the USP protein family (Komander et al., 2009), acts as a potential deubiquitinase of P53 (Zhou et al., 2019). It can enhance deubiquitination and prevent MDM2-mediated degradation of the ubiquitin-proteasome pathway (Muniyappan and Lee, 2019).
HBV infection is a major global health issue, particularly in its role in the development of liver cirrhosis and HCC. While previous research has highlighted the viral mechanisms that influence liver pathology, our study uniquely explores the regulatory role of USP14 in modulating the ubiquitination and degradation of the tumor suppressor protein P53 in the context of HBV infection (Ma et al., 2020). This is the first study to systematically investigate how USP14 stabilizes P53 through deubiquitination, thereby delaying HBV invasion and progression of liver disease (Liu et al., 2019). Unlike prior studies that focus on HBx interactions or general P53 dynamics, we provide novel insights into the USP14–P53 interaction as a potential therapeutic target (Elmore et al., 1997; Lim et al., 2022). Our findings offer significant advancements in both the molecular understanding of HBV pathology and the development of targeted interventions to manage chronic HBV infection.
Objectives
While ubiquitination is well studied, the specific role of USP14 in regulating P53 during HBV-induced liver cirrhosis and cancer remains underexplored. This study aims to fill this gap by demonstrating the crucial function of USP14 in stabilizing P53 and its potential as a novel therapeutic target for HBV treatment.
Materials and Methods
Study design
Cell culture and transfection
WRL68 cells (Bio 105797, Biobw, Beijing, China) were cultured in a complete MEM medium (KGM41500S Keji Bio), supplemented with 10% fetal bovine serum (FBS, 10099-141, Gibco, USA), and incubated at 37°C with 95% air and 5% CO2. Cells were passaged every 48 h upon reaching 80–90% confluence. For transfection, LipofectamineTM 3000 (L300015 InvitrogenTM, USA) was used following the manufacturer’s protocol. Cells were transfected in a serum-free medium for 6 h, followed by the addition of complete MEM containing 20% serum. Transfection efficiency was assessed 48 h post-transfection via qPCR. Cells were collected at 24, 48, and 72 h for downstream analysis.
Vector construction
The USP14 gene (GenBank accession number: NM_005151.4) was synthesized using cDNA derived from human liver tissue. The USP14 coding sequence was amplified and inserted into the pcDNA3.1(+) vector (BamHI/EcoRI sites) using T4 DNA ligase. The recombinant plasmid was transformed into DH5α competent cells via heat shock. Following transformation, bacterial colonies were selected on LB agar plates containing ampicillin (100 µg/mL), and positive clones were further cultured in LB broth. The plasmids were purified using the Qiagen Plasmid Mini Kit and verified by restriction enzyme digestion (BamHI/EcoRI) and Sanger sequencing.
Setting
CCK8 detection
After cell transfection for 24 h, each well was provided with CCK8 reagent (10 µL, C0071S, Beyotime) for 2 h of incubation at 37°C. Each well’s OD value at a wavelength of 450 nm was assessed via a multifunctional enzyme labeling analyzer (SuPerMax3100, Shanghai Shanpu Biotechnology Co., Ltd.). Last, the cell survival rate was calculated.
Flow cytometry
First, 2× EdU working fluid (20 μM) was put to the 6-well plate to achieve a final EdU concentration of 1×. Incubation of the cells for 2 h was followed by digestion and collection for fixation. Each well was offered a Click reaction solution (0.5 mL), followed by a 30-min incubation at ambient temperature in the dark and subsequent centrifugation to remove the Click reaction solution. After washing for 3 times, the cells were suspended in PBS and subjected to detection. Upon resuspension with 300 μL PBS, the cell samples were analyzed with a FACSCanto™ II flow cytometer (BD Biosciences).
qPCR detection
RNA extraction was performed, and the UV visible spectrophotometer (OD260/OD280) was utilized to determine the mRNA concentration and purity. The cDNA synthesis was carried out with mRNA reverse transcription kits, and the qPCR was conducted utilizing a fluorescence PCR instrument [CFX ConnecTM real-time, BioRad Life Medical Products (Shanghai) Co., Ltd.]. The PCR setup encompassed: cDNA 1 μL, upstream primer 0.4 μL, 2 × SYBR Green PCR Master Mix 10 μL, RNase-Free dH2O 8.2 μL, and downstream primer 0.4 μL. The primers were shown in Table 1. The following were the steps of the reaction: predenaturation at 95°C for 10 min; denaturation at 95°C for 10 sec, annealing at 58°C for 30 sec, extend at 72°C for 30 sec, 40 cycles. The genes’ relative expression levels were computed through the 2−△△Ct method, with β-actin serving as the internal reference.
Primers Used in This Study
Western blot
The samples were lysed using cell lysis buffer to extract total protein, which was then stored at −20°C. A BCA kit was recruited to test the protein concentration. After denaturation, the protein underwent sodium dodecylbenzene sulfonate gel electrophoresis (SDS-PAGE) for 1.5 h, followed by membrane transfer using 300 mA constant flow for 1.5 h. Incubation of the PVDF membrane was performed based on the primary antibodies including Mouse Anti β-Actin (HC201, TransGen Biotech, 1/2000), Rabbit Anti-MDM2 (AF0208, Affinity, 1/1000), Rabbit Anti-P53 (10442–1-AP, Proteintech, 1/2000), Rabbit Anti-USP14 (sc-515812, Santa Cruz, 1/1000), Rabbit Anti-USP4 (2651S, CST, 1/1000); Rabbit Anti-USP411 (DF4586, Affinity, 1/1000); Mouse Anti-GAPDH (TA-08, ZSGB-BIO, 1/2000) overnight at 4°C, followed by incubation with the Goat Anti-Rabbit IgG (H + L) HRP (S0001, affinity, 1/2000), HRP-conjugated Goat Anti-Mouse IgG (H + L) (GB23301, Servicebio, 1/2000) at ambient temperature for 2 h. Upon rinsing, the luminescent liquid was added, and the membrane was placed in the sample placement area of the ultra-high-sensitivity chemiluminescence imaging system [Chemi DocTM XRS+, BioRad Life Medical Products (Shanghai) Co., Ltd.] for imaging.
Protein immunoprecipitation
IP cell lysis buffer (RQ-IP01, Zhongzheng) was used to lyse the sample at 4°C. Following 30 min, samples underwent centrifugation at 4°C, 16,000 g. Upon 15 min, 50 μL of supernatant was collected as input, and another 400 μL of supernatant was put in a new 1.5 mL EP tube for Co-IP experiment. Then, 2–5 μg of target antibodies Rabbit Anti-MDM2 (AF0208, Affinity, 1/1000) and Rabbit Anti-P53 (10442-1-AP, Proteintech, 1/2000) were offered to the IP sample and subjected to an overnight of incubation at 4°C for rotation. Next, 40 μL Protein G Plus/Protein A Agarose Suspension (Millipore, IP05) was supplemented to the IP sample and incubated with a vertical mixer at 4°C for 3 h. Following centrifuging at 4000 rpm for 1 min at 4°C, the supernatant was removed. The pellet was subsequently washed 5 times with IP washing solution 1, centrifuged at 4000 rpm for 1 min at 4°C, succeeded by removal of the supernatant. Upon washing once utilizing IP washing solution 2, the samples were centrifuged at 4000 rpm at 4°C for 1 min, and the supernatant was eliminated. Last, the samples were given 80 μL of IP lysis buffer and loading buffer, mixed well, and boiled in boiling water for 10 min.
Participants
Animal modeling and treatment
We used 6- to 8-week-old female BALB/c mice provided by Beijing Huafu Biotechnology Co., Ltd. [License number: SCXK (Beijing) 2019–0008]. The mice were housed in a controlled environment with a temperature of 20–26°C and relative humidity of 40–70%, under a 12-h light-dark cycle, with ad libitum access to food and water.
The mice were anesthetized using pentobarbital sodium (50 mg/kg, intraperitoneal injection), and the depth of anesthesia was confirmed by the absence of the pedal reflex. The mice were placed on a warming pad during the procedure. The abdominal area was shaved and disinfected with iodine solution, and a midline incision was made to expose the hepatic portal vein. In the model+empty vector group, 50 μL of the virus was injected into the liver via the hepatic portal vein using a 32G insulin needle. Similarly, in the model+overexpression group, 50 μL of overexpression virus was injected. Hemostasis was achieved, and the abdomen was closed with sutures. The mice were kept on an insulation pad during recovery from anesthesia. Approval for this study was acquired from the Ethics Committee of Guangxi Medical University Cancer Hospital (LW2023155).
A mouse HBV model was constructed by injecting 15 μg of HBV gene DNA replication plasmid into the tail vein. After anesthesia with pentobarbital sodium, the mice were shaved on the abdomen, disinfected, and opened, and the hepatic portal vein was exposed. In the model+empty vector group, 50 μL of empty virus was extracted using a 32G insulin needle and injected into the liver through the hepatic portal vein. After pressing to stop bleeding, the abdomen was closed and sutured, and an insulation pad was placed to keep warm. The model+overexpression group used the same method to extract 50 μL of overexpression virus and inject it into the liver through the hepatic portal vein, while the control group only exposed the hepatic portal vein. After 4 days of adenovirus intervention, a tail vein injection of 15 μg of HBV was performed to induce the model. After 4 days of injection of 15 μg of HBV, samples were taken for subsequent testing.
H&E staining
Running water was adopted for rinsing the tissue for several hours. After dehydration with 70%, 80%, and 90% ethanol solutions, the samples were incubated with the mixture of xylene and pure alcohol for 15 min, xylene I and II for 15 min (until transparent). Then, the tissue incubation was performed with a mixture of paraffin and xylene for 15 min, paraffin I and paraffin II for 50–60 min each. Then, paraffin embedding, slicing, grilling, dewaxing, and hydration were performed. The tissue was stained with hematoxylin aqueous solvent for 3 min. Upon differentiation in hydrochloric acid ethanol differentiation solution for 15 sec, the tissue was washed marginally via water, returned to blue with blue solution for 15 sec, cleaned through running water, stained utilizing eosin for 3 min, bathed by running water, dehydrated, cleared, and sealed. Finally, a microscopic examination was performed.
Masson staining
After dewaxing, the paraffin slices were immersed in Masson A solution (G1006, Servicebio) at room temperature overnight (about 15 h). Soak the slices in Masson A solution and incubate in a 65°C oven for 30 min. Wash with tap water for 30 sec until the yellow color on the tissue fades away. Mix Masson B solution (G1006, Servicebio) with Masson C solution (G1006, Servicebio) in equal volume. Immerse the slices in the mixed solution for 1 min and wash slightly with running water. The slice differentiation was performed utilizing 1% hydrochloric acid alcohol until the nucleus was grayish black and the background was almost colorless or light gray. Wash with tap water, immerse the slices in Masson D solution (G1006, Servicebio) for 6 min, and immerse them in Masson E solution (G1006, Servicebio) for about 1 min. Differentiate until the collagen fibers are light red and the fibers are red. Directly stain with Masson F solution (G1006, Servicebio) for 2–30 sec. The slices were subjected to differentiation with 1% acetic acid, followed by dehydration with anhydrous ethanol and n-butanol. Finally, xylene was used for transparency, and the slices were sealed with neutral gum.
Statistical analysis
Statistical analysis was conducted with SPSS 20.0 software. Each experiment was reiterated 3 times, with the quantitative outcomes presented as mean ± standard deviation. The quantitative values between the two groups were compared using independent sample t-tests. The quantitative values between multiple groups were compared using one-way analysis of variance. The S-N-K method was exercised for pairwise comparisons. The significance level was set at α = 0.05.
Results
USP14 was low expressed in the model group, and the USP14 expression trend was consistent with P53
For studying USP14’s in the development of hepatitis B, WRL68 cells were infected with HBV DNA plasmid to establish a cell model, and the USP14, MDM2, and P53 mRNA and protein expression levels were analyzed. Relative to the control group, the qPCR results demonstrated a significant increase in HBX expression in the model group, indicating that the in vitro model of hepatitis B was established successfully (Fig. 1A) (p < 0.05). As against the control group, the mRNA expression of USP14 and P53 was notably decreased (Fig. 1B) (p < 0.05). According to the western blot results, the model group exhibited markedly lower levels of USP14 and P53 and considerably higher levels of MDM2 compared to the control group (Fig. 1C) (p < 0.05). Hence, USP14 may affect the progression of HBV-related liver cirrhosis/HCC by affecting the expression of P53.
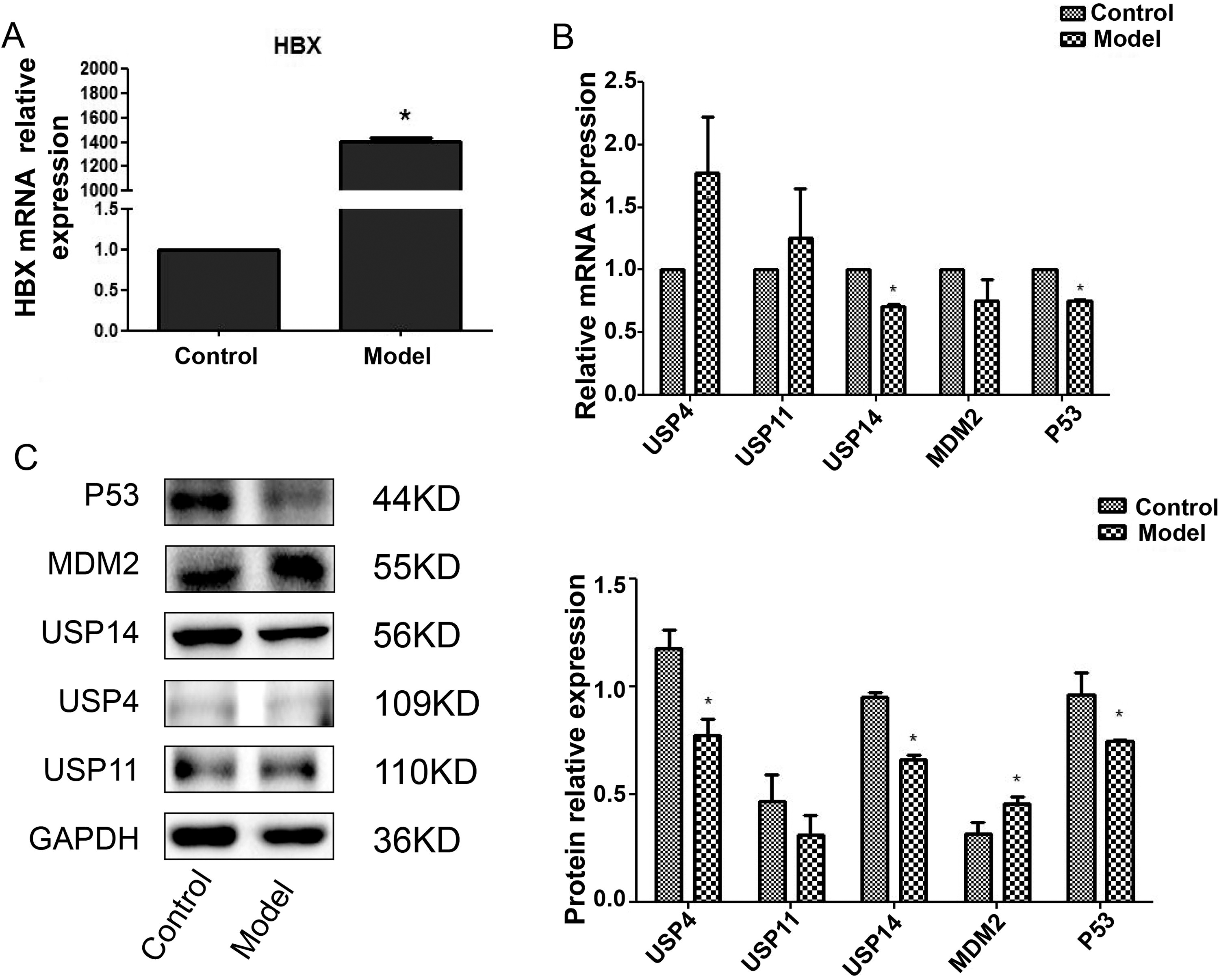
USP14 was low expressed in the model group, and the expression trend of USP14 was consistent with P53.
Effect of USP14 overexpression on proliferation and apoptosis of model group
For further exploring USP14’s functions in hepatitis B development, USP14 overexpression vector was constructed. The USP14 mRNA expression was increased remarkably following transfection of USP14 plasmid (p < 0.05) (Fig. 2A), indicating that the construction of the overexpression vector was successful.
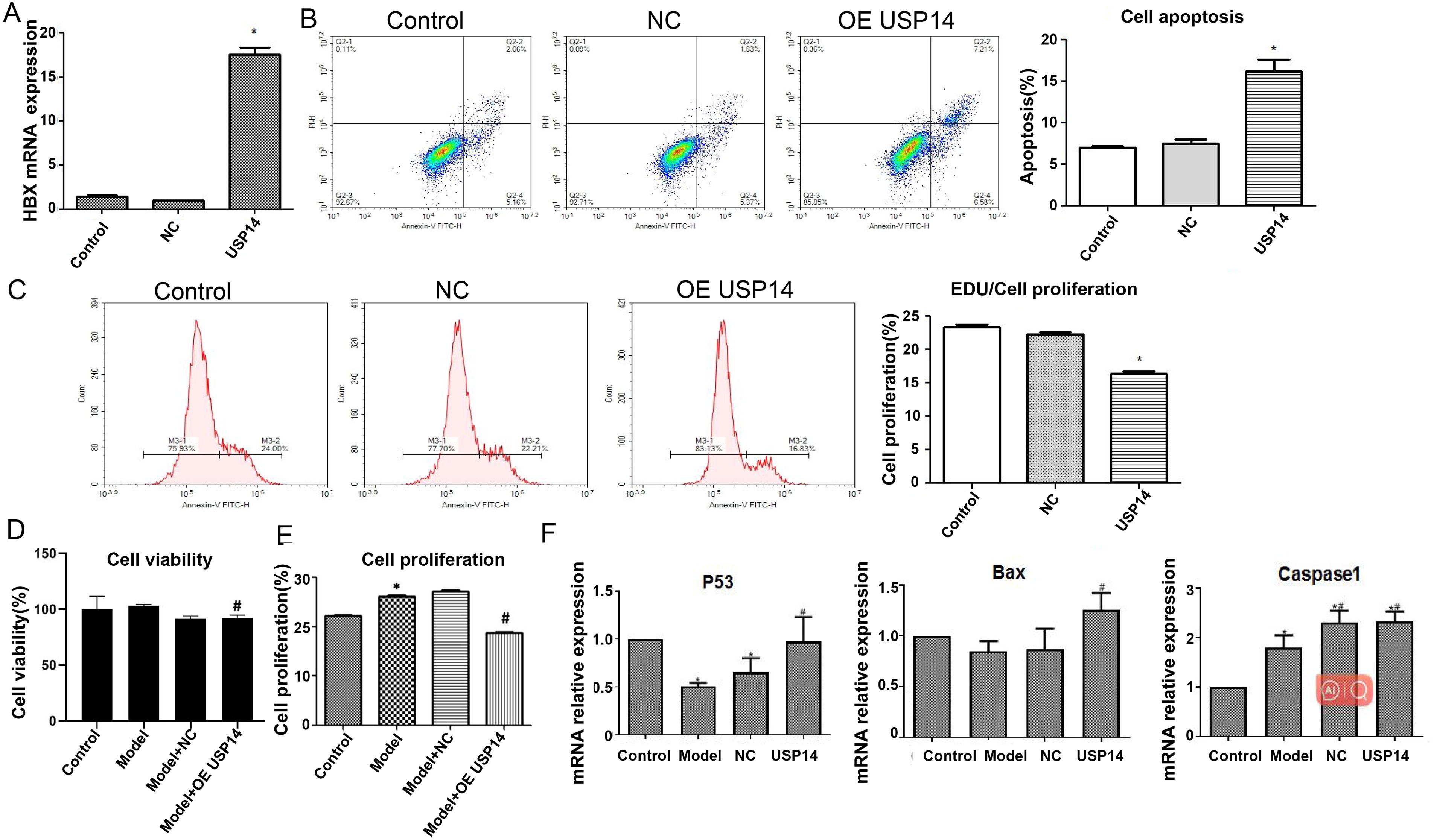
Effect of USP14 overexpression on proliferation and apoptosis of model group.
Subsequently, the effect of USP14 on cell proliferation and apoptosis was examined. As indicated by flow cytometry analysis, contrasted with the control group, USP14 overexpression elevated cell apoptosis rate. The Edu proliferation results showed a decrease in cell proliferation rate (Fig. 2B, C) (p < 0.05).
After transfection with the HBx plasmid, the cell viability was slightly increased with no statistical significance. After overexpression of USP14, the cell viability was dropped, with the difference being statistically significant (Fig. 2D) (p < 0.05). The Edu proliferation outcomes disclosed that in relation to the controls, the transfection of HBx plasmid significantly enhanced the cell proliferation ability, while USP14 overexpression decreased the cell proliferation ability, with a difference of statistically significant (Fig. 2E) (p < 0.05).
Figure 2F showed that P53 mRNA expression was reduced in the model group compared to the control group. Upon overexpression of USP14, the P53, Bax, and Caspase-1 mRNA expression was increased, with a statistically significant difference (p < 0.05). The observation outcomes indicated that HBV infection could hinder liver cancer cells’ apoptosis, while overexpression of USP14 increased apoptosis.
USP14 maintained P53 stability through its deubiquitination activity
To research the correlation of USP14 and MDM2 with P53, protein immunoprecipitation detection was performed. As shown in Figure 3A, MDM2 and USP14 proteins had clear target bands in the input. IP reactions were performed using MDM2 (IP) antibody, and MDM2 control and model showed clear bands in the IP product; however, MDM2 was not detected in the IgG sample. The results indicate that USP14 and MDM2 interact with each other.
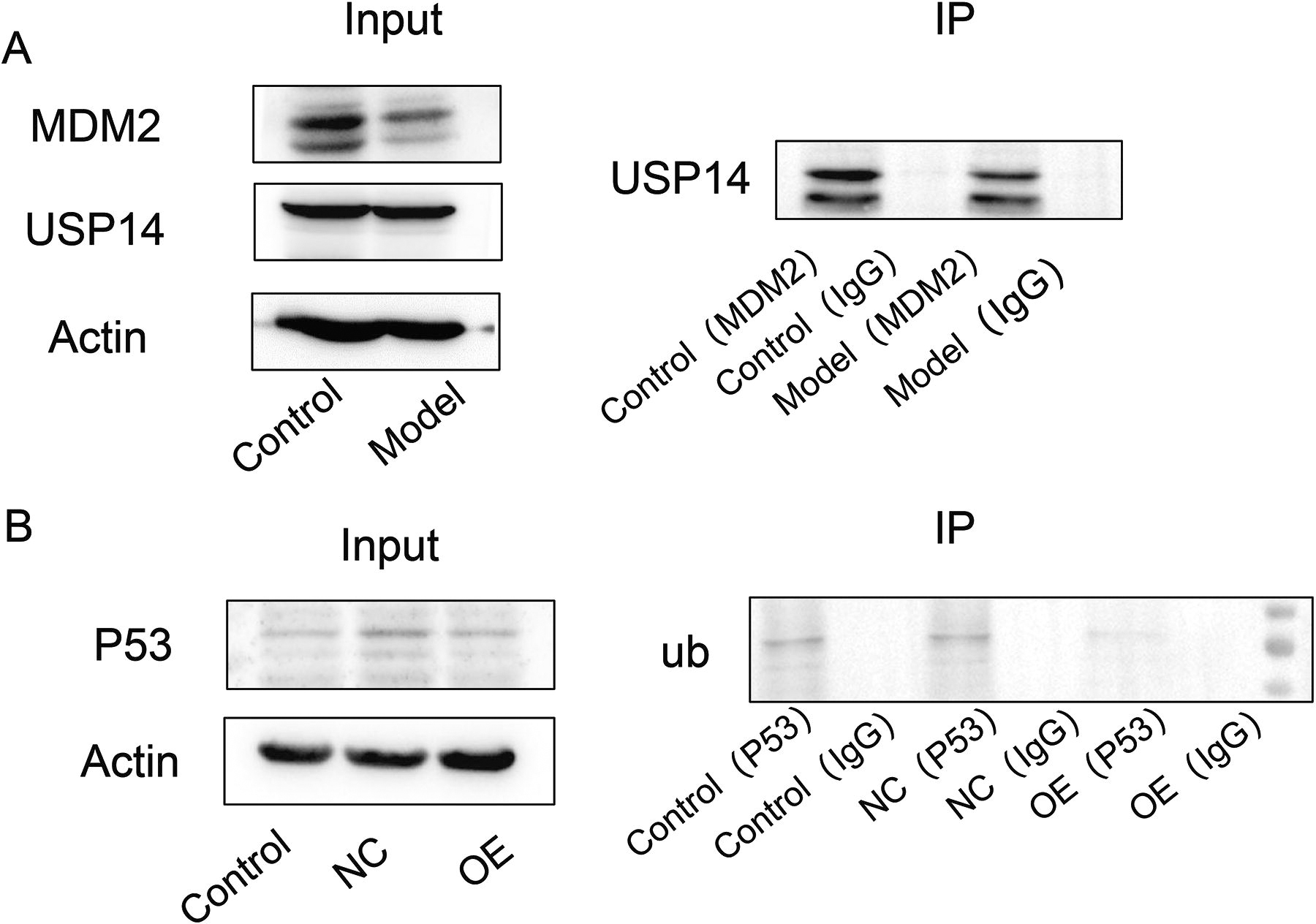
USP14 maintained P53 stability through its deubiquitination activity.
As shown in Figure 3B, ubiquitination bands were detected in the IP products of both the control and NC groups using the P53 (IP) antibody for IP reaction. However, after overexpression of USP14, no expression of ub was detected in the IP products, indicating that USP14 inhibits the ubiquitination of P53 (causing it to deubiquitination).
Overexpression of USP14 reduced HBx-induced hepatitis B in vivo
A mouse HBV model was constructed by injecting 15 ug of HBV gene DNA replication plasmid into the tail vein. Four days before modeling, USP14 overexpression adenovirus and empty adenovirus were injected into the hepatic portal vein to verify whether USP14 plays a key role in hepatitis B. After injecting HBV plasmid into the tail vein for 4 days, liver tissue was taken for testing. As shown in Figure 4A, the model group exhibited substantially increased HBx gene expression in liver tissue in contrary to the control group (p < 0.05). The HE outcomes displayed that the liver cell structure in the model group was severely damaged with nuclear pyknosis. Overexpression of USP14 alleviated liver cell damage (Fig. 4B). Masson’s results showed an increase in fibrosis in the model group, and after overexpression of USP14, the fibrosis situation was alleviated (Fig. 4B). qPCR analysis revealed that as against the controls, the MDM2 and Bcl-2 expression was increased in the model group, while the P53, Bax, and Caspase-1 expression was decreased. After overexpression of USP14, the expression of MDM2 and Bcl-2 was diminished, while the P53, Bax, and Caspase-1 expression was increased, with statistical significance (Fig. 4C) (p < 0.05).
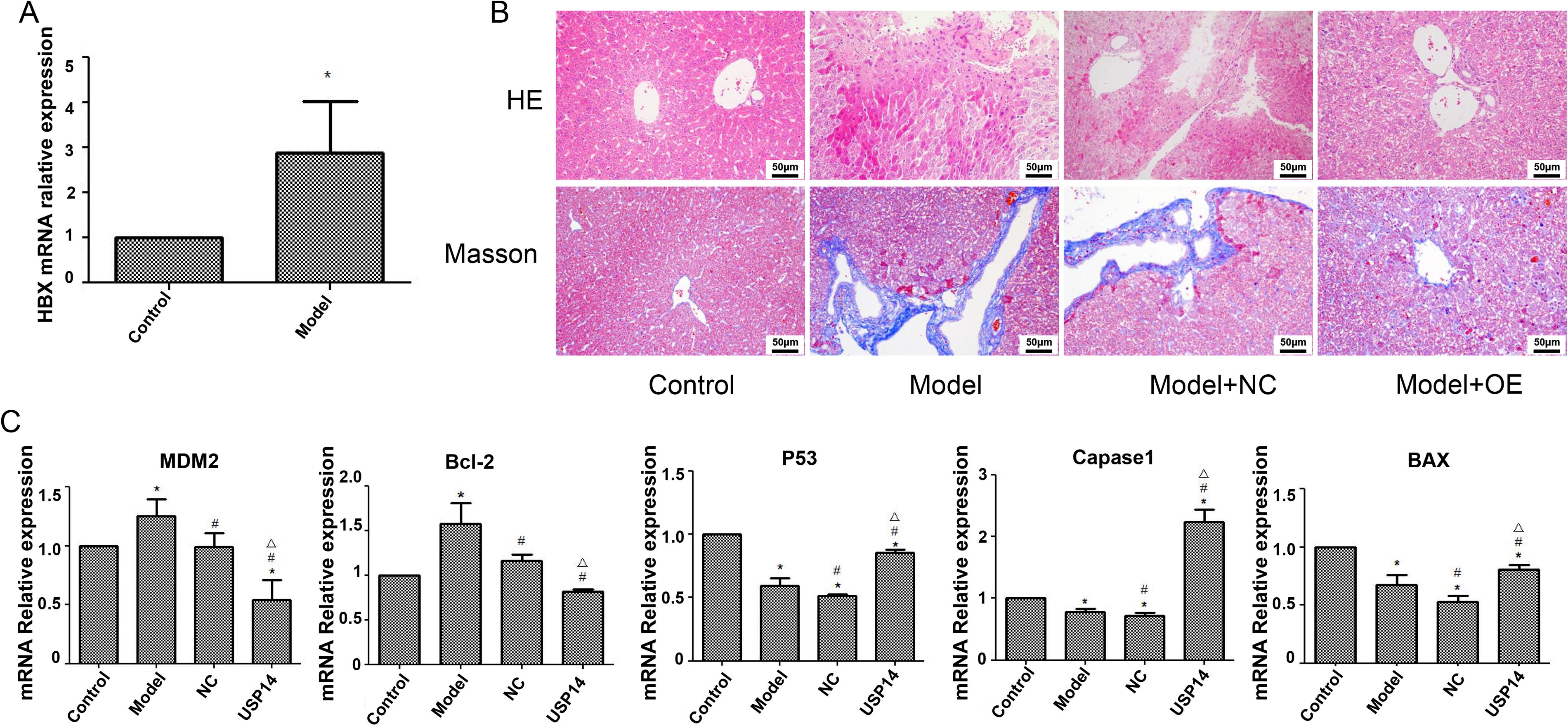
Overexpression of USP14 reduced HBx-induced hepatitis B in vivo.
Discussion
HBV infection is a public health issue worldwide, and reducing the occurrence of HBV-related liver diseases is of great significance. As a transcription factor, P53 binds to HBV enhancers and inhibits the activity of both HBV enhancers and promoters (Wu et al., 2022). MDM2, an E3 ubiquitin ligase, serves as a negative P53 regulator.
The MDM2 levels in HBV-related HCC patients increase (Wang et al., 2020). Overexpression of HBx is an important driving factor for HBV infection, triggering changes in cellular behavior through interactions with various host cytokines, such as stimulating Src kinase and cyclin gene expression (Bouchard et al., 2001) or increasing the activation rate and level of cyclin-dependent kinase and RAS (Benn and Schneider, 1995), inducing liver cells to enter the cell cycle faster, disrupting the balance between normal liver cell proliferation and apoptosis, promoting proliferation but inhibiting cell apoptosis, thus resulting in malignant transformation of normal liver cells (Fu et al., 2012; Li et al., 2022).
This study not only confirms the importance of ubiquitination in HBV progression but also identifies USP14 as a pivotal regulator of P53 stability. This novel insight suggests that targeting USP14 could represent an innovative strategy for delaying HBV invasion and development of HBV-related liver diseases, providing a unique therapeutic angle not previously explored in depth (Kong et al., 2019). Following transfection with the USP14 overexpression plasmid, the model group exhibited inhibited cell proliferation, decreased MDM2 and Bcl-2 expression in cells and liver tissue, and upregulated P53, Bax, and Caspase-1 expression. These findings suggested the importance of USP14 in delaying HBV invasion and hepatitis development.
P53 acts as a transcriptional repressor for USP14. Using chromatin immunoprecipitation (ChIP) assays, we found that P53 directly binds to the promoter region of USP14, leading to its downregulation17. This repression of USP14 may play a role in controlling HBV-induced tumor progression by limiting the deubiquitination activity of USP14 on P53 itself. Conversely, SP1 was found to enhance USP14 transcription. Dual-luciferase reporter assays revealed that SP1 binding to the USP14 promoter significantly increases its transcriptional activity, promoting USP14 expression (Vellingiri et al., 2020).
USP14 exerts its effect by stabilizing the tumor suppressor protein P53 through deubiquitination, preventing MDM2-mediated degradation (Ma et al., 2020). This stabilization promotes the transcription of proapoptotic genes such as Bax and Caspase-1 (Lv et al., 2020), enhancing apoptosis. The balance between cell proliferation and apoptosis is further modulated by the increased levels of P53, which acts as a central node in regulating the cell cycle. In addition to its role in apoptosis, USP14 inhibits HBV-induced cell proliferation by activating the P53 signaling pathway. Elevated P53 levels lead to the suppression of cell cycle regulators, thus reducing the proliferation of infected liver cells (Pant et al., 2019). This regulation is crucial in preventing the oncogenic transformation typically seen in HBV infections.
In addition to these direct regulators, our study uncovered that the NF-κB signaling pathway plays a significant role in modulating USP14 expression during HBV infection (Tarocchi et al., 2014). NF-κB activation, which is often induced during chronic inflammation in HBV-infected liver tissue, enhances USP14 transcription. This pathway was investigated using specific NF-κB inhibitors, which led to a marked reduction in USP14 mRNA and protein levels. These results suggest that USP14 regulation is part of the broader inflammatory response orchestrated by NF-κB in HBV-induced HCC.
These findings provide a more comprehensive understanding of how USP14 is regulated at both transcriptional and post-transcriptional levels. The interaction between transcription factors, noncoding RNAs, and signaling pathways in controlling USP14 expression highlights its critical role in the pathogenesis of HBV-related liver diseases.
Limitations of the Study
Our research is preliminary. There are quite a few problems with the study that are not clarified. First, we did not further explore the mechanisms affecting cell proliferation and apoptosis. Second, we did not reveal the specific transcription factors and noncoding RNAs that regulate USP14. Further studies are needed to explore these deficiencies.
Conclusion
To sum up, both the cell and animal model results showed that in hepatitis B models overexpressing USP14, MDM2 is downregulated, thus upregulating P53 to delay the invasion of HBV and the progression of hepatitis, confirming USP14’s inhibitory effects on HBV transcription and replication. We further found that USP14 weakens HBV activity by upregulating the expression of P53. This study reinforces the importance of the interaction between P53 and ubiquitin-proteasome pathway in virus invasion, suggesting that influencing this interaction may be a promising antiviral strategy. This study provided new ideas and targets for the treatment of HBV.
Footnotes
Acknowledgments
The authors would like to thank the Key Laboratory of Early Prevention and Treatment for Regional High-Frequency Tumor, Ministry of Education (GKE-ZZ202216); Key Laboratory of High-Incidence-Tumor Prevention and Treatment of the Ministry of Education (GKE-ZZ202129); Key Laboratory of High-Incidence-Tumor Prevention and Treatment of the Ministry of Education (GKE-ZZ202004); Guangxi Medical University Training Program for Distinguished Young Scholars; and Key Laboratory of Early Prevention and Treatment for Regional High-Frequency Tumor, Ministry of Education (GKE-ZZ202309) for their support in publishing this article.
Authors’ Contributions
All authors contributed to the study’s conception and design. Material preparation, data collection, and analysis were performed by R.-y.M., X.-b.W., J.C., J.-z.Y., M.W., B.Z., K.L., and Z.-m.G. The first draft of this article was written by T.B., Z.-h.T., F.-x.W., and L.-q.L., and all authors commented on previous versions of the article. All authors read and approved the final article.
Availability of Data and Materials
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Author Disclosure Statement
The authors have no conflicts of interest related to this publication.
Funding Information
This study is supported by the