
Abstract
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), the virus that causes COVID-19, has been spreading and changing globally. Adjuvant-based vaccines can improve vaccine protection by enhancing the immune response. Bacterial flagellin is a potent adjuvant and promotes protective immune responses. Here, we successfully expressed and purified the S1 subunit of SARS-CoV-2. The adjuvanticity of flagellin (FliC) of Salmonella Typhimurium in mice was determined by combining it with the recombinant S1 subunit vaccine. FliC-adjuvanted S1 vaccine could induce significantly enhanced S1-specific Immunoglobulin G (IgG), IgG1 and IgG2a titers, SARS-CoV-2-neutralizing antibodies, and levels of Th1 type (TNF-α and IFN-γ) and Th2 type (Interleukin-5 (IL-5), IL-4, IL-10, and IL-13) cytokines in splenocytes compared with the S1 alone group. Additionally, the titers of S1-specific IgG antibodies in the FliC adjuvant group could maintain a high level for at least 2 months. These results indicated that the FliC-adjuvanted S1 subunit vaccine could trigger strong humoral and cellular immune responses, which could promote the ongoing development of COVID-19 vaccines.
Introduction
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has caused the outbreak of coronavirus disease 2019 (COVID-19). It has been spreading and changing since 2019. Although many COVID-19 vaccines have been developed, additional effective vaccines are still needed to meet the global demand (Hadj Hassine, 2022). Of these, recombinant protein vaccines are safe and effective in preventing SARS-CoV-2 infection (Tabynov et al., 2022).
Spike (S), membrane (M), nucleocapsid (N), and envelope (E) are the four primary structural proteins of SARS-CoV-2 . S protein consists of the receptor-binding domain (RBD) in the S1 subunit, which encompasses multiple neutralizing epitopes crucial for the elicitation of neutralizing antibodies and the establishment of protective immunity. Furthermore, T-cell responses targeting the S protein have been extensively characterized in patients with COVID-19 (Arashkia et al., 2021; Pollet et al., 2021; Suryawanshi, 2023; Yadav et al., 2023). Hence, the recombinant S1 protein of SARS-CoV-2 was investigated as a promising vaccine candidate in this study.
Recombinant protein vaccines are safer options. Adjuvanted vaccines have the capacity to elicit robust immune responses, encompassing both humoral and cellular immunity, particularly in populations with compromised or suboptimal immune function, and the development of adjuvants is crucial for the research of subunit‐based vaccines against COVID‐19 (Castrodeza-Sanz et al., 2023). Flagellin, the structural protein constituting bacterial flagella, serves as a potent agonist for TLR5. Owing to their capacity to modulate innate immune responses, flagellins are gaining recognition as potential adjuvants in vaccine development. Numerous vaccines incorporating flagellin as an adjuvant are currently undergoing preclinical evaluation, targeting diseases such as influenza, malaria, AIDS, tetanus, and leptospirosis (Nedeljković et al., 2021). In our previous study on the H7N9 influenza virus subunit vaccine, it was demonstrated that FliC significantly enhances antigen-specific humoral and cellular immune responses (Song et al., 2018). However, additional research is necessary to clarify the function of FliC as an adjuvant in COVID-19 subunit vaccines.
In this study, we successfully expressed and purified the SARS-CoV-2 S1 subunit protein and Salmonella Typhimurium FliC. The S1 protein was combined with FliC to evaluate the adjuvant activity in promoting S1-specific immune responses. It was demonstrated that Salmonella FliC is a strong adjuvant that can enhance the efficacy of the S1 subunit vaccine and trigger robust and balanced Th1 and Th2 immune responses in mice. FliC adjuvant promotes high levels of antibodies neutralizing SARS-CoV-2 and lymphocyte responses.
Materials and Methods
Mice and ethical statement
Specific pathogen-free BALB/c female mice of 6–8 weeks were obtained from Beijing Vital River Laboratory Animal Technology (Beijing, China). The mice were housed in 12 h cycles of light and darkness at 25°C and 50% humidity. All mice were provided ad libitum with standard chow diets in the assays. All mouse experiments were approved by the Ethics Committee of the Animal Experiments of Yangzhou University [Approval ID: SYXK (Su) 2022-0044] and conducted according to the guidelines for animal care and ethics.
Expression, purification, and identification of the S1 subunit protein
We prepared Escherichia coli expressing the SARS‐CoV‐2 S1 subunit protein as follows. The DNA sequence encoding the S1 subunit (NC_045512.2, Val16‐Arg685) was synthesized by GenScript. The recombinant pCold-I-S1 and pGEX-6p-1-S1 plasmids were constructed using primers shown in Table 1 and the One Step Cloning Kit (Vazyme, Nanjing, China). We performed sequencing to verify the recombinant plasmids. Subsequently, the transformation of these plasmids was conducted in E. coli (BL21, Takara, Dalian, China). The recombinant S1 subunit protein with a His-tag was expressed with the pCold-I-based Cold Shock Expression System according to instructions specified by the manufacturer (Sugiki et al., 2017). The recombinant protein GST-S1 was induced with Isopropyl β-D-1-thiogalactopyranoside (IPTG) at 30°C for 5 h. The purification of the soluble His-S1 and GST-S1 subunits was conducted with a His Bind Purification Kit (Novagen, Billerica, MA, USA) and GST Fusion Protein Purification Kit (GenScript), respectively. Finally, the purified S1 subunit was separated on sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and Western blotting (WB) was conducted to verify the immunoreactivity using the anti-RBD antibody (Sino Biological, Beijing, China). Endotoxin was removed from the protein and residual endotoxin was measured as described previously (Song et al., 2023a).
Primer Sequences Used for the Construction of the Recombinant Plasmids
Animal vaccination and sample collection
We randomly categorized BALB/c female mice at 6–8 weeks into three experimental groups (9 mice per group). Mice immunization was conducted via intraperitoneal injection of phosphate buffered saline (PBS) control, 10 μg S1, 10 μg S1 + 10 μg FliC [S1 + FliC, Salmonella Typhimurium Flagellin (FliC) was extracted as described previously (Song et al., 2023a)] in 100 μL on day 0, 14, and 28 (at 2-week intervals). The sera from animals on days 26, 40, 56, 70, 84, and 98 were collected to detect S1-specific total IgG antibodies. After 2 weeks of the last immunization, the spleens of immunized mice were harvested to evaluate the cellular immune responses.
Enzyme-linked immunosorbent assay
The titers of total IgG, IgG1, and IgG2a isotypes of S1-specific antibodies in the sera of immunized mice were measured as described previously (Song et al., 2018). Costar polystyrene high binding 96-well plates (Corning, Tewksbury, MA, USA) were coated using 0.05 μg/well GST-S1 subunit protein in 100 μL sodium carbonate coating buffer at 4°C overnight. The HRP-conjugated goat antimouse IgG (1:10,000), IgG1 (1:5,000), and IgG2a (1:5,000) (Invitrogen, CA, USA) were used as the secondary antibodies.
Isolation of splenocytes from immunized mice
The spleen from mice was harvested after 2 weeks of the last vaccination and collected in RPMI1640 medium containing 1% penicillin–streptomycin and 10% FBS. A single-cell suspension of the spleen was made as previously described (Song et al., 2018). Briefly, the lymphocytes from the spleens were isolated using Lymphoprep (specific gravity 1.077, Sigma, MO, USA) and density gradient centrifugation following the guidelines specified by the manufacturer. We seeded 1 × 106 splenocytes from immunized mice/well in 24-well plates and treated the cells with 5 μg/mL recombinant S1 subunit protein (Sino Biological) at 37°C for 5 h. The total RNA of the splenocytes was extracted, and quantitative real-time PCR (qRT-PCR) was conducted to determine the expression levels of cytokines. The secretion levels of cytokines in culture supernatants were quantitated using IFN-γ and IL-4 enzyme-linked immunosorbent assay (ELISA) kits (Beyotime, Shanghai, China) performed according to the manufacturer’s instructions.
RNA extraction and qRT-PCR
The total RNA of the splenocytes of immunized mice was harvested with the total RNeasy Plus Mini Kit (Qiagen, Hilden, Germany). Next, cDNA was synthesized from RNA with a PrimeScript RT reagent Kit (Takara) following the manufacturer’s instructions. IL-4, IL-5, IL-10, IL-13, TNF-α, and IFN-γ expression was determined with SYBR Green master mix (Roche Diagnostics, Tokyo, Japan) by qRT-PCR using the ABI 7500 Fast Real-Time PCR System as described previously (Song et al., 2023a). The sequences of qRT-PCR primers were synthesized by GenScript (Table 2).
Primer Sequences Used for the Quantitative Real-Time PCR
Pseudoviral neutralization assay
We determined the titers of neutralizing antibodies in the sera of the immunized mice using SARS-CoV-2 Pseudovirus Neutralization Kit_Luc Reporter (GenScript) as described previously (Song et al., 2023b).
Statistical analysis
All experiment data are represented as the standard error of the mean. We statistically analyzed the data with the GraphPad Prism (version 8.0.2) software (San Diego, CA, USA). One-way Analysis of Variance (ANOVA) and Student’s t test were performed for analyzing the data. p < 0.05 (*), p < 0.01 (**), and p < 0.001 (***) were considered to be statistically significant.
Results
Expression and characterization of the recombinant S1 proteins
The S1 sequence of SARS-CoV-2 was cloned into pCold-I and pGEX-6p-1 vectors with a His-tag and GST-tag, respectively. The soluble recombinant His-S1 and GST-S1 proteins were successfully expressed and purified using His- and GST-tags affinity chromatography. This procedure was conducted utilizing a native purification protocol, thereby obviating the need for any refolding steps. The results showed the expected molecular weights of the proteins, that is, 75 kDa for His-S1 and 101 kDa for GST-S1 proteins (Fig. 1A, B). WB was conducted using anti-RBD antibodies to validate the immunoreactivity of the purified His-S1 and GST-S1 proteins (Fig. 1C, D).

Expression, purification, and identification of S1 protein of SARS‐CoV‐2. A DNA sequence encoding the S1 protein was inserted in the pCold-I and pGEX-6p-1 plasmids, respectively. Escherichia coli (BL21) was used for plasmid transformation and IPTG was used for protein induction.
Humoral immune responses triggered by FliC-adjuvanted S1 subunit vaccine
The adjuvant effect of FliC on the recombinant S1 protein was determined. BALB/c mice were immunized intraperitoneally with three doses of 10 μg recombinant S1 subunit protein adjuvanted with FliC or unadjuvanted at an interval of 2 weeks. The indirect ELISA was performed to evaluate the titers of IgG and IgG subtypes, such as IgG1 and IgG2a, specific to S1 in the serum of mice collected after 2 weeks of the second or third vaccination (Fig. 2A). On day 40, a significantly strong S1-specific IgG response was induced by S1 + FliC compared to S1 alone or PBS control. In addition, 10-fold higher titers of S1-specific IgG were induced by S1 + FliC following the third vaccination compared to the second vaccination (Fig. 2B). A significantly high titer of IgG1 and IgG2a specific to S1 was observed on the S1 vaccination compared to the PBS control group, and the S1-specific IgG subtypes induced by S1 + FliC were significantly increased compared to those induced by S1 or PBS control (Fig. 2C). Furthermore, the IgG1/IgG2a ratios were 0.6 and 1.4 in the group immunized with FliC-adjuvanted vaccine and S1 alone group, respectively, which indicating that the S1 + FliC vaccine induced a balanced IgG1 and IgG2a response.
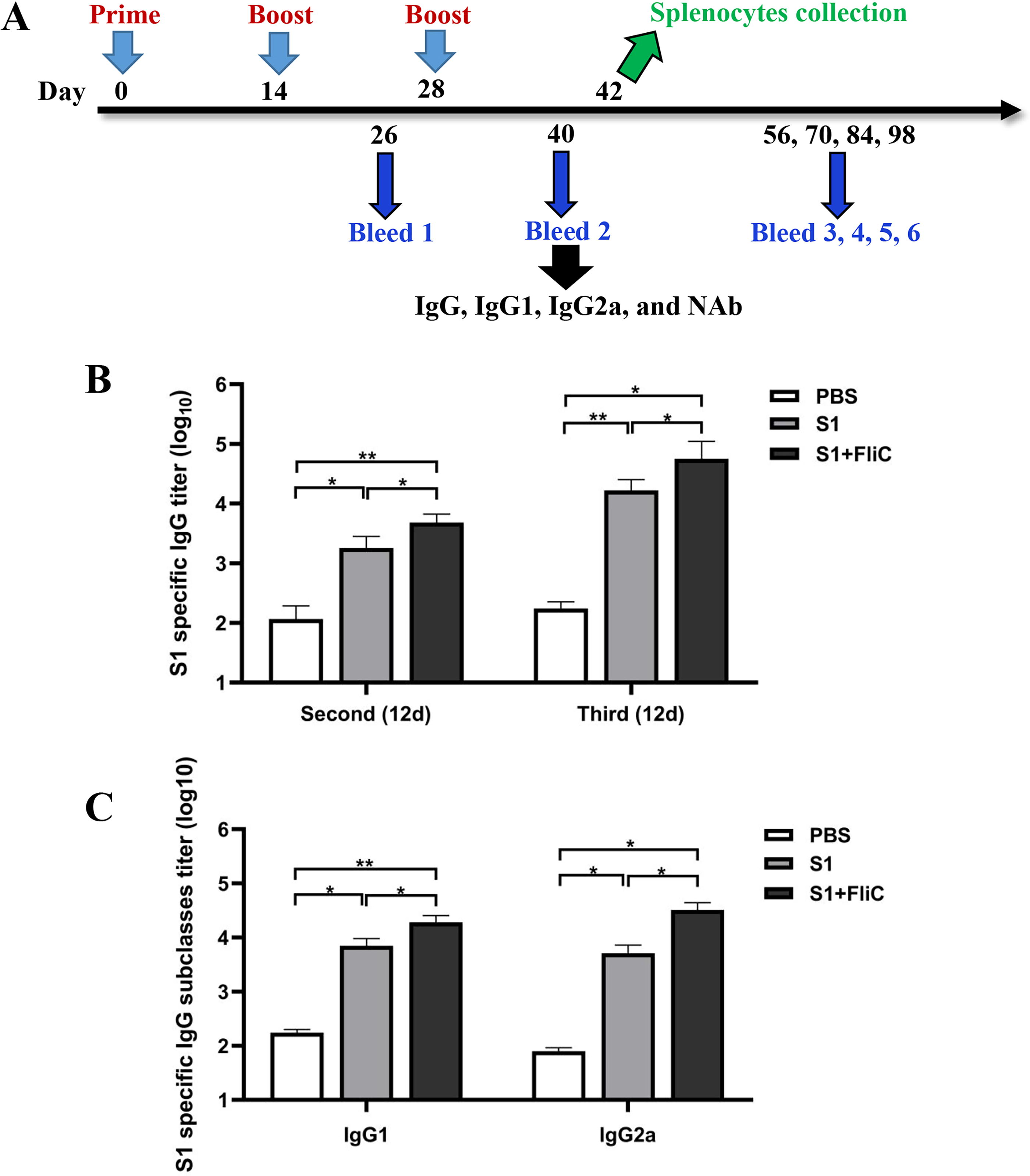
S1-specific antibody responses of FliC-adjuvanted S1 vaccine in mice.
Cellular immune responses to FliC-adjuvanted S1 subunit vaccine
To determine whether the FliC adjuvant could induce S1-specific cellular immune responses, we measured the levels of cytokines produced by the spleen in response to the S1 antigen. The spleens were harvested from mice sacrificed after 2 weeks of the third vaccination. The splenocytes were treated with S1 subunit protein for 5 h, and cytokine expression was determined. qRT-PCR results suggested a significant increase in the expression levels of Th1 type cytokines (IFN-γ and TNF-α) and Th2 type cytokines (IL-4, IL-5, IL-10, and IL-13) in the group of S1 + FliC compared to S1 alone group (Fig. 3A). In addition, the results of cytokines release showed that significantly higher levels of IFN-γ and IL-4 were observed in culture supernatants from FliC-adjuvanted group compared with the S1 or PBS group (Fig. 3B). Overall, the upregulation of splenic cytokine production indicated that S1 + FliC promoted Th1/Th2-balanced cellular immune responses.
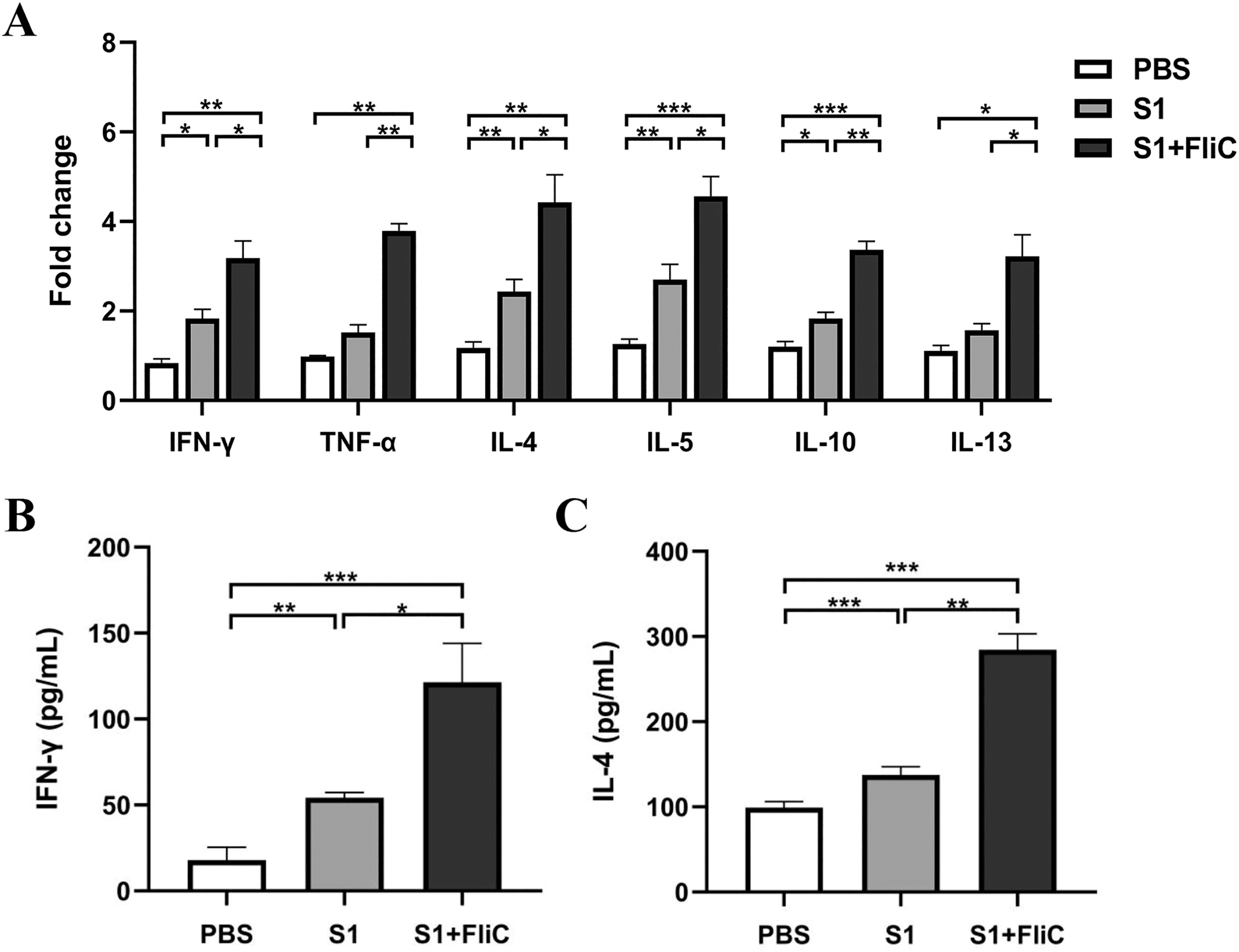
FliC-adjuvanted S1 subunit vaccine induces the production of Th1/Th2 type-specific cytokines in splenocytes. The spleen was harvested after 2 weeks of the last vaccination. The splenocytes were isolated and treated using 5 μg/mL of S1 protein for 5 h.
FliC-adjuvanted S1 subunit vaccine triggers strong neutralizing antibody responses
To further investigate the efficacy of the antibodies generated by the vaccine, we collected the sera from immunized mice and the neutralization assay of SARS-CoV-2 pseudovirus was performed. The results revealed significantly high titers of neutralizing antibodies in the sera collected from the S1 and S1 + FliC groups after 2 weeks of the third vaccination, compared to the control group. The mean antibody titers were 41 and 115, respectively. A significant increase in the neutralizing antibodies level was also observed in the mouse sera of the S1 + FliC group compared to the S1 alone group (Fig. 4A). These data showed that sera of mice immunized with the FliC-adjuvanted S1 vaccine could effectively neutralize SARS-CoV-2 pseudovirus.

Titers of neutralizing antibodies and the long-term antibody responses in mice immunized with the S1 subunit vaccines.
The longevity of S1-specific IgG titers in the sera
We observed S1-specific IgG antibody titers for 3 months after the first vaccination. The sera of mice were collected and analyzed every 2 weeks after vaccination. The long-term immunogenicity of the S1 subunit vaccines in mice was measured until day 98. The results showed S1-specific antibody responses in mice in the S1 alone and S1 + FliC groups. The S1-specific antibody titers could be detected following the second vaccination and gradually increased following the third dose. The IgG titers in the sera of mice in the S1 group were highest on day 40. In the S1 + FliC group, the geometric mean titer (log10) of the SARS-CoV-2 antibody was 4.8 on day 40, which decreased to 4.4 on day 70, and 3.9 on day 98 (Fig. 4B). These results indicate that S1 + FliC vaccination conferred humoral responses against SARS-CoV-2 for at least 2 months.
Discussion
Adjuvant-based COVID-19 subunit vaccines can improve vaccine protection. In this study, we determined the potential of FliC adjuvant to the S1 subunit protein of SARS-CoV-2. The results showed enhanced immunogenicity and highly robust immune response in mice in the S1+ FliC group compared to the S1 alone group. FliC-adjuvanted S1 subunit vaccine elicited S1-specific antibody responses for at least 2 months, indicating the FliC-adjuvanted S1 protein antigen has been shown to elicit robust humoral immune responses in mice. Comparable outcomes have been observed with subunit vaccines incorporating flagellin adjuvants in the context of animal infectious viruses. For instance, immunization with the African swine fever p30 protein fused with Salmonella Typhimurium FliCΔD2D3 significantly enhanced humoral immune responses in mice (Huang et al., 2025). Similarly, two immune-subdominant (ISD) antigens of Babesia bovis, when combined with a FliC adjuvant, induced strong humoral immune responses in cattle (Rojas et al., 2024).
The RBD of the S1 protein forms crucial interactions with ACE2 receptors in humans for viral entry. We measured pseudovirus-neutralizing antibody titers to determine the efficacy of the induced antibodies. The results revealed that the FliC-adjuvanted S1 vaccine could elicit significantly higher neutralizing antibodies against pseudovirus compared to the control group. Comparable high levels of serum-neutralizing antibody titers were observed following immunization with the FliC-adjuvanted H7N9 influenza subunit vaccine (HA1-2-FliC) (Song et al., 2019). Additionally, prior research has confirmed the efficacy of flagellin as an adjuvant in mCOE vaccines against the porcine epidemic diarrhea virus, eliciting an immune response with neutralizing antibodies similar to those produced by an inactivated vaccine (Murtaza et al., 2024). Collectively, these studies substantiate the adjuvant potential of flagellin.
An ideal vaccine should trigger a potent Th1-type response for conferring protection against SARS-CoV-2 (Vabret et al., 2020). An increase in IgG1 and IgG2a levels was observed in mice in the S1 + FliC group, and the IgG1/IgG2a ratio was 0.6, thus indicating a balanced Th1/Th2 profile. Ratios between 0.5 and 2.0 indicate a mixed or balanced response (Yang et al., 2008). The similar result was also observed in the study of flagellin (FliC) of enteroaggregative E. coli fused to FimH antigen (Hong et al., 2012). However, it is in contrast to another study reporting that codelivery of flagellin with inactivated influenza vaccine induced a Th2 response in mice (Honda-Okubo et al., 2015). We speculate that the difference in mouse strain and vaccination strategies affects the immune response bias. Moreover, the Th1/Th2 cytokine profiles (IFN-γ and IL-4) indicate that the FliC adjuvant avoided strong Th2-biased immune responses, observed previously in vaccine adjuvanted with alum for respiratory disease in SARS-CoV-2 (Wang et al., 2021). Compared to the SpikoGen® vaccine, which utilizes the Advax-CpG55.2 adjuvant for clinical applications, the FliC-adjuvanted S1 vaccine demonstrated a higher induction of IFN-γ and IL-4 expression levels in mice (Li et al., 2022; Petrovsky, 2024).
The antigen-flagellin fusion strategy maintains the independent structures and functions of both components, improving protein folding and antigen presentation (Honko et al., 2006). It offers a stronger adjuvant effect than just mixing flagellin with antigen (Mizel and Bates, 2010). Our earlier studies showed that the HA1-2 and FliC fusion boosts antibody responses and targets influenza virus antigens effectively (Song et al., 2018). To achieve specific effects, adjuvants are sometimes combined, such as with aluminum salts, to boost vaccine responses. Various formulations, such as AS04 and AS01B, incorporate TLR agonists. In a human papillomavirus vaccine study, TLR4/5 agonists induced Th2 responses, but adding a TLR7 agonist balanced Th1/Th2 responses (Mashhadi Abolghasem Shirazi et al., 2023). Future research could investigate using flagellin in more effective vaccine strategies.
Conclusion
The Salmonella FliC exhibits potent adjuvant activity and is suitable for developing the S1-based SARS-CoV-2 subunit vaccine. The FliC-adjuvanted S1 subunit vaccine could trigger the production of a high level of virus-neutralizing antibodies and Th1/Th2-balanced immune responses. Furthermore, the FliC adjuvant could be formulated with other potential subunit vaccines and other forms of vaccines in future studies.
Footnotes
Authors’ Contributions
Conceptualization, planning, and experimental design: L.S., X.J., and Z.P. Performed the experiments and data acquisition: L.S., Y.C., Q.W., R.T., and Y.W. Data analysis: L.S., Y.C., Q.W., and C.M. Drafting and revision of the article: L.S., X.J., and Z.P.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.
Funding Information
This work was supported by the National Key Research and Development Program of China (2022YFC2604200), the National Natural Science Foundation of China (32102679), the Key Research and Development Program (Modern Agriculture) Project of Jiangsu Province (BE2021331), the 111 Project (D18007), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).