
Abstract
Abstract
Apart from playing a crucial role in tumor suppression, the p53 signaling pathway also has important functions in the regulation of metabolism, organ development, and cellular senescence. A recent important advance in the p53 field is the uncovering of presence of various p53 isoforms in vivo. Although full-length p53 is well known to promote cell apoptosis and cell cycle arrest, the roles of p53 isoforms in these processes are still elusive. In this review, we summarized the recent studies on the p53 isoform Δ113p53 regarding its regulation and function in zebrafish and discussed the potential use of Tg(Δ113p53:gfp) transgenic fish for identification of novel factors involved in the p53 pathway and for screening of new compounds for cancer therapy.
Introduction
Although the transcriptional activity of p53 plays a key role in p53 function, a seemingly unrelated activity for p53 in the cytoplasm has also been described. A number of reports have demonstrated that p53 can directly localize to the mitochondria and induce apoptosis without its transcriptional activity.10,11 In mitochondria, p53 can interact directly with antiapoptotic proteins, such as Bcl-2 and Bcl-XL, acting in a way similar to the so-called BH3-only protein (PUMA).
Activation of p53 plays a central role to repress tumorigenesis, but too much p53 would cause various developmental abnormalities and lethality. In normal conditions, p53 protein interacts with Mdm2, an E3 ligase, which ubiquitylates p53 and targets it for proteasomal degradation. The p53–Mdm2 interaction motif is within the p53 transactivation domain. mdm2 itself is a p53 target gene. The negative feedback loop between p53–Mdm2 plays a key role in maintaining the low level of p53 in a normal cell.12,13
Although p53 has been extensively studied, a number of crucial questions remain unclear; for example: How does p53 integrate so many different stress signals that lead to an ultimate choice between life and death? What roles do the p53 network genes play in tumorigenesis and cancer therapies? In year 2005, up to 10 p53 isoforms were identified. It has been demonstrated that these p53 isoforms can modulate p53 functions either synergistically or antagonistically, depending on the isoform structures and the target genes affected.14–16 It has also been shown that nine isoforms are abnormally expressed in tumor tissues compared with the normal cells. 15 Understanding the regulation and functions of p53 isoforms will have a profound impact on these crucial questions. Because a number of reviews have already provided an extensive summary on p53 isoforms in human,17–20 this review mainly focuses on the studies of p53 isoform Δ113p53 in zebrafish.
p53 Isoforms
There are three members in the p53 family: p53, p63, and p73. Compared to p53, p63 and p73 each has two additional alternative domains at their C-terminus: sterile alpha motif and transcription inhibition domain. Although p63 and p73 are able to transactivate p53-responsive genes, p53, p63, and p73 proteins are not entirely functionally redundant. p53 plays a crucial role in stress response and tumor suppression, whereas p63 and p73 have specific biological functions in development and cell differentiation. 17 Members of the p53 family express multiple mRNA variants due to alternative splicing and alternative promoter utilization. The human and mouse p63 gene encodes at least six isoforms (two N-terminus variants generated by one alternative promoter located at intron 3, three C-terminus variants generated by alternative splicing); p73 encodes at least 29 isoforms that are translated from 35 mRNA variants (five N-terminal variants: one from an alternative promoter located at intron 3, four from alternative splicing; seven C-terminal variants from multiple alternative splicings).17,19
For many years, the structure of the p53 gene was thought to be much simpler than p63 and p73. This concept was torn down when up to 10 p53 isoforms were identified in human cells (Fig. 1).14,21–23 There are three N-terminus (full-length p53, Δ40p53, and Δ133p53) and three C-terminus (full-length p53, p53β, and p53γ) isoforms. The Δ40p53 is generated by either alternative initiation of translation or alternative splicing of intron 2.14,24 The Δ133p53 is generated by an alternative promoter in intron 4. The alternative splicing in intron 9 generates the β and γ isoforms. The oligomerization domains of β and γ are, respectively, replaced by 10 and 15 additional residues. 15 The combination of three N-termini and three C-termini produces nine p53 isoforms (full-length p53, p53β, p53γ, Δ40p53, Δ40p53β, Δ40p53γ, Δ133p53, Δ133p53β, and Δ133p53γ) (Fig. 1A). The 10th isoform Δp53 is generated by noncanonical splicing between exon 7 and exon 9, resulting in the deletion of 198 nucleotides at the mRNA level, which corresponds to part of the DNA-binding domain and complete nuclear localization signal at the protein level (Fig. 1A). 23
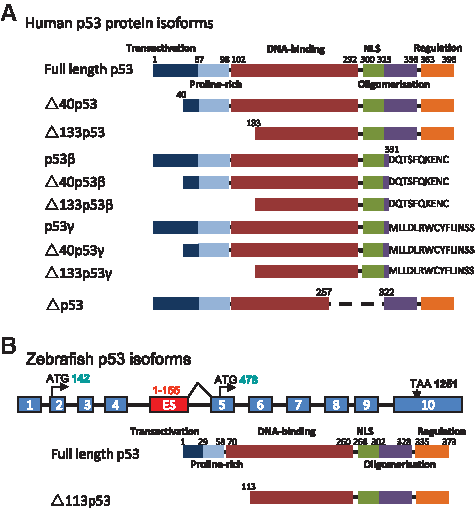
p53 isoforms. (
Very interestingly, the Δ133p53 transcribed from intron 4 was found to be conserved in zebrafish, Drosophila, and mouse.15,25 This evolutionary conservation suggests that p53 isoforms may play a very important role in the p53 functions. Up to date, Δ113p53 (ortholog of human Δ133p53) was the only p53 isoform reported in zebrafish. In fact, the finding of Δ113p53 in zebrafish was accidental. During the course of investigation of the zebrafish def−/− (digestive-organ expansion factor) mutant, Chen and colleagues carried out a microarray analysis to compare the gene expression profiles between the wild-type and the def−/− embryos. p53 was found to be upregulated by more than twofolds in the def−/− embryos. While trying to confirm this profile via RNA gel blot hybridization, the authors found a short form of p53 transcripts being expressed at very high level in the def−/− mutant, whereas it was almost undetectable in the wild-type embryos. To investigate the nature of this intriguing short-form p53 transcript, 5′-RACE and 3′-RACE experiments were performed. Two 5′-RACE products were observed, whereas only one product was found in 3′-RACE. Sequence analysis revealed that the longer 5′-RACE product corresponds to the original p53. As for the short form, it contains exons 5 to 12 together with 155 nucleotides derived from intron 4 immediately adjacent to exon 5 (Fig. 1B), suggesting it is initiated from an internal promoter in intron 4. 25 The short-form transcript encodes for a N-truncated p53 protein initiated at codon 113 (named as Δ113p53 thereafter), and this product is an apparent counterpart of the human Δ133p53 isoform. 15 Until now, several zebrafish p53 antibodies have been used to detect zebrafish p53 protein.26,27 Among these antibodies, one (from Lane's lab) was mapped to recognize the C-terminus of the p53 and was able to detect the induction of p53 by immunohistochemical assay but not by western analysis. 26 The other two antibodies from Lees' lab have not had their epitopes mapped yet. 27 Unfortunately, none of the antibodies was found to be able to detect Δ113p53 protein in the conditions which the Δ113p53 mRNA was induced. Apparently, there is an urgent to develop p53 antibodies that can recognize Δ113p53 protein.
Expression Regulation of Δ113p53
The finding of Δ113p53 and its aberrantly elevated expression in the def−/− mutant raise two questions: how is Δ113p53 regulated? What are the functions of Δ113p53? The regulation of Δ113p53 expression is implied in a work reported by Langheinrich et al. 13 Langheinrich and colleagues performed a study on the p53 pathway using zebrafish as a model system. The authors found that a short p53 transcript was strongly induced in zebrafish embryos when these embryos were either injected with mdm2 morpholino or treated with DNA-damaging drugs. The induction of the short p53 transcript was p53 dependent. The authors suggested that the short p53 transcript would be a result of the usage of the internal polyA signal site that exists in zebrafish p53.
Using a Δ113p53-specific probe to perform RNA blot hybridization, Chen and colleagues demonstrated that the short p53 transcript induced by DNA-damaging signals is Δ113p53. To test whether Δ113p53 induction during DNA-damaging stress is p53 dependent, they treated p53−/− embryos with γ-ray and DNA-damaging drugs and found that these treatments failed to induce Δ113p53 expression in the p53−/− mutant background. This result immediately prompted the authors to speculate if the upregulation of Δ113p53 in the def−/− mutant was also p53 dependent. The authors tested out this speculation by generating def−/− p53−/− double mutant, and the data showed that the elevated Δ113p53 expression is abolished in the double mutant which firmly establishes the relationship between Δ113p53 and p53: Δ113p53 expression is p53 dependent. 16
Promoter analysis demonstrated that a 4.113-kb genomic DNA fragment containing intron 4 of the p53 gene not only displays promoter activity but also can truly recapitulate the response of endogenous Δ113p53 both to the developmental and DNA damage signals. 16 There are three p53 REs within intron 4 of the p53 gene. Promoter deletion coupled with green fluorescence protein (GFP) reporter transient assays showed that the regions containing p53 REs are crucial for the promoter activities. ChIP assay demonstrated that the full-length p53 protein can directly bind to the promoter region containing two out of the three p53 REs. Therefore, p53 directly regulates Δ113p53 expression by binding to its promoter. 16
Functions of Δ113p53
Because Δ133p53 protein is an N-terminal-truncated p53 with the deletion of both the Mdm2-interacting motif and transcription activation domain together with partial deletion of the DNA-binding domain, it was postulated that Δ133p53 might act as a dominant-negative regulator of p53. In fact, cotransfection of full-length p53 with Δ133p53 impaired p53-induced cell apoptosis in the human cell line system. 15 However, recent findings from zebrafish model system indicated that Δ113p53 does not act in a dominant-negative manner toward p53 as expected, but instead it differentially modulates p53 response gene expression to antagonize p53 apoptotic activity. 16 Although injection of Δ113p53 mRNA alone showed little transcription activities in p53−/− embryos, coinjection of Δ113p53 and p53 altered the expression patterns of p53 response genes differentially in a gene-dependent manner. For example, compared to p53 injection alone, it was observed that coinjection of Δ113p53 and p53 greatly enhanced the expression of p21 and mdm2 whereas it did not do so for bax. In addition, p53 injection alone downregulated bcl2L, an antiapoptotic protein similar to human bcl-xL, but in the presence of Δ113p53, bcl2L levels actually increased. Manipulating Bcl2L expression itself confirmed that this was an important contributor to p53-dependent apoptosis. These experiments suggested that Δ113p53 antagonizes p53 apoptotic activity by specifically upregulating antiapoptotic gene expression. 16
The antiapoptotic activity of Δ113p53 raises the speculation that knockdown of Δ113p53 might sensitize the zebrafish embryos to γ-ray irradiation treatment. Indeed, Δ113p53 is strongly induced by γ-ray irradiation. Knockdown of endogenous Δ113p53 by a Δ113p53-specific morpholino in γ-ray-treated embryos significantly enhanced p53 apoptotic activity and resulted in 100% mortality, whereas γ-ray-treated normal control embryos retained more than 50% viability despite abnormal development. 16 This observation suggests that one of the biological functions of Δ113p53 is to protect zebrafish embryos against environmental or physiological stress conditions by suppressing p53 apoptotic activity. From the point of view of evolution, the Δ113p53 pathway is evolved to fulfill the first priority for a living organism: that is to survive under extreme stress conditions at the expense of abnormal growth and cancer development at the later stage of its life.
Although Δ113p53 functions to antagonize p53 apoptotic activity, apoptosis was still obviously enhanced even when Δ113p53 expression was induced in most of the DNA damage stress conditions, such as under DNA damage drug treatments, 13 morpholino injection, 28 γ-ray irradiation. 16 One possible explanation for this is that the efficiency of Δ113p53 antiapoptotic activity is dependent on the ratio of Δ113p53/p53 proteins. This rationale is inferred from the results showing that Δ113p53 mRNA rescue of the high mortality caused by p53 mRNA is in a Δ113p53 mRNA dosage-dependent manner. 16 Another explanation is that Δ113p53 might not be induced equally in all embryonic cells but is induced in a cell type or tissue-specific manner under stress conditions. This argument is based on the fact that Δ113p53 mRNA injection greatly reduced apoptotic cells throughout the wild-type embryos treated with γ-ray irradiation, likely due to the ubiquitous distribution of injected Δ113p53 mRNA in all cells. 16 Certainly, we cannot exclude other possibilities, for example strong induction of p53 by γ-ray irradiation causes apoptosis even before p53 can have a chance to induce the expression of Δ113p53 to protect cells.
Based on their extensive studies on the relationship between p53 and Δ113p53, Chen and colleagues propose that Δ113p53-p53 forms a new negative feedback regulation loop in the p53 pathway. 16 p53 is usually kept at a low level in normal cells.12,13 Mdm2 plays a key role in maintaining the low level of p53 in a cell. 29 In addition to the Mdm2-p53 negative feedback regulation loop, p53 is also regulated at multiple levels via protein stabilization and activation. 30 In the new p53-Δ113p53 negative feedback regulation loop, p53 activates Δ113p53 expression in response to the developmental or DNA-damaging signals that in turn inhibits p53-mediated apoptosis via activation of the Bcl2-related antiapoptotic gene (Fig. 2). In the future it would be essential to find out if the p53-Δ113p53 pathway operates only specific for zebrafish embryo development or is also active in adult fish, or represents a general mechanism in all vertebrates, especially in human.
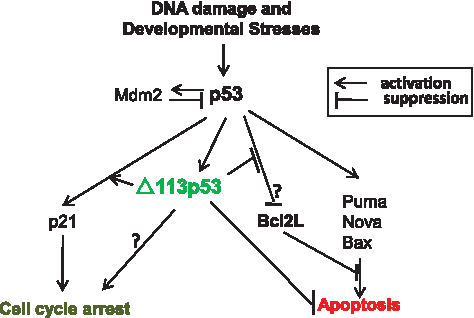
Δ113p53/p53 negative feedback loop. p53 and Mdm2 form a negative feedback regulation loop, in that p53 activation induces the Mdm2 expression and the latter then triggers the degradation of p53 to remove p53. p53 and Δ113p53 also form a negative feedback loop, in that activation of p53 induces the expression of Δ113p53 that in turn acts specifically to antagonize the apoptotic activity of p53 without inactivating cell cycle inhibitor gene expression. We propose that Δ113p53 might switch the p53 pathway to favor cell cycle arrest in response to the developmental or stress signals.
Tg(Δ113p53:gfp) Transgenic Fish for Genetic and Chemical Screening
At the core of the DNA damage signaling apparatus is a pair of related protein kinases, ataxia telangiectasia mutated (ATM), and ATM and RAD3-related (ATR), which are activated by DNA damage. ATM with its regulators the MRN (Mre11-RAD50-NBS1) complex senses double-strand breaks. 31 ATR with its regulator, ATR-interacting protein, senses single-strand DNA breaks. Both ATM and ATR then phosphorylate Chk2 and Chk1 (checkpoint kinases), respectively, to initiate a secondary wave of phosphorylation events to extend signaling. In human, ATM and ATR phosphorylate p53 on Serine-15 and Chk1 and Chk2 phosphorylate p53 on Serine-20. 32 Phosphorylation of p53 at N-terminal sites stabilizes the p53 protein by disrupting Mdm2 binding and promoting acetylation and phosphorylation of the C-terminus. 32 However, mutations in many of p53 protein serine and threonine residues that block p53 phosphorylation events still result in fairly normal p53 activation and function. 2 These studies indicate that there are other mediators in the activation of the p53 protein in DNA damage response in addition to ATM, Chk2, and ATR, Chk1. One of the best ways to identify upstream mediators of the p53 pathway is to screen for genetic mutants. For well-known reasons, human cell line and mouse systems are not suitable for genetic screening. Caenorhabditis elegans and Drosophila are excellent genetic systems, but the functions of p53 is only limited to the germline cells in adult worms and flies.33,34 Seemingly zebrafish is an excellent system for this purpose. It is necessary to point out that the N-terminus of p53 is very poorly conserved between humans and zebrafish. There is one Serine-6 Glutamine-7 site in the N-terminus of zebrafish p53 which might be analogous to Serine-15 in humans. Whether this Serine-6 Glutamine-7 site is the target of ATM/ATR is still unverified, and there is no clear Chk1/Chk2 target serine in zebrafish p53 protein. The lack of conservation of the Mdm2 interaction domain also makes us wonder how DNA damage signals disrupt the interaction between p53 and Mdm2 to activate p53 in zebrafish although genetic evidence shows that Mdm2 regulates p53 in fish in a manner analogous to the mammals. 13
In the study of Δ113p53 regulation, Chen and colleagues cloned a 4.113-kb genomic DNA fragment that starts immediately upstream of the start codon ATG of Δ113p53 and ends downstream of the transcriptional start site of full-length p53. This 4.113 kb DNA fragment was used to generate the Tg(Δ113p53:gfp) transgenic fish. 16 The GFP expression in the transgenic fish recapitulates the endogenous Δ113p53 expression, which normally keeps at a very low level but is strongly upregulated by DNA damage signals. Because the GFP expression is totally p53 dependent, the Tg(Δ113p53:gfp) transgenic fish provides an excellent reporter system to screen for zebrafish mutants affected in the upstream events of p53 activation in the DNA damage response via genetic approach.
The human p53 codon Methionine-246 (M246) is mutated in 124 different human tumors, 8 of which exhibit the same amino acid change from a methionine to a lysine (K). In addition, this codon is positioned between other known mutation hot spots in the DNA-binding domain of the human TP53 gene (e.g., codons 245, 248, and 249). 35 The zebrafish tp53M214K mutant is an ortholog to the human tp53M246K. The tp53M214K mutant embryos failed to undergo apoptosis and did not arrest cell cycle in response to γ-radiation. 36 Beginning at 8.5 months of age, 28% of tp53M214K mutant fish developed malignant peripheral nerve sheath tumors. Immunohistochemistry with a monoclonal antibody against zebrafish p53 revealed that the tp53M214K protein in the mutant embryos was present and accumulated in response to the DNA-damaging agents in a manner similar to that of the wild-type protein in wild-type embryos; however, p53 target genes were only activated in response to DNA damage in the wild-type embryos, not in the tp53M214K mutants. 26 These results suggested that the mutated protein is not unstable, but instead has lost its transcriptional activation activity in response to DNA damage stress. Similarly, like the endogenous Δ113p53 expression, the gfp expression in Δ113p53:gfp is also not induced by DNA-damaging signal in the tp53M214K mutants. Essentially, the Tg(Δ113p53:gfp) transgenic fish provides an excellent system to screen for small molecules that can restore p53 transcriptional abilities in response to the DNA damage in the tp53M214K mutant background. Such small molecules are good candidates for cancer therapeutics.
Tg(Δ113p53:gfp) Transgenic Fish for Evaluating DNA-Damaging Toxicities and Screening DNA-Damaging Reagents
There is a high demand for reliable and ethically acceptable methods to evaluate the developmental toxicity of pharmaceuticals, industrial chemicals, and the environmental conditions. Offering experimental advantages such as large offspring population, small size embryo, cheap maintenance, availability of genome sequence, and transgenic reporter systems, the zebrafish model undoubtedly becomes one of the most promising vertebrate systems for this purpose. The GFP expression in the Tg(Δ113p53:gfp) transgenic embryos is sensitive to the DNA-damaging drugs such as camptothecin and roscovitine (Fig. 3A, B). To test whether the GFP induction in the transgenic embryos is a specific response only to the DNA damage signals, the transgenic embryos were treated with a panel of 10 model toxicants namely PbCl2, methyl-mercury chloride, As2O3, CdCl2, 4-chloroaniline, valproic acid, acrylamide, Aroclor1234, 2,3,7,8-tetrachlorodibenzo-p-dioxin, and tert-butylhydroquinone at toxic concentrations. 37 The environmental toxicity of all of these compounds is well documented. The results showed that 9 out of 10 of these toxicants induce little GFP (Fig. 3B, upper panel) and endogenous Δ113p53 expression (Fig. 3B, middle panel), except for 4-chloroaniline, which elicited a significant inductive effect. These experiments highlighted that the Tg(Δ113p53:gfp) transgenic embryos can not only be used to specifically evaluate the DNA-damaging toxicities for the environment pollutions but also be an excellent system to screen DNA-damaging reagents. This is because the transgenic embryos can sense DNA damage subtle enough that the embryos might not have other visible defects and distinguish other causes of toxicity from those caused specifically by DNA damage.
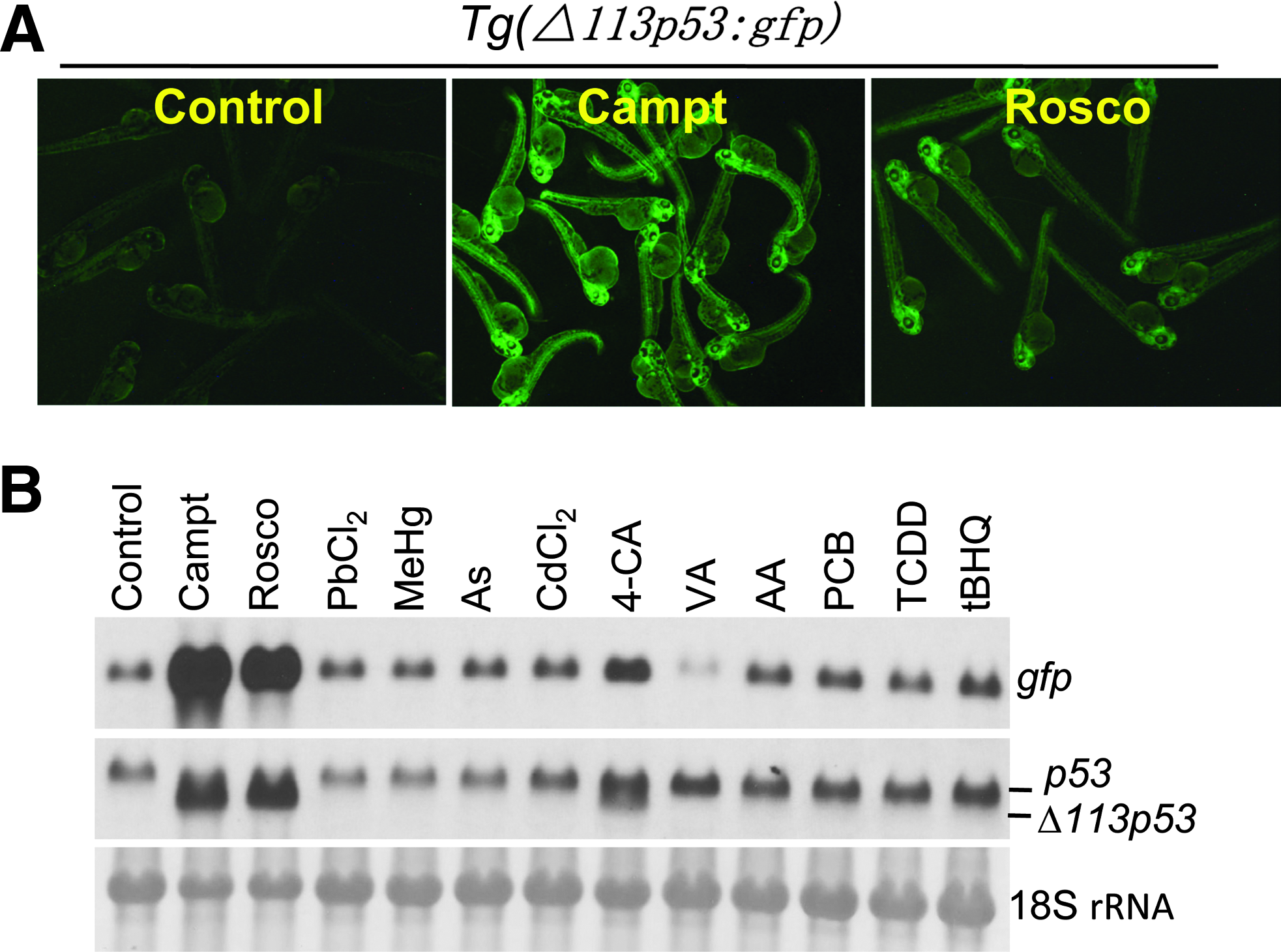
Induction of GFP and Δ113p53 expression in Tg(Δ113p53:gfp) transgenic embryos upon the treatments with DNA-damaging drugs and 10 model toxicants. Tg(Δ113p53:gfp) transgenic embryos at 1 day after fertilization were treated with 500 nM camptothecin (Campt), 50 mM roscovitine (Rosco), 2.8 mg/l PbCl2, 60 μg/l methyl-mercury chloride (MeHg), 79 mg/l arsenic trioxide (As), 5 mg/l CdCl2, 50 mg/l 4-chloroaniline (4-CA), 50 mg/l valproic acid (VA), 71 mg/l acrylamide (AA), 33 mg/l Aroclor1254 (PCB), 500 ng/l tetrachlorodibenzo-p-dioxin (TCDD), and 1.7 mg/l tert-Butylhydroquinone (tBHQ). (
Other Perspectives
The antiapoptotic activities of Δ113p53/Δ133p53 have now been demonstrated in both human cell line and zebrafish systems.15,16 Interestingly, there are reports showing that Δ133p53 is frequently overexpressed in human breast tumors and some other tumors.15,38 Do these data put forward that Δ113p53/Δ133p53 plays a role in tumorigenesis? Yet, on the other hand, coexpression of Δ113p53 with p53 in zebrafish embryos promotes the expression of not only the antiapoptotic gene bcl2L but also p21, an inhibitor of G1-to-S transition. 16 This result in turn suggests that Δ113p53 might function as a knob switching p53 activities from apoptosis promotion to cell cycle arrest. Data obtained from the zebrafish def−/− mutant also support this proposition. A loss-of-function mutation in def confers hypoplastic digestive organs and upregulates and limits the expression of Δ113p53 within these organs. This increased expression selectively induces the expression of p53-responsive genes such as p21, which in turns triggers the arrest of the cell cycle but not apoptosis, resulting in compromised organ growth in the mutant. Knockdown of Δ113p53 in the def−/− mutant significantly enhanced cell apoptosis in digestive organs causing more severe phenotypes. 16 Whether Δ113p53 acts as an oncogene or functions to switch p53 activities from promoting apoptosis to arresting cell cycle requires more direct evidence. Def is likely a nucleolar protein. Similar to the def−/− mutant, a number of genetic mutants with nucleolar or ribosomal protein deficiency, for instance, S19, 39 WDR55, 40 and L11, 41 develop abnormally through activation of the p53 protein family. It is possible that this exactly happens in the mutants of other putative nucleolar proteins with phenotypes like def−/−, for example npo−/−. 42 It would be worth to study whether loss of functions of these different genes result in common stress condition, such as protein synthesis stress, which would trigger the p53 pathway. It would also be interesting to know what roles Δ113p53 play in these different situations.
Δ113p53 retains the oligomerization domain and selectively changes p53 response gene expressions. 16 In this context, an apparent question to ask is whether Δ113p53 can form a complex with full-length p53. And if so, is such a Δ113p53/p53 heterodimer/tetramer required for Δ113p53 antiapoptotic activities or for modulating p53 transcriptional selectivity?
Gene-specific antisense morpholinos have been widely used to knockdown target gene expression in zebrafish and other systems. However, a common problem encountered by using morpholino is that morpholino injection sometimes causes undesirable phenotypes due to the nonspecific activation of the p53 pathway which is termed as “off-targeting effect” of morpholinos. 28 Activation of the p53 pathway by morpholino injection would lead to upregulation of the Δ113p53 expression as reported. 28 To our experience, although some morpholinos do not cause phenotypic abnormalities to the injected embryos, they can still induce Δ113p53 expression (Chen J and Peng JR, unpublished data). Therefore, Δ113p53 can be used as a marker to monitor both genuine and off-targeting activation of p53 pathway by morpholinos. Unfortunately, except for those genes having genetic loss-of-function mutants that can be used as references, currently there is no authentic way to distinguish between a genuine targeting and off-targeting activation of the p53 pathway in a morphant.
One interesting fact is that although there are a total of 10 p53 isoforms in human, only 1 p53 isoform has been identified in zebrafish to date.15,16,25 Do any of the other p53 isoforms exist in zebrafish? One way to address this question might require expression examination of these isoforms individually after subjecting the animal to a combination of different stress conditions.
The functions of genes in the p53 pathway, such as ATM, p53, p21, Bax, Bcl2, and BclxL, are all conserved from zebrafish to humans. 43 Studies from human cell lines showed that intron 4 of human p53 also has a promoter activity, 15 as that found in zebrafish. 16 These suggest that the findings from zebrafish system would undoubtedly provide invaluable information for human tumorigenesis and cancer therapies. The various advantages of zebrafish and the availability of transgenic fish will make the animal an excellent system for the identifications of more novel factors involved in the p53 pathway and allow screenings for antitumorigenesis drugs. Based on the exciting data summarized so far, the potential of this humble teleost to make an impact in the field of p53 should be tracked closely in the near future with great anticipation, while its role as a cancer model system should gain more extensive attention from the research community.
Footnotes
Disclosure Statement
No competing financial interests exist.