
Abstract
Abstract
Five protocols for raising zebrafish larvae were tested and compared in this study. These include four variations of feeding Fry Feed Kyowa N-250 (Kyowa) obtained from Miyako Kagaku, as well as an established protocol using Paramecia and supplemented with Zeigler dry feeds. The performance of these feeding protocols was evaluated using larvae obtained from two wild-type strains AB and AB/TL and two mutant strains gassi and smarca4a50. Several quality parameters including survival rate, rearing density, growth rate, sex ratio, fecundity, eggs spawned by the resulting fish, and the corresponding survival rate were compared by logistic regression analyses. The results showed that the quality of AB and AB/TL larvae raised using one of the Kyowa feeding protocols was equally good when compared with the standard Paramecium protocol for up to 20 larvae/L. The protocol also provided a comparable but slightly lower survival rate for smarca4a50 and inbred gassi lines, respectively. Nonetheless, all other quality parameters in the case of gassi were comparable to the established Paramecium protocol, even though the line carried multiple pigmentation mutations. Therefore, the optimized Kyowa feeding protocol can substantially reduce routine workload in maintaining a zebrafish research facility by virtually eliminating the need to culture live Paramecia feed.
Introduction
The increasing number of genetically modified fish has imposed a substantial workload on fish maintenance in major fish facilities, especially in food preparation. It is generally believed that live prey provides the best nutrients to the fish and that larval fish are particularly vulnerable. For example, Paramecium (particularly Paramecium multimicronucleatum) is a commonly used feed for raising larval fish6,12,13 and is often supplemented with dry feeds and Artemia nauplia. The routine preparation and quality control of Paramecium is not trivial. Therefore, it would be beneficial to have a convenient dry feed for raising larvae, which is comparable to Paramecium, to streamline the operation of zebrafish facilities.
Several studies have evaluated different formulations of dry feeds as a convenient means to raise larval fish14–16 and juvenile fish 17 with various success. For example, Goolish et al. compared the survival and growth rates of 10 processed diets with those of live feed, including Paramecium and Artemia at 2–3 weeks postfertilization. 14 They found that the processed feeds were generally inferior compared with the live feeds and that water flow was also a confounding factor. When the water flow was continuous, the survival rate of fish raised on dry feed was less than half (12%–19%) of those that received live food (69%). However, their growth rates were similar. When fish raised on the dry feed received a water change daily, the survival rate improved in some cases (62%) but was still lower than fish raised on live food (94%). Also, the growth rate of fish receiving a daily water change was substantially lower (14%–37%) compared with those with continuous water flow. Carvalho et al. 16 compared four commercially available, practical, and purified feeds to Artemia feed. They also used 21 days postfertilization (dpf ) survival and growth rates as criteria. One of the diets (practical diet) gave a similar survival rate (84%) compared with the Artemia feed (86%). Nonetheless, the growth of these fish was inferior (∼72%) to those fed with Artemia. Recently, Siccardi et al. investigated five commercially available and two laboratory-prepared diets in raising juvenile fish starting from 28 dpf. 17 These juvenile fish were first raised to 28 dpf on rotifers and were then randomly assigned to receive one of the seven dry feeds for an additional 9 weeks. At the end of the experiment, the authors reported survival rates ranging from 66.7% to 100% and various growth rates, depending on which feed the fish received.
Although it is essential to have an efficient feeding protocol that provides the best growth and survival rates for raising larval fish, the quality of the eggs spawned by these fish is equally, if not more, important. Therefore, an ideal dry feed should possess the following characteristics: (1) minimum preparation is required, (2) easy administration, and (3) the larvae raised using the dry feed should have comparable survival and growth rates, sex ratio, and fecundity compared with those raised using the live feed. In this study, a new dry food—Fry Feed Kyowa N-250—produced by Miyako Kagaku (Tokyo, Japan) was thoroughly tested and was found to potentially fulfill these criteria. An optimized protocol for feeding larval fish until adulthood was established.
Materials and Methods
Fish strains
Wild-type zebrafish AB and AB/TL (a hybrid of AB and TL) (http://zfin.org/cgi-bin/webdriver?MIval = aa-wtlist.apg) and the gassi and smarca4a50 lines18,19 were used in this study. The gassi line used in this study was not exactly the same transparent mutant that contained random assortments of five pigmentation mutations as originally reported 18 (personal communication with Patrick Gibbs), but was derived from a random inbreeding of these founder fish twice per year for 19 years (personal communication with Paul Collodi). The resulting adult phenotype was invariably albino. Although the exact genotype was not known, it could be assumed that the inbred gassi contained a stable number of mutations after years of inbreeding. The smarca4a50 mutation is recessive and embryonic lethal, and so the mutation was maintained as heterozygous adult fish. The heterozygous larvae and their wild-type siblings obtained from crossing these heterozygous parents were indistinguishable and were randomly mixed in the tank for the study. Hence, their ratio should be approximately 2:1.
Adult fish maintenance
Adult fish were maintained in a commercially built aquarium (Aquatic Habitats, Apopka, FL) with a recirculation water filtration system. Water was pumped through the aquarium, aerated, degassed, mechanically filtered via a 50-μm filter bag, then filtered via a 100-μm carbon filter bag, and lastly disinfected by UV irradiation. Ten percent of the system water was automatically replaced over a 24-h period with reverse osmosis (RO) water. System parameters including temperature, pH, conductivity, and dissolved oxygen were monitored by YSI probes (YSI, Yellow Springs, OH) daily. Different fish lines were maintained in individual 3-L tanks with approximately six males and six females per tank. Brood stock adult fish were fed twice a day with Artemia (Brine Shrimp Direct, Ogden, UT) and supplemented with a dry feed mixture (see next section). Fish were retained and bred in an environmentally controlled room with the photoperiod set at a 14:10-h light:dark cycle and the temperature at 28°C ± 1°C. All protocols were approved by the Purdue Animal Care and Use Committee.
Adult fish food preparation
Artemia decapsulation and hatching
Premium-grade brine shrimp eggs were obtained from Brine Shrimp Direct. One hundred and forty grams of these cysts was decapsulated by soaking in RO water (1 g/L) for 90 min with moderate aeration. Next, they were strained through a brine shrimp net, rinsed with 250 mL of prechilled 40% NaOH, and soaked in bleach (10 mL/g) with moderate aeration for about 10 min. During this step the cysts turned from gray to orange, which indicated the completion of the decapsulation process. The outer shells of the cysts became suspended in the bleach solution, which were then aspirated off, leaving only the decapsulated cysts. Finally, all cysts were thoroughly rinsed with RO water and then 250 mL of 0.6% acetic acid to neutralize any remaining bleach. All decapsulated cysts were stored in 500 mL of 32 parts per thousand Instant Ocean (Aquarium Systems, Mentor, OH) salt solution (brine solution) at 4°C for up to 2 weeks.
For hatching, 30 mL of decapsulated Artemia cysts from the stock solution was resuspended in 4 L of fresh brine solution in a hatching cone (Aquatic Habitats) with constant aeration. After 24 h, half of the hatched cysts were drained from the hatching cone through a brine shrimp net for the morning feeding at 8:30 am. They were rinsed with RO water and put into a 500-mL Nalgene wash bottle. The remaining half of the hatching cone was used in the afternoon feeding session at 3:30 pm in the same manner.
Adult dry feed supplement
The adult dry feed supplement was based on a recipe from the Zebrafish International Resource Center (http://zebrafish.org/zirc/documents/protocols.php#Fish%20Feeding). It was a mixture of 2000 g of Zeigler Zebrafish Granule (Aquatic Habitats), 500 g Spirulina flake (Drs. Foster and Smith, Rhinelander, WI), 200 g Cyclopeeze (Custom Aquatics, Carlsbad, CA), 8 oz Golden Pearl 300–500, and 8 oz Golden Pearl 500–800 (Brine Shrimp Direct). Approximately 6.5 g of this mixture was added into the Nalgene wash bottle with the hatched Artemia, which was then filled with RO water. This amount of food was used to feed approximately 2000 adult fish in the system and was dispensed to each tank until the fish were fed to satiation. In our case, this required approximately three to eight drops, depending on the density of the fish in the tank.
Egg production
The night before breeding, one male and one female fish were placed in a common 2-L static breeding tank (Aquatic Habitats, etc.) with system water, separated by a divider. On the day of breeding, fish were moved into a fresh tank of system water as soon as the room light was on. The divider was then removed and the fish were allowed to breed for up to 3 h. After breeding, the adults were put back into their system tank and the embryos were collected. About 60 embryos were maintained in a 100-mm Petri dish with E3 media, 6 which was changed daily to avoid metabolic waste accumulation and to ensure consistent water quality. All embryos obtained from an individual breeding pair were maintained at 28°C and inspected daily under a stereomicroscope until 5 dpf to allow for the selection of healthy embryos for subsequent analyses.
Larval fish maintenance
The nursery system used in this study was the 1-L Library System from Aquatic Habitats. As it was connected to the water circulation of the adult fish system as described earlier, it was believed that the water quality would not be different when regular system tanks were used to raise larvae. However, using 1-L tanks can maximize the space usage, which is often the concern of many research laboratories. The embryos spawned by a pair of parents that had more than 80% survival rate and were morphologically normal at 5 dpf were raised in this nursery system. All embryos from different parents that met the criteria were combined in equal ratio in all 1-L tanks, typically 10 or 20 fish per tank, for the subsequent food studies. To transfer the embryos from a Petri dish to a nursery tank, 1/3 of the tank was filled with system water before embryos were pipetted into the tank. The food and the water flow that each tank received depended upon the feeding protocol assigned to that tank, which will be described in subsequent sections.
Larval food preparation
Paramecium feed and supplements
In this study, the Paramecium feed was a mixture of Paramecium multimicronucleatum (Carolina Biological Supply, Burlington, NC) and commercially available dry feeds: Zeigler Larval AP100 Supplements 100–150 microns (Zeigler 100–150 μm) and 250–400 microns (Zeigler 250–400 μm) (Zeigler Bros, Gardners, PA). The Zeigler feeds generally contain >50% protein, >12% fat, <9% ash, and <2.5% fiber. The Paramecium culture was set up using the following procedures: First, a quarter cup of wheat seeds was boiled in 500 mL of RO water for 15 s and then cooled to room temperature. Then, 10–12 seeds were placed in a 750-mL Gladware container with 500 mL of autoclaved system water and mixed with 10 mL of the Paramecium stock culture. This culture would be ready to be used the next day. Before each feeding, a few drops of the culture were examined under a stereomicroscope to evaluate the health of the Paramecia. The detailed sequence of food administration is described in the next section.
The Zeigler dry feed supplements were administered by scooping an appropriate amount from the storage bottle. In 30 repeated measurements, the mean weights (μ) and the standard deviations (SDs) were determined (μ = 5.64 mg, SD = 1.30 for Zeigler 100–150 μm; μ = 8.46 mg, SD = 1.31 for Zeigler 250–400 μm).
Kyowa feed
Fry Feed Kyowa N-250, or Kyowa Feed, was obtained from Miyako Kagaku. A purchase order was placed directly to the company using the following information:
Name: Miyako Kagaku Co. Ltd. Address: 4-1, Marunouchi 3-chome, Chiyoda-ku, Tokyo 100-0005 Bank: Bank of Tokyo-Mitsubishi UFJ Account number: 3051192 Swift code: BOTKJPJT Contact person: Y. Harada (
The feed contains >55% protein, >10% fat, <17% ash, and <4% fiber and has a particle size of <250 μm (see Supplemental Material 2 for a pamphlet on Kyowa [available online at www.liebertonline.com; available only in Japanese]). A small portion of Kyowa Feed was kept in a 100 × 15 mm Petri dish and stored at 4°C for routine feeding. Kyowa was dispensed to each tank with a 7-mL disposable transfer pipette with a 3.2-mL bulb draw (Dot Scientific, Burton, MI). The pipette was dipped into the Petri dish and the collected food was puffed into the tank. This was defined as 1 puff of Kyowa feed (1 puff ). If the dipping and puffing actions were repeated twice, it was defined as 2 puffs of Kyowa feed (2 puffs). The amounts of thirty 1 puff and 2 puffs feed were weighed and the mean weights (μ) and SDs were 4.04 mg and 0.57 for 1 puff, and 9.62 mg and 0.92 for 2 puffs, respectively.
Larval feeding protocols
Five feeding protocols were tested and compared in this study. They were all based on the Paramecium and Kyowa feeds described in the previous section.
Paramecium feeding protocol
The Paramecium feeding protocol (Paramecium) was the feeding regime based on the Paramecium feed. It was based on a protocol from the Zebrafish International Resource Center (http://zebrafish.org/zirc/documents/protocols/pdf/Fish_Nursery/Nursery/Detailed_Nursery_and_Food_Instructions.pdf ) and was the control for all experiments. Table 1 shows the detailed descriptions of the administration procedure and the associated changes in water flow in the nursery tanks. Approximately 1 mL of the culture was fed twice a day, once at 8:30 am and once at 3:30 pm. Various Zeigler dry foods were also administered at appropriate stages as described.
Embryos were maintained in Petri dish.
Larvae were transferred to 1-L nursery tank on day 6.
Juvenile fish were transferred to the adult system.
dpf, days postfertilization.
Kyowa feeding protocols
Four variations of feeding regime based on the Kyowa feed were tested (Table 2). In the first two kinds (1 puff Kyowa without transition [1 puff no trans] and 2 puffs Kyowa without transition [2 puffs no trans]), the food source was changed directly from Kyowa to Artemia on day 14; in the last two kinds (1 puff Kyowa with transition [1 puff & trans] and 2 puffs Kyowa with transition [2 puffs & trans]), the Kyowa was fed in conjunction with Artemia starting on day 14 until switching to the adult diet at 45 dpf. In all conditions, the Kyowa feed was administered twice per day, once at 8:30 am and once at 3:30 pm. The date of the change in the water flow was the same for all feeding variations; the only difference was the length of time of Kyowa food administration.
Embryos were maintained in Petri dish.
Larvae were transferred to 1-L nursery tank on day 6.
Juvenile fish were transferred to the adult system.
Fish length measurement
Juvenile fish at 60 dpf were anaesthetized with 0.2 mg/mL Tricaine (Sigma, St. Louis, MO) 6 and placed in a Petri dish for measuring body length, defined as the length from the tip of the mouth to the caudal peduncle. Two alternative ways were used for measuring this length. First, a silk suture thread was placed on top of the fish to follow the curvature of the body and cover this length. This thread was cut and placed with the whole fish under an Olympus SZX16 Stereomicroscope (Olympus, Center Valley, PA). Images were taken by a Spot RT3 slider CCD camera (Diagnostic Instruments, Sterling Heights, MI). The length of the thread and the body length of the fish were measured by iSolution image analysis software version 8.4 (IMT i-Solution, Vancouver, BC, Canada). In two independent experiments, the Pearson correlation coefficients for the length of the same group of fish measured by two different measuring methods were 0.95 and 0.97, and the corresponding p-values of the Welch's modified two-sample t-tests 20 were 0.61 and 0.18. These results suggested that both measurements were comparable. The thread measurements were subsequently used for wild-type analyses, whereas the image measurements were used for the gassi analysis.
Fluorescence profile of various larval foods in vivo
Larvae fed with different larval foods were anaesthetized at 8 dpf as previously described, 6 placed laterally in 3% methylcellulose, and observed under an Olympus SZX16 Stereomicroscope (Olympus) with a X-Cite 120 fluorescence illuminator (Exfo Life Sciences, Ontario, Canada). The fluorescence profile in the green fluorescence channel (i.e., GFP) was inspected and imaged by a Spot RT3 slider CCD camera (Diagnostic Instruments). Same imaging settings were used for acquiring all images, which were then analyzed by iSolution image analysis software version 8.4 (IMT i-Solution). First, six 100 × 100 pixels areas at the same empty location were selected as background references in all images. Second, fluorescing region of interest, which was presented exclusively in the gut area, was manually selected. The mean fluorescent intensities were extracted from these areas. The average of the mean background intensities was subtracted from the region of interest in the same image to obtain the fluorescent intensity for data analysis.
Statistical analysis
All statistical analyses were performed in the R statistical environment (http://www.r-project.org) version 2.9.0. Logistic regression models21,22 were built to evaluate the potential effect of larval feeding protocol change (food) and other applicable variables such as the survival rate, sex ratio, or hit rate of breeding when the fish reached adulthood (for a step-by-step example, see Supplemental Material 1 [available online at www.liebertonline.com]: A primer for the logistic regression approach used in this study). A stepwise procedure was used to identify the most parsimonious model. The significance of the added variable was calculated by analysis of deviance of the nested models. The overall model significance was evaluated by deviance goodness-of-fit test. For the body length and in vivo food fluorescence data, the homogeneity of the variances between treatment groups was evaluated using Bartlett's test. 23 Differences in means of body length and in vivo food fluorescence data were evaluated by Welch's modified F statistic 20 and ANOVA, respectively. Post hoc comparisons were done using Bonferroni corrected t-test. For testing the potential correlation between the survival count of the embryos at 5 dpf and the total number of embryos spawned by the parents, the Spearman's rank correlation ρ was used. An alpha level of 0.05 was used for all statistical tests.
Results
Survival rate of different feeding protocols
Thirty AB/TL larvae were assigned to each of the five feeding protocols (Tables 1 and 2) and divided into 10 fish per 1-L tank. Therefore, three technical replicates were performed for each feeding protocol. The idea of the Kyowa with transition protocols was conceived after preliminary experiments showing that the Kyowa feeding protocols without transition had a comparable survival rate to Paramecium protocol but an inferior growth rate (data not shown). It was hypothesized that by feeding Kyowa in conjunction with Artemia, the fish would grow to a comparable size of the Paramecium-fed fish.
Survival counts were performed on individual tanks every day until 45 dpf. The 14 and 45 dpf survival counts and rates are shown in Table 3. The reasons for performing the analysis at these 2 days are as follows: (1) because the larvae received Kyowa exclusively until 14 dpf, checking the survival statistics at this time would allow a direct comparison with the control Paramecium protocol; (2) because the Kyowa feeding protocol ended at 45 dpf, the survival count would show the effect of these feeding protocols on overall survival. To evaluate the influence of the feeding protocol on fish survival, logistic regression was performed with the change of the feeding protocol (food) as the independent variable for the 14 and 45 dpf data. The results of the analysis of deviance are shown in Supplemental Table S1 (available online at www.liebertonline.com). The p-values for the addition of the food variable in the 14 and 45 dpf models were 0.12 and 0.74, respectively. Also, the p-values of the deviance goodness-of-fit tests of the null models at both stages were 0.42 and 0.49, suggesting the null models were satisfactory fits. The results suggest that a change in feeding protocol did not alter the survival rate of the fish at both stages.
Three tanks were set up for each feeding condition and 10 larvae were put in each tank. See Supplemental Table S1 for the analysis of deviance.
As stated earlier, it was noted that fish fed with the Paramecium protocol had grown considerably larger than both Kyowa feeding protocols without transition (1 puff no trans and 2 puffs no trans) by the end of 45 dpf, indicating that these two Kyowa protocols were inferior in terms of growth rate. One obvious reason was that the larvae did not receive as much food compared with those fed by the Paramecium protocol (i.e., no additional dry food when Artemia was fed; the growth data will be discussed later) (Tables 1 and 2). However, the growth of the fish fed by the Kyowa with transition protocols (1 puff & trans, and 2 puffs & trans) appeared to be comparable to the fish fed by the Paramecium protocol. This indicates that the Kyowa protocols with transition were potentially better than the no-transition counterparts and might potentially be equivalent to the Paramecium protocol. To fully investigate this, additional studies were conducted and will be elaborated in the following section.
Rearing density
If larvae could be reared at a higher density per tank, it would save space and time to feed. To investigate this, Kyowa feeding protocols 1 puff and 2 puffs with transition and Paramecium feeding protocol were used to rear twice as many (20) larvae per 1-L tank. Again three technical replicates (tanks) were used for each feeding protocol. The whole experiment was repeated on different days using two different wild-type strains, AB/TL and AB.
The survival counts were again performed on individual tanks every day until 45 dpf. The 14 and 45 dpf survival counts and rates are shown in Table 4A. Logistic regression was performed with the change in the feeding protocol (food) and the fish strain (strain) as the independent variables for the 14 and 45 dpf data. The results of the analysis of deviance are shown in Supplemental Table S2A (available online at www.liebertonline.com). For the 14 dpf models, the p-values for the addition of food and the strain variables were 0.59 and 0.0024, respectively. Therefore, the best-fit 14 dpf model was “Intercept + strain.” This was also supported by the model's insignificant deviance goodness-of-fit test p-value of 0.26. For the 45 dpf models, the p-values for the addition of food and the strain variables were 0.019 and 0.89, respectively, indicating the “Intercept + food” was a better-fit model. However, its deviance goodness-of-fit test p-value was 0.019, suggesting the model was not a good fit and there were other factors affecting the final survival rate. The lack of association between the strain and the survival rate at 45 dpf also suggests that the difference observed at 14 dpf was probably by chance or other factors. The fitted 45 dpf “Intercept + food” model showed that the fish fed with the 1 puff & trans protocol had a slightly better overall survival rate compared with the other two feeding protocols (p-value = 0.095). Therefore, these results indicate that the change in feeding protocol did not decrease the survival rate of the fish up to 20 larvae/L.
Three tanks were set up for each feeding condition and 20 larvae were put in each tank. See Supplemental Table S2 for the analysis of deviance.
Growth rate
After determining that the 1 puff & trans protocol was as good as the Paramecium protocol in terms of survival rate, its effect on growth, sex ratio, fecundity, and the quality of eggs spawned by the fish raised using the protocol were determined. The length of the AB fish used in the rearing density experiment was measured at 60 dpf and the resulting data are plotted in Figure 1. As the variances were significantly different from each other (Bartlett's test of homogeneity of variances, p-value = 0.0055), the difference in means for fish length raised using different feeding protocols was evaluated by Welch's modified F statistic. 20 The analysis shows that there was a significant effect on fish length by the change in the feeding protocol: F(2, 82.809) = 5.3677, p-value = 0.0064. Post hoc comparisons using Bonferroni corrected t-test show that the mean body length of fish raised using the 2 puffs & trans protocol (μ = 1.78 cm, SD = 0.35) was significantly longer than those raised using the Paramecium (μ = 1.56 cm, SD = 0.22) and 1 puff & trans (μ = 1.60 cm, SD = 0.31) protocols (Bonferroni adjusted p-values = 0.0035 and 0.013, respectively). The Paramecium and 1 puff & trans protocols, however, were not significantly different from each other (Bonferroni adjusted p-value = 1). Together these results suggest that the growth of the fish raised using the 1 puff & trans protocol was comparable to those raised using the Paramecium protocol at 60 dpf. As all fish were fed an adult diet after 60 dpf, this also strongly suggests that this protocol can support growth as well as the Paramecium protocol. Fish raised using the 2 puffs & trans protocol were larger than the other two protocols. This was likely due to the fact that fewer of these fish survived (73.33% for the 2 puffs & trans protocol versus 88.33% and 88.67% for the Paramecium and the 1 puff & trans protocols, respectively; Table 4), allowing the remaining individuals to consume more food and thus grow faster.
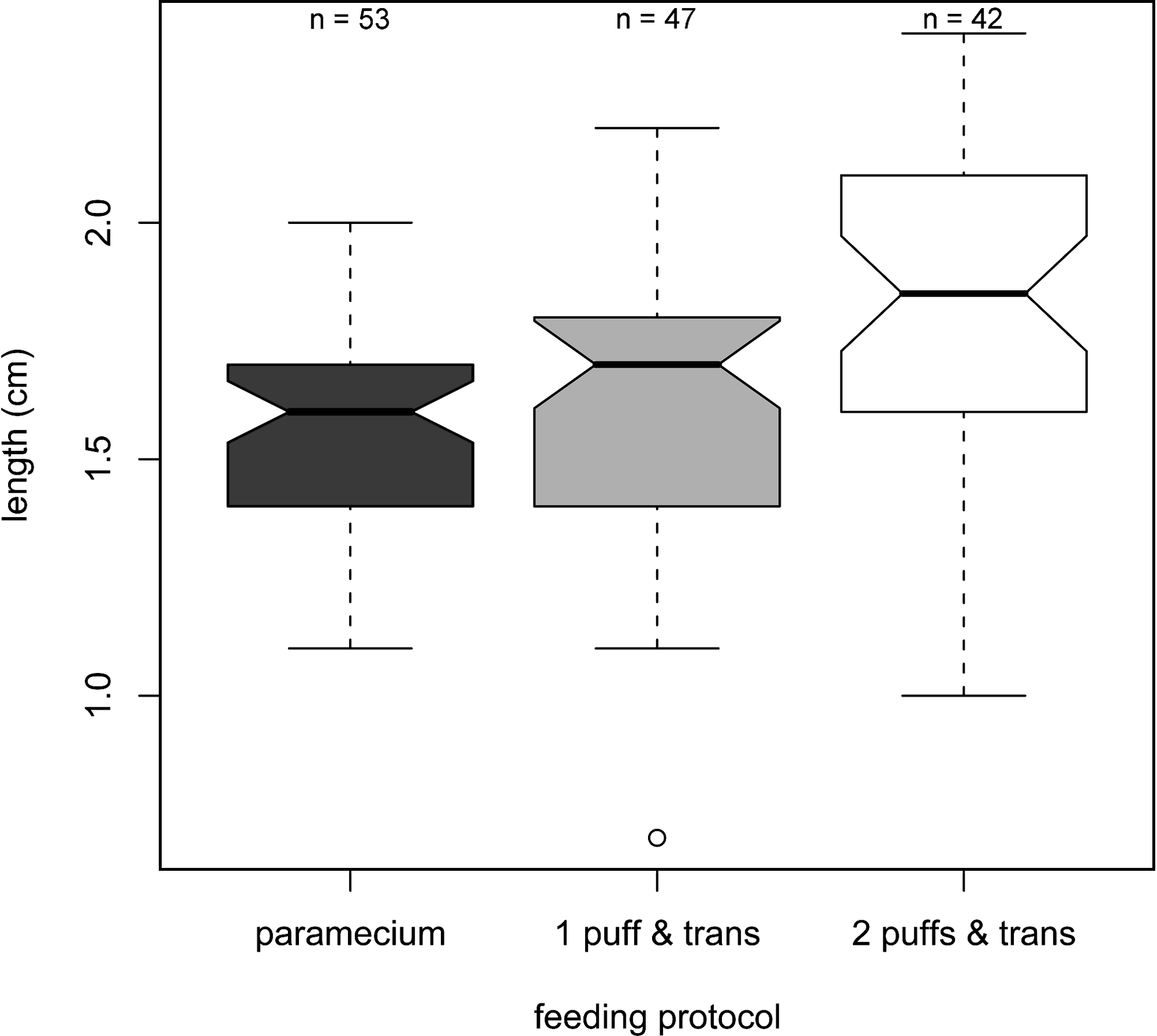
Length of AB fish raised using different feeding protocols at 60 dpf. A boxplot21,28 showing the length of the fish raised using Paramecium, 1 puff & trans, and 2 puffs & trans protocols. These were the same fish used for the analysis shown in Table 4. The thick black lines in the boxes indicate the medians of the individual groups. The two hinges of the boxes are the first and third quartile, the notches extend to ±1.58 IQR/square root (n), and the whiskers show the largest/smallest observation(s) that falls within a distance of 1.5 times the box size from the nearest hinge. All values that fall outside this range are considered outliers and are plotted separately.
Sex ratio
To compare the sex ratio of the fish raised using different feeding protocols, the sex of each fish was determined based on morphology (Table 4B). Logistic regression was performed with the change in the feeding protocol (food) and the fish strain (strain) as the independent variables. The results of the analysis of deviance are shown in Supplemental Table S2B (available online at www.liebertonline.com). The p-values for the addition of food and the strain variables were 0.99 and 0.67, respectively. Also the p-value of the deviance goodness-of-fit test of the null model was 0.83, suggesting the null model was a satisfactory fit. It was noticed that more males were present in all conditions than females. Although it was not clear what caused the overabundance of males, it might be related to rearing density or other uncharacterized parameters in the fish water. Nonetheless, these results show that the change in feeding protocol and the difference in strain did not affect the sex ratio.
Fecundity and embryo health
One major advantage of using zebrafish as a research model is that they can produce a large number of embryos at regular intervals. To find out whether fish raised using different feeding protocols could regularly produce a high quality and quantity of embryos, five pairs of sexually mature males and females that were at least 4 months old were selected randomly from the AB/TL and AB experiments as described in Table 4A and B. Individual pairs were separated into 3-L tanks and bred once every week for 3 h for five consecutive weeks. A successful breeding event was defined as embryos being spawned by a pair of parents and these embryos were not obviously degraded. The number of successful breedings over five trials is shown in Table 4C. The effect of the change in the feeding protocol on breeding success was studied by logistic regression analysis with the change in the feeding protocol (food) and the fish strain (strain) as the independent variables. The results of the analysis of deviance are shown in Supplemental Table S2C (available online at www.liebertonline.com). The p-values of all variables in the full model “Intercept + food + strain + food*strain” were significant, suggesting both variables and their interaction significantly affected the breeding success. The deviance goodness-of-fit test had a significant p-value (0.0019), suggesting that the model was not the best fit. However, even though there were other additional factors affecting a successful breeding, the current analysis still indicates that the change in feeding protocol affected the breeding success. In the logistic regression model, both coefficients for 2 puffs ( foodpuff_2; see Primer for the logistic regression for syntax and analysis output) and 2 puffs interaction with AB/TL ( foodpuff_2:strainTLAB) were significant (p-values = 0.00038 and 0.0093, respectively). The estimates of these two coefficients are −3.932 and 3.258, respectively. The remaining variables in the model were not significant. These data suggest that (1) the 1 puff & trans protocol did not change the breeding success compared with the Paramecium protocol, and (2) the 2 puffs & trans protocol significantly decreased the breeding success compared with the Paramecium protocol and this effect was more severe for the AB strain.
All embryos obtained from the breeding exercise were observed under the microscope daily and the number of healthy and unhealthy embryos were counted until 5 dpf. The total survival counts for each feeding protocol for each strain are shown in Table 4D. The survival counts and rates of the embryos collected from the individual breeding events are shown in Figure 2A and B, respectively. The total number of eggs that could be collected during the experimental period was similar between the Paramecium and the 1 puff Kyowa & trans protocols (2071 vs. 2078 for AB/TL and 2216 vs. 2356 for AB), whereas there were obviously fewer embryos produced by the fish raised using the 2 puffs & trans protocol (1359 and 315 for AB/TL and AB, respectively). To fully evaluate the effect of change in the feeding protocol on the survival of the embryos, a logistic regression analysis was performed using the change in the feeding protocol (food) and the fish strain (strain) as the independent variables. The results of the analysis of deviance are shown in Supplemental Table S2D (available online at www.liebertonline.com). The addition of the food variable was highly significant (p-value = 2.94 e − 57), whereas the addition of the strain variable was not (p-value = 0.2). The deviance goodness-of-fit test of the “Intercept + food” model showed that it was not a good fit (p-value = 0). However, the analysis still indicated that the change in the feeding protocol significantly affected the fecundity. In the logistic regression model, the 1 puff & trans coefficient was significant (p-values <2 e − 16), whereas the 2 puffs & trans variable was not (p-values = 0.68). The estimate of the 1 puff & trans coefficient was 0.7722, suggesting that this protocol gave a better survival rate compared with the Paramecium protocol. This was likely contributed by the superior survival results from the AB fish (Fig. 2A, B). Also, there was no correlation between the survival count of the embryos at 5 dpf and the total number of embryos spawned by the parents. This was supported by the insignificant p-values of Spearman's rank correlation ρ of the Paramecium, 1 puff & trans, and 2 puffs & trans protocols for AB/TL (0.45, 0.63, and 0.63) and for AB (0.18, 0.42, NA; there was only one observation for AB raised using the 2 puffs & trans protocol). Together with a reasonably high individual survival percentage (Fig. 2B), these data suggest that the number of embryos spawned by a pair of fish was not a good indicator of the resulting survival rate and health. Also, the fecundity of the fish raised using the 1 puff & trans protocol and the survival rate of the embryos spawned by these fish were comparable to, if not better than, the Paramecium protocol.
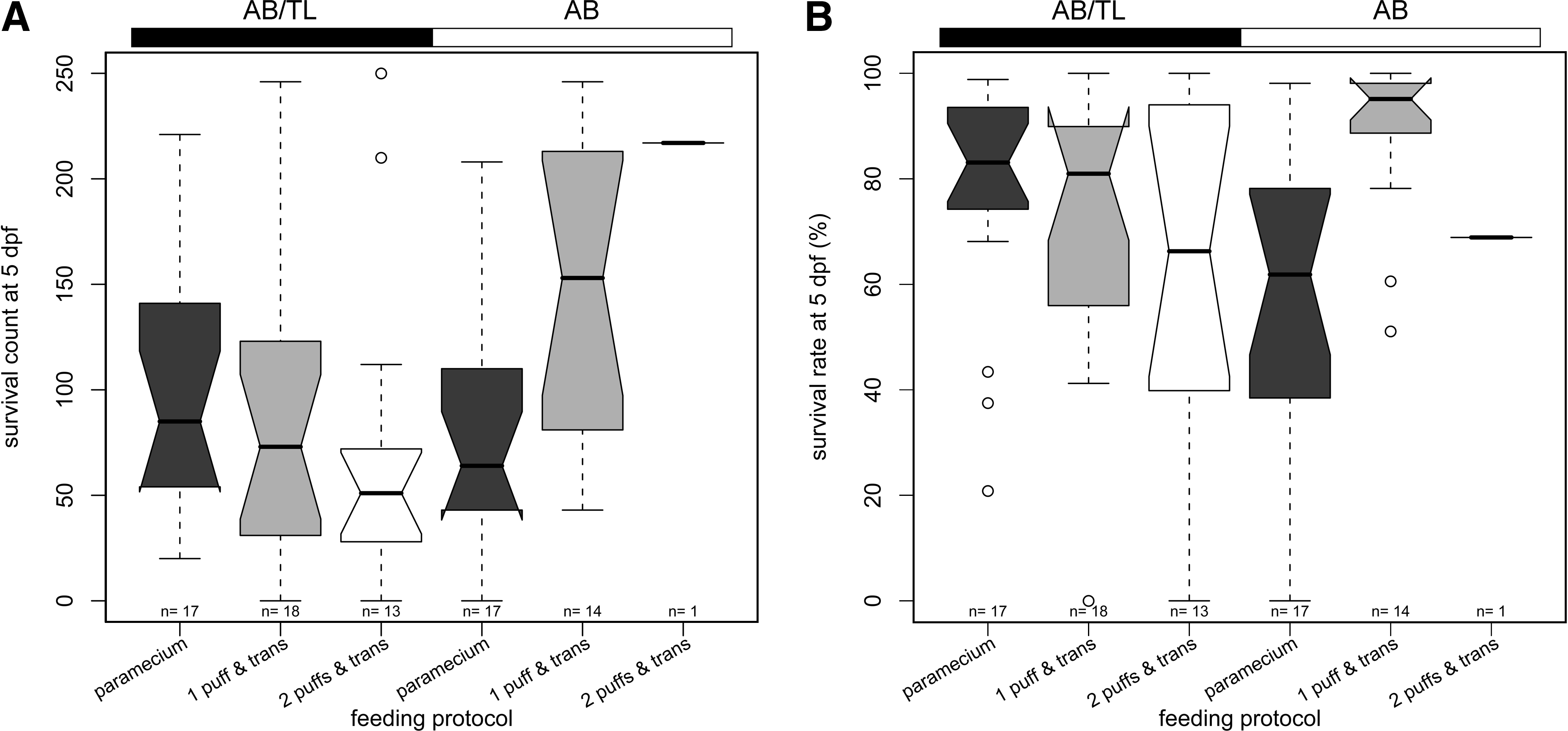
Survival counts (
Raising mutant fish
Many mutant and transgenic fish are being used in research laboratories. They may not be as fit as wild-type strains because of the mutation or genetic modification. To investigate the suitability of the 1 puff & trans protocol in raising mutant lines, two mutant lines were chosen and raised using this protocol. The first line was an inbred gassi line. These embryos are historically difficult to raise. The second line was a smarca4a50 line that had been used in our research for years 24 and had been raised successfully in the facility (see Materials and Methods section for an in-depth discussion of these lines and their genotype). Together they would serve as a reference of the effectiveness of the feeding protocol in raising mutant embryos.
Ten inbred mutant gassi and 20 smarca4a50 larvae (with a heterozygous carrier to wild-type ratio of 2:1) raised using the 1 puff & trans protocol were compared with their siblings and wild-type AB counterparts raised using the Paramecium protocol, respectively. The survival counts of the two experiments are shown in Table 5A. Logistic regression analysis was performed with the change in the feeding protocol (food) and the mutation status (mutation) as the independent variables whenever applicable. The results of the analysis of deviance for gassi and smarca4a50 are shown in Supplemental Table S3A and B (available online at www.liebertonline.com), respectively. The change in the feeding protocol to 1 puff & trans decreased the survival rate of the gassi fish from 100% to 70% (p-value = 0.0002). The p-value for the deviance goodness-of-fit test of the “Intercept + food” was 0.91, suggesting that this model was a good fit. Therefore, the survival of the gassi fish raised using the 1 puff & trans protocol was inferior to those raised using the Paramecium protocol. Interestingly, both the change in the feeding protocol and the mutation did not affect the survival rate of smarca4a50 compared with wild-type (89% vs. 90.83%; p-values of the food and mutation variables were 0.81 and 0.46, respectively). Therefore, it was concluded that the 1 puff & trans protocol did not affect the survival rate of smarca4a50.
For gassi experiments, three tanks were set up for each feeding condition and 10 larvae were put in each tank. For smarca4a50 experiments, five tanks were set up and 20 larvae were put in each tank. These data were compared with the wild-type AB/TL and AB raised using Paramecium protocol as described in Table 4.
See Supplemental Table S3 for the analysis of deviance.
For smarca4a50 raised using 1 puff & trans protocol, the survival counts were collected at 48 dpf.
Wild-type AB/TL.
Wild-type AB.
The gassi fish were then characterized to see if the change in the feeding protocol to 1 puff & trans affected other qualities of the fish. First, the growth of the fish was analyzed by body length measurement. The means of the gassi fish body length raised using the Paramecium and 1 puff & trans protocols were 1.79 cm, SD = 0.15 and 1.88 cm, SD = 0.23, respectively. The p-value of Welch's modified two-sample t-test 20 shows that there was no difference between the mean length of the two groups (p-value = 0.13). There were fewer fish in the group fed by the 1 puff & trans protocol. Based on the observation from the wild-type growth rate analysis (Fig. 1), it would be reasonable to expect that they could grow larger than the Paramecium group. The lack of difference here can be explained by the lower starting larval density (10) and the food supply was probably not a limiting factor. Second, the sex of the gassi fish raised using these two feeding protocols was recorded (Table 5B). The Pearson chi-squared test of the sex counts gives a p-value of 0.33, suggesting that the change in feeding protocol did not significantly alter the sex ratio.
Then five pairs of 4-month-old sexually mature males and females were randomly selected from the two feeding groups. These fish were again kept separated as individual pairs in 3-L tanks and allowed to breed once every week for 3 h for five consecutive weeks. Similar analyses of the fecundity and embryos' health as in the case of wild-type were performed. First, the breeding hit rate over 5 weeks was recorded (Table 5C) and analyzed by logistic regression with the change in the feeding protocol (food) as the independent variable. The results of the analysis of deviance are shown in Supplemental Table S3C (available online at www.liebertonline.com). The p-values for the addition of food variable and the deviance goodness-of-fit test were 1.00 and 0.51, respectively, suggesting that the change of food did not significantly affect the breeding success and the null model was a satisfactory fit. Second, the embryos spawned by these adults raised using different feeding protocols were observed until 5 dpf. The survival counts and rates of the embryos collected from the individual breeding events are shown in Figure 3A and B, respectively. Fewer eggs were spawned by the fish raised using the 1 puff & trans protocol compared with the Paramecium protocol (802 vs. 1120; Table 5D) during the experimental period, probably because of one breeding event in the Paramecium group that produced a substantial number of eggs (more than 350; Fig. 3A). The effect of the change in the feeding protocol on the survival of the embryos obtained was analyzed by logistic regression analysis using the change in the feeding protocol (food) as the independent variable. The analysis of deviance results are shown in Supplemental Table S3D (available online at www.liebertonline.com). The p-value of the addition of the food variable was 0.11, suggesting that the change in the feeding protocol did not change the survival rate of the spawned embryos. Nonetheless, the deviance goodness-of-fit test for the null model gave a very small p-value of 2.84 e − 96, suggesting that there were other factors affecting the survival of the fish.

Survival counts (
The results from the gassi and smarca4a50 experiments suggest the following points: (1) Different mutant strains may have different sensitivity to the Kyowa food. In this study, the survival rate of the smarca4a50 larvae, a line that was known to be easier to raise, was not affected by the feeding protocol, whereas the inbred gassi, a line that was known to be difficult to raise, had a lower survival rate. (2) Despite having a lower survival rate, the inbred gassi fish raised using the 1 puff Kyowa with transition protocol had a similar growth rate and breeding success rate and the spawned eggs had a similar 5 dpf survival rate compared with those siblings raised using the Paramecium protocol.
Long-term survival of second-generation fish
The results indicate that wild-type and mutant parents (first generation; F1) raised using the 1 puff & trans protocol could spawn second-generation embryos that had equally good 5 dpf survival rate when compared with those raised using the Paramecium protocol. The results also suggest that these second-generation (F2) embryos raised using the 1 puff & trans protocol can be efficiently used for genetic analysis of early embryogenesis. However, the experiment did not investigate the effect of the protocol on the long-term survival of these F2 embryos. To investigate this potential effect, survival data of F2 fish raised using the 1 puff & trans protocol from routine breeding were extracted from the facility database (Hensley et al., in prep.) (Table 6). These fish were maintained the same way as described in the Materials and Methods section, except (1) their parents were raised using the 1 puff & trans protocol, and (2) the counting was not done on the individual tanks, but was on the whole batch of embryos coming from the same breeding exercise. Two sets of breeding fulfilled these criteria, one was a breeding of wild-type AB and the other was smarca4a50. Both breedings were conducted in a way analogous to the analyses mentioned earlier. The counting was first done at 5 dpf, when the larvae were put into the nursery system and started to be fed with 1 puff & trans protocol, and at 60 dpf, when the juveniles were transferred to the adult system and started to be fed with the adult diet.
Two breeding exercises in which both F1 and F2 generations were raised using 1 puff & trans protocol. The smarca4a50 F2 generation came from the intercross of the two F1 generations (# and *), both of which were raised using 1 puff & trans protocol.
See Supplemental Table S4 for the analysis of deviance.
To evaluate the potential effect of 1 puff & trans protocol on the long-term survival rate of the F2 fish, a logistic regression analysis was performed using the generation of the fish (gen) and the fish strain (strain) as the independent variables for the survival count data. The results of the analysis of deviance are shown in Supplemental Table S4 (available online at www.liebertonline.com). The p-values for the addition of the gen and the strain variables were 0.103 and 0.528, respectively. Therefore, the survival rate did not change between generations or strains. It was noticed that the p-value of the deviance goodness-of-fit test of the null model was 2.06 e − 12, suggesting that it was not a satisfactory fit and there were other factors that affected the survival rate. Nonetheless, these results show that the 1 puff & trans protocol gave a consistent long-term survival rate of the F2 fish compared with the F1.
Fluorescence profile of various larval foods in vivo
Dry food generally fluoresces, including the ones used in this study (data not shown). To investigate the unwanted fluorescence effect in the larvae fed with dry food, 10 larval fish were fed with nothing (control), Paramecium only, Paramecium + Zeigler, or Kyowa only from 5 to 8 dpf. At 8 dpf, these fish were imaged within 1 h of feeding to investigate the fluorescence profile of the food in vivo. The fluorescence signal was generally observed in the gut region (data not shown) and was extracted from the images. Their log10 values were plotted (Fig. 4). As the variances of the four feeding groups were not significantly different from each other (Bartlett's test of homogeneity of variances, p-value = 0.3952), the difference in means of the fluorescence signals was evaluated by ANOVA. The analysis shows that there was a significant effect on the fluorescence intensity in fish fed by different food: F(3, 35) = 12.803, p-value = 8.382 e − 6. Post hoc comparisons using Bonferroni corrected t-test show that the larvae fed with Paramecium + Zeigler had a significantly stronger fluorescence than the control, Paramecium only, and Kyowa only groups (Bonferroni adjusted p-values = 0.0013, 4.1 e − 6, and 0.0193, respectively). As the Paramecium only condition has the lowest fluorescence, the additional fluorescence must come from the Zeigler feed, which was also supported by imaging of the food itself (data not shown). Although the fluorescence in those larvae fed with Kyowa food was significantly higher than those fed with Paramecium only (Bonferroni adjusted p-values = 0.0328), it was not significantly different from the larvae fed with nothing (Bonferroni adjusted p-values = 1). Also, the fluorescence in those larvae fed with only Paramecium was not significantly different from those fed with nothing (Bonferroni adjusted p-values = 0.359).
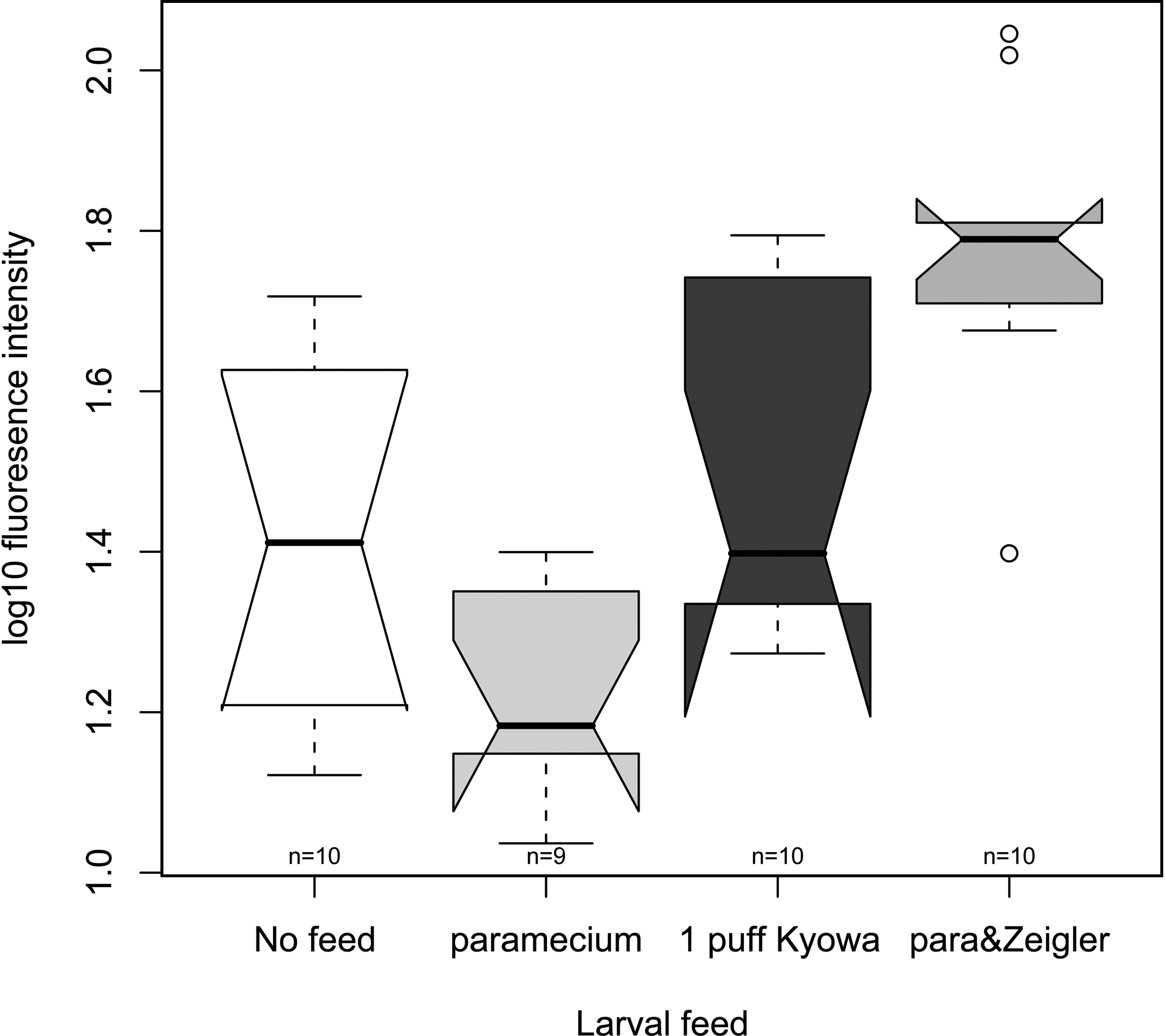
Fluorescence profile of various larval foods in vivo at 8 dpf. Each group of larvae was fed with nothing, Paramecium only (Paramecium), 1 puff Kyowa, and Paramecium +Zeigler (para&Zeigler) feed from 5 to 8 dpf. The log10 fluorescence intensity was acquired from the images of the larvae as described in the Materials and Methods section. The thick black lines in the boxes indicate the medians of the individual groups. The convention of the boxes is the same as in Figure 1.
Discussion
This study successfully established a convenient procedure for raising zebrafish larvae on a Kyowa dry feed. The aim was to establish a convenient dry feed protocol to eliminate the need of Paramecium culture for routine maintenance. Therefore, the comparison was made at the protocol level rather than the food level. The most comparable protocol was found to be 1 puff Kyowa with transition for up to 20 larvae per 1-L tank (Table 2). For two wild-type strains AB/TL and AB, this protocol provided a comparable survival rate (Table 4A and Supplemental Table S2A), growth rate (Fig. 1), sex ratio (Table 4B and Supplemental Table S2B), breeding success rate (Table 4C and Supplemental Table S2C), fecundity, and resultant egg quality (Fig. 2, Table 4D and Supplemental Table S2D) to their siblings raised using the Paramecium protocol. The long-term survival rate in the second-generation fish raised using this established protocol was also comparable to the first generation (Table 6 and Supplemental Table S4).
In particular, the survival rate of the larvae raised using the optimized Kyowa protocol was at least 85% for the two wild-type fish strains tested. The introduction of Artemia at 14 dpf in our protocol obviously contributed to this high survival and growth rates (similar to live feed control) of the fish compared with the study by Goolish et al., 14 who found that the survival rates were at most 62%. The growing density of the larvae was higher in that study (84 larvae/L) compared with our study (10–20 larvae/L). These factors would certainly affect food availability and water quality and in turn the survival rate of the fish. The fish density (18 larvae/L) used in the study by Cavodeassi et al. 25 is comparable to our study. The survival rates of their fish fed with Artemia and the best dry feed (Practical diet) were highly similar (86% and 84%) to the optimized Kyowa protocol, further supporting the idea that 20 larvae/L is preferable to 84 larvae/L. Also, the high survival rates of the larvae at 14 dpf when they only received Kyowa feed suggest that the dry food can efficiently support the survival of the larvae during the most critical period of development from 6 to 15 dpf. 6 Further, the total number of eggs spawned by the wild-type fish raised using the 1 puff & trans protocol over a 5-week period and the associated 5 dpf survival rate were comparable to the siblings raised using the Paramecium protocol. Therefore, this protocol is also suitable for raising fish to produce a large number of high-quality eggs for transient genetic analyses and expression studies.
For the two types of mutant fish, smarca4a50 and inbred gassi, the 1 puff & trans protocol provided a similar survival rate in smarca4a50 but a lower rate in gassi (Table 5A, Supplemental Table S3A, B) compared with the Paramecium protocol. The inbred gassi has an adult albino phenotype that is hard to raise (see Materials and Methods section for the discussion on its pigmentation mutations). It has been noticed that for some mutants that have visual and motor impairments, the larvae may not be able to track the food well (unpublished observations). This is often true for albino fish. To raise these fish successfully, it is essential to present the food to their close proximity. Indeed, this is one of the major differences between live Paramecium and dry feed. The former is motile within the water and can be easily spotted by the fish, whereas dry feed tends to stay on the surface of the water and so the fish required more effort to locate and ingest. However, this is believed to be a desirable property for dry food because if it absorbs water rapidly and sinks to the bottom of the tank, it may substantially affect the water quality. 14 In this study, the Kyowa feed floated on the water surface exceptionally well compared with the Zeigler feeds. Therefore, the lower survival in gassi may be explained by a potential visual or motor impairment of the larvae, which compromised their ability to detect food mostly available on the water surface. This is supported by the observation that the majority of the casualties in the gassi fish occurred before 14 dpf when the larvae were fed exclusively with the dry food (data not shown). The other alternative hypothesis for the inferior survival rate is that the pigmentation mutations in the inbred gassi fish affected food absorption, feeding, or metabolism. Even though this hypothesis is less likely given that the mutations are supposed to affect pigmentation only, our experimental design could not rule out this possibility. Nonetheless, the gassi larvae raised using the 1 puff & trans protocol still had a comparable growth rate, sex ratio (Table 5B), breeding success rate (Table 5C and Supplemental Table S3C), fecundity, and quality of the resultant eggs spawned (Fig. 3, Table 5D and Supplemental Table S3D) to the Paramecium protocol. These data suggest that the quality of the mutant fish successfully raised using the dry food was equally good compared with those raised using Paramecium.
Although many studies only investigate the early embryogenesis of zebrafish until 5 dpf, before the feeding starts, there are some studies that require the investigation of metabolism and gut development at a later stage. As transgenic fluorescent reporter lines are commonly used in zebrafish research, the usage of dry foods that fluoresce under fluorescent illumination may impose a problem in these later-stage studies. In this study, the Paramecium feed (Paramecium + Zeigler food) had the strongest fluorescence in vivo (Fig. 4). Even though the Kyowa feed showed some fluorescence by itself (data not shown), the in vivo fluorescence intensity was not significantly different from those unfed larvae. Interestingly, the larvae fed with only Paramecium had less fluorescence than even the ones unfed. Although the cause of this difference is unclear, the data suggest that the Kyowa food may be suitable for studies that require the usage of fluorescence reporter fish. However, as the Kyowa food did fluoresce and the data for the fish fed by this food also showed a trend to have higher fluorescent intensities than the control (Fig. 4), one should use caution when using Kyowa food in studies that require fluorescent quantitation.
There are two potential drawbacks of using dry feed in general. First, one has to rely on the manufacturing company for quality control. Given the contamination issues in feeds for food-production animals, including bacteria 26 and prions, 27 there is always a risk for food-borne disease associated with the adoption of using commercially prepared feeds for research animals. Indeed, the raw ingredients of the Kyowa feed contain fish powder and krill (see Supplemental Material 2 for a pamphlet on Kyowa [available only in Japanese]). Even though zebrafish are not raised for consumption, any unwanted contamination in the feed that can compromise fish health would affect the progress and quality of scientific research. Although this issue was not investigated in our study, there have not been any major health issues in our fish facility. Second, a consistent and well-defined formulation of food is often crucial for many toxicological and physiological researches. Unfortunately, the formulation of commercial feed is usually proprietary, which makes it impossible to elucidate any confounding effects on a study if the ingredients vary from batch to batch. The drawback is partially alleviated by the fact that zebrafish can be successfully maintained and spawned in a wide variety of diets and feeding regimes, which can be suboptimal. 12 In this study, all quality measures tested indicated that the 1 puff & trans protocol was comparable to live feed, indicating the nutritional value of the Kyowa feed was optimal.
In conclusion, the results indicate that the quality of the fish raised using the 1 puff Kyowa with transition protocol with up to 20 larvae/L (Table 2) is at least as good as the Paramecium protocol in most scenarios and can substantially reduce the routine maintenance workload. As the amount of Kyowa food delivered was measured, it is also possible to administer the food by automatic feeder to further reduce the workload. When the fish are particularly hard to raise, it may be advisable to supplement dry food with live food to increase the survival rate.
Footnotes
Acknowledgments
The authors thank Paul Collodi for providing the inbred gassi mutant line; Jennifer Crodian and Patrick Gibbs for their background information and technical assistance on maintaining gassi line; Deborah Biesemeier for her assistance in general fish maintenance in the joint zebrafish facility; Yuqing Zhang for her assistance in fish length measurement; Cindy Lui and Yang Zhao from the statistical consultation service in the Department of Statistics at Purdue University for their help in data analysis; Jessica Miller, Steven Zimmerman, members from Donna Fekete, Yuk Fai Leung, and Daniel Szeto's laboratories for helpful discussions.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.