
Abstract
Abstract
Aggression is a key component of the behavioral repertoire of animals that impacts on their Darwinian fitness. The available genetic tools in zebrafish make this species a promising vertebrate neurogenetic model for the study of neural circuits underlying aggressive behavior. For this purpose, a detailed characterization of the aggressive behavior and its behavioral consequences is first needed. In this article we establish a simple protocol that reliably elicits the expression of fighting behavior in zebrafish dyads and characterized it. The agonistic behavior expressed during dyadic fighting behavior has a temporal structure, indicating the existence of an underlying architecture prone to genetic manipulation. Social interactions have consequences for subsequent behavior with a potential fitness impact, which stresses the validity of this species for the study of aggression. These effects of experience seem to be mediated by different mechanisms in winners and losers. Winners increase the probability of winning subsquent fights without changing their fighting behavior, suggesting the existence of social status cues. On the other hand, losers decrease the probability of winning subsequent fights by decreasing their motivation to escalate fights. Together, these results are a first step to the development of a quantitative framework for the study of aggressive behavior in zebrafish.
Introduction
Recent studies in zebrafish combining molecular genetics with behavioral analyses have allowed the identification of genes involved in neuronal circuits underlying specific behaviors and mechanisms involved in neuropathogenesis.3,7 Zebrafish models of brain function and disease have started to be developed, including insomnia and sleep disturbances,8–10 movement disorders, 11 autism, 12 neurodegenerative diseases, 13 cognitive impairment during aging, 14 and nicotine and alcohol addiction.2,15–17 Complex behaviors that are goal-directed (e.g., escape from predators) or emotion-related (e.g., aggression, anxiety, and fear) have also started to be characterized in adult zebrafish, and the first results suggest conserved regulatory mechanisms with mammals, 18 including shared modulatory neurotransmitter systems13,19 and homologous brain areas. 20
Aggression serves various adaptive functions, such as the establishment of dominance relationships and hierarchies and the competition for key resources such as food, shelter, or mates and territories, 21 and therefore plays a major role in Darwinian fitness. Despite its biological relevance and the large body of literature dedicated to the study of aggression, there is not yet an established vertebrate neurogenetic model organism for its study that would allow the use of powerful genetic tools for the dissection of the neural circuits involved, and for the understanding of how they are activated by social cues and regulated by humoral factors (but see ref. 22 for the development of a neurogenetic model of aggression in fruit flies and refs.23,24 for previous work on knock-out mice for candidate genes in the serotonergic system). For reasons mentioned above zebrafish can play such a role. For that purpose one needs first to establish that aggressive behavior is present and has a temporal structure (i.e., its sequence is not random, suggesting an underlying regulatory mechanism prone to genetic dissection), and that it has consequences for the animals (i.e., subsequent behavior is shaped by previous interactions). Since zebrafish is a gregarious species that exhibits shoaling behavior in captivity, only recently its aggressive behavior has attracted the interest of researchers. Several studies have now demonstrated that both male and female zebrafish exhibit aggressive behavior (see refs.19,25 for recent reviews), that stereotyped behavioral patterns can be observed and described in detail during agonistic interactions (e.g., refs.15,26), that territoriality and dominance hierarchies can be present,27–29 and that neuropeptides (i.e., AVT) and steroids are associated with aggressive behavior.30–32
The main goals of this article are (i) to establish a behavioral paradigm under which male zebrafish would consistently express fighting behavior; (ii) to characterize the structure (i.e., temporal pattern) of fighting behavior in male dyads; and (iii) to study the effects of social experience (i.e., winning/losing effects) on subsequent fights. Together, these goals will contribute to the establishment of male–male fights in zebrafish as a standardized behavioral paradigm for the study of the genetics of aggression.
Materials and Methods
Subjects and maintenance
The individuals used in this experiment belong to an F2 generation population bred at Instituto Gulbenkian Ciência, which derived from wild-type (AB) zebrafish (Danio rerio) acquired from Zebrafish International Resource Center. Before the experiment, animals were kept in 8.0 L tanks (30 × 22 × 15 cm) with a sex ratio of two females per each male. Fish were kept at 26°C ± 2°C on a 14D:10L and fed twice daily with freshly hatched brine shrimp, in the morning, and with commercial food flakes, in the afternoon. In this study, the average male size was 28.1 ± 1.7 mm (standard length, SL).
Experimental procedure
One of the main aims of this study was to establish a reliable behavioral paradigm to study aggression in zebrafish. Although mirror image stimulation (MIS) has been widely used as an aggression test for zebrafish, it does not elicit the full agonistic repertoire and the brain activation pattern and hormonal response associated with MIS differ significantly from those triggered by a fight with a live opponent.33–35 Therefore, we focused on dyadic fights between size-matched males. Since we wanted to create the simplest situation possible in which zebrafish would express their agonistic repertoire, in pilot studies we tested if male dyads would fight in the absence of a limited resource (e.g., shelter, mate, and food) after a period of social isolation. A previous study 30 has already used successfully an isolation-induced aggression paradigm with zebrafish, using a social isolation period of 5 days. In our pilot studies we have established that 24 h of social isolation was enough to promote the consistent expression of aggressive behavior in male dyads and this is the behavioral paradigm that we have used in this study.
Twenty-two male dyads were formed with individuals matched for standard length (size difference < 1 mm, which is on average 3.6% of body size) within each dyad. Subjects were individually recognized by fin clips on the extremities of the caudal, dorsal, or anal fins, which were conspicuously distinguishable. Each pair was placed in a 700 ml polycarbonate tank (18 × 10 × 9 cm) visually, but not chemically, isolated by a removable opaque PVC partition and allowed to acclimate overnight. After 1 day in isolation, the opaque divider was removed and the fish were left to interact for 30 min, a duration that exceeded the necessary time to determine a clear winner of the contest. After each interaction, the fish were separated again by placing back the opaque partition. Behavioral interactions were videotaped and were subsequently observed in detail. After this first interaction, both fish were separated into two new tanks and paired up with two other fish, matched for size, but with no prior fighting experience (i.e., naïve individuals), separated by an opaque partition. After a 1 h acclimation period, the opaque partitions were removed and the experienced and naïve fish could interact for 30 min (Fig. 1). These second interactions were also videotaped for subsequent behavioral analysis.
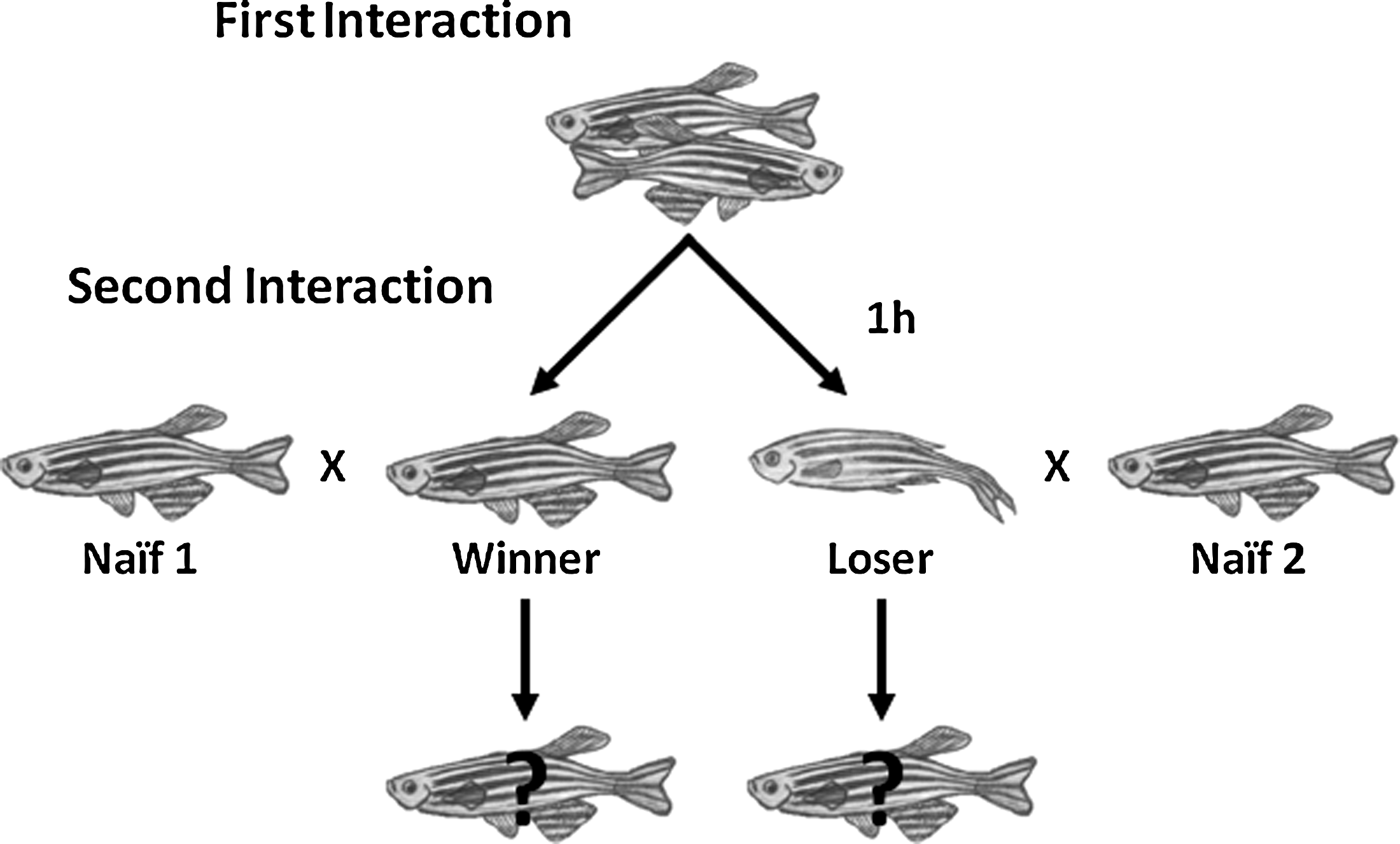
Diagram illustrating the self-selection protocol used to test experience effects in the agonistic behavior of male zebrafish. During the first interaction, two naïve fish matched for size and previously isolated for 24 h are paired up. One hour after the resolution of the first interaction, during which each fish is kept in social isolation, the winner/loser of the first interaction is again paired up against a naïve size-matched opponent.
Behavioral observations
Video recordings (Sony KDL × 200) were analyzed using the software Observer XT (Noldus). An experienced observer analyzed the behavioral interactions and identified all agonistic behaviors according to the ethogram presented in Table 1 and determined, based on the asymmetry of aggressive behavior (i.e., who attacks and who is submissive in the later part of the interaction) the winner and loser of each interaction. The ethogram used in this study was build based on ca. 20 h of ad libitum observations of male zebrafish fights that have been previously videotaped. Stereotyped behavioral patterns that were consistently present in the interactions were described in detail.
Frequency, latency and duration of agonistic behavior were registered using a behavior sampling with continuous recording (sensu 36 ). After detailed scrutiny of behavioral patterns exhibited during interactions, a switching point in the interaction was identified, where symmetric aggressive behaviors (e.g., displaying, circling) gave place to asymmetric ones (e.g., attack/flee, chase). This was considered the point at which the resolution of the fight took place, and since after the establishment of an asymmetry we have never observed a status reversal (i.e., the attacker becoming the attacked), one of the fish clearly could be considered the winner of the interaction. Based on this fight resolution point we defined two phases in the fight: (1) a preresolution phase and (2) a postresolution phase. In the former phase, all behaviors from both subjects were quantified until the fight resolution point; in the latter phase, due to the consistency of the behavioral patterns exhibited by the fish (i.e., chase/flee, bite), only the last 5 min of the 30 min sampling were analyzed.
Eleven dyads for which the identity of the fish (i.e., the fin clips) were recognizable in the video-images of the three interactions (i.e., first interaction between emerging winner and loser, the second interaction between the previous winner and the naïve male, and the second interaction between the previous loser and the naïve male) were used for detailed analysis behavioral analyses. The observer had to identify each animal in frames where clips were clearly observable and then track each animal individually along the video recording of each behavioral trial. When there were doubts on the identity of the fish during the video-analysis the observer had to reverse the video until an unmistakable image was found and then progress again with the analysis. When it was not possible to solve the identity of the subjects in a given part of the trial, and therefore it was not possible to track individually the two opponents along the whole session, these trials were dropped from the analysis.
For the other dyads in which the identity of the fish could not always be followed in the videos, but could be assessed at the beginning and at the end, only the outcome of the fight (i.e., identity of the winner and of the loser) and the identity of the initiator of the fight were collected.
Statistical analysis
To characterize the structure of the behavioral sequences present in zebrafish fights, a transition matrix was build indicating the frequency with which each behavioral pattern followed and was followed by each other behavior of the zebrafish agonistic repertoire within each individual. The diagonal was kept at zero since we considered that each behavior pattern could not be followed by itself. This behavior sequence matrix was analyzed using a first-order Markov chain analysis to identify nonrandom transitions between behavioral elements (i.e., nonrandom temporal associations between behavioral patterns 37 ). Only data from the first interaction were used for this sequential analysis. These analyses were performed using a collection of freeware programming functions developed by Robert Huber (Bowling Green State University) for the analysis of behavioral data (Java Grinders Library v.4.0 “Essential Equipment for Ethology,” available on the Internet at http://caspar.bgsu.edu/∼software/Java/).
To study the effect of the phase of the fight (pre- vs. postresolution) and the status of the fish (putative winner vs. putative loser) on the expression of different aggressive behaviors, the frequency and duration (when appropriate) of each behavior pattern in the first interactions was analyzed using a repeated measures ANOVA. To study the impact of previous experience on subsequent behavior the frequency and duration (when appropriate) of each behavior pattern, the latency for the first interaction and fight duration were compared between the first interaction (i.e., putative winner vs. putative loser) and each of the two second interactions (i.e., prior winner vs. naïve and prior loser vs. naïve), using an ANOVA model with the phase of the fight (preresolution vs. postresolution) as a repeated factor and status (winner vs. loser [WL]) as an independent variable, followed by planned comparisons using contrast analysis. Apart from these behavioral variables we have also computed a composite measure of fight escalation as follows:
(1) Escalation index = overt aggression/overt + ritualized aggression = (bite + chase + strike)/(bite + chase + strike) + (display + circle)
All descriptive and inference statistics were run on the statistical software package STATISTICA v. 8.0 (StatSoft 2007). Differences between proportions of second fights won versus lost by previous winners versus losers of first fights were tested by computing the qui-squared value for the resulting 2 × 2 contingency table. All tests were two-tailed and used a significance value of p < 0.05.
Results
Agonistic behavior in zebrafish
The behavioral patterns observed during male–male fights in zebrafish are described in Table 1. Dyadic fights among male zebrafish have two distinct phases. The first phase consists mainly of mutual assessment behaviors, with fish assessing each other by exhibiting display, circle, and bite behaviors (Table 1) to determine the other fish's relative fighting ability. This phase starts with the first interaction of the behavioral trial (latency for first interaction =70.6 ± 164.9 ms) and lasts on average for 379.2 ± 331.0 ms, until the first chase/flee is observed which marks the point of the resolution of the fight. In the second phase, that occurs after the fight's resolution, all agonistic behaviors are initiated by the winner (e.g., bite, chase, and strike), whereas the loser tries to flee and displays submission and freezing postures. At the end of fights losers usually stay near the bottom or top of the tank adopting a submissive posture. During the 30 min of the behavioral trial, male zebrafish displayed agonistic behavioral patterns at a rate of 1.19 behavior/s. Bite was the most frequent behavior, representing roughly 65% of all behaviors exhibited (N = 5769 behavioral acts) by the fish in the first phase. In the postresolution phase only the winner of the interaction exhibited Bite behavior, but it represented approximately 50% of its behavioral output (N = 2842 behavioral acts). All other behaviors, though less frequent in number, represent part of a complex and highly structured behavioral sequence, which characterizes zebrafish agonistic behavior. To better understand and describe these sequences, a behavior transition matrix was analyzed and nonrandom transitions between behavioral elements were identified. This analysis reveals a temporal structure in male zebrafish fights (i.e., behavioral sequences are nonrandom; behavioral sequence matrix X 2 = 2242.8, p << 0.0001) with assessment behaviors (i.e., display and circling) significantly associated with each other and with bite that is also then significantly associated with chase and strike that correspond to the asymmetric phase of the fight (Fig. 2). A set of behaviors associated with losing the fight (i.e., freeze, flee, and retreat) also appear significantly associated among themselves (Fig. 2).
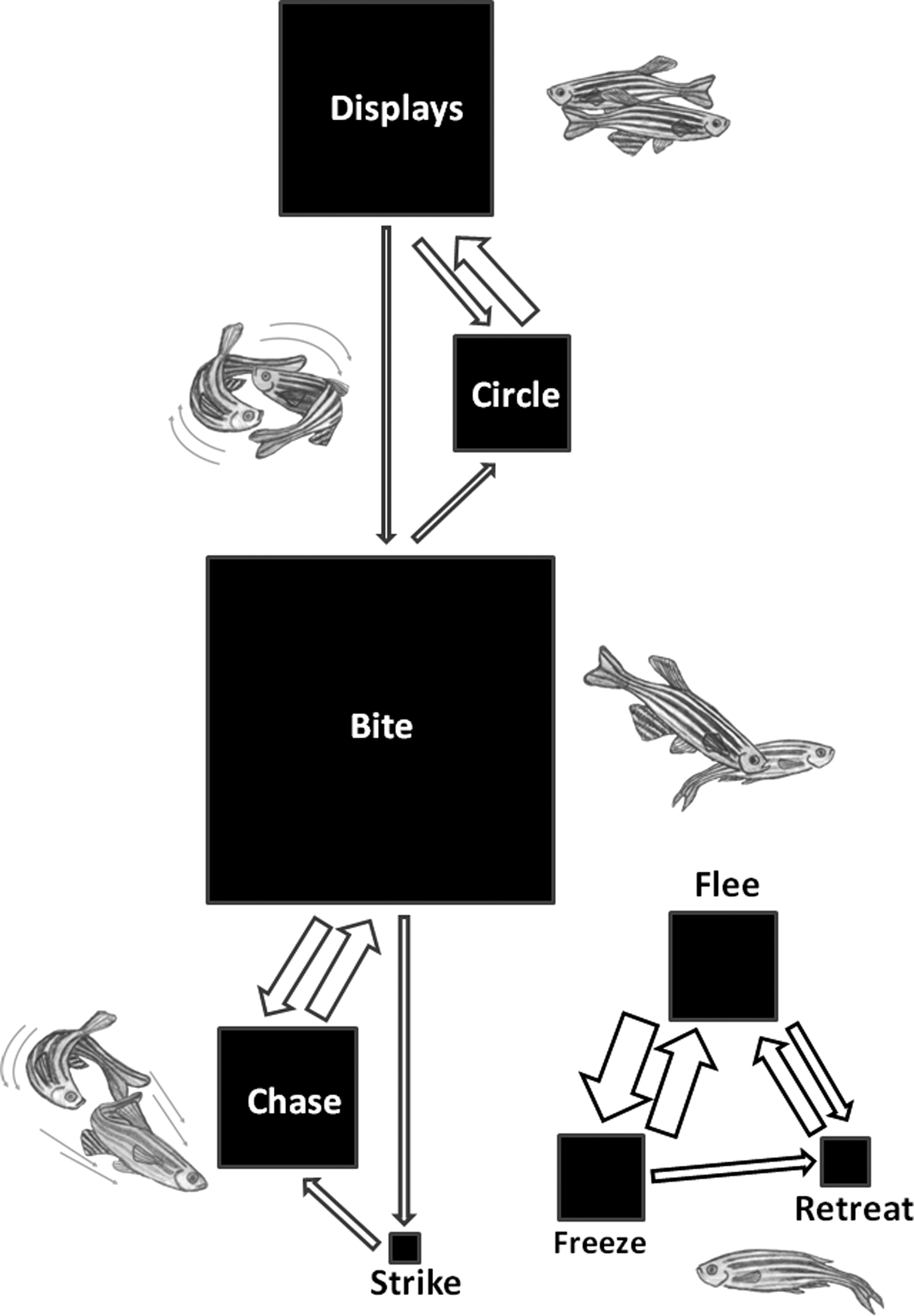
Temporal structure of fighting behavior in male zebrafish dyads using a first-order Markov chain analysis. The size of each box is proportional to the relative frequency of occurrence of each behavioral pattern. Transitions between behaviors whose frequency is significantly higher than chance levels are depicted as arrows, and their size indicates the degree of significance.
Temporal dynamics of the fights and early predictors of success
To further study the effect of the phase of the fight (pre- vs. postresolution) and the status of the fish (putative winner vs. putative loser) on the expression of different aggressive behaviors, the frequency and duration (when appropriate) of each behavior pattern and the composed measure escalation index were analyzed using a repeated measures ANOVA (Table 2).
Frequency, duration, and escalation index were analyzed using a repeated measures ANOVA, with the phase of the fight as a repeated measure (R1; pre- vs. postresolution phase) and status (winning vs. losing) as an independent variable. Apart from the main effects and the interaction between the two factors, results of contrast analysis for the winner loser comparison in the pre- and postresolution phases are also given. All significant results (p < 0.05) are underlined. Note:
As expected behaviors associated with mutual assessment were significantly more frequent and had a longer duration in the preresolution phase of the fight (Table 2) and a nonsignificant trend for an increase in the frequency of Chase, Freeze, Flee, and Retreat was also observed from the pre- to the postresolution phase (Table 2). The phase of the fight had no main effect on the escalation index. Social status has a main effect on the expression of bite and flee, with the former being more frequent in winners and the latter in losers, and on escalation index that is higher in winners (Table 2). We have also performed planned comparisons to test if differences in the expression of aggressive behavior were already present in the first phase of the fight between individuals that subsequently became winners versus losers. None of these planned comparisons was significant for the preresolution phase, indicating that neither the frequency of any of the behaviors measured in the early stage of the fight nor fighting escalation at this phase are good predictors of fight outcome (Table 2). On the other hand, similar planned comparisons comparing the frequency of each behavior and of fight escalation between winners and losers in the postresolution phase of the fight revealed that winners express significantly more bites and chases and a higher escalation index, and losers more flee in the advanced stage of the fight (Table 2). Interestingly, the escalation index significantly increased in winners and decreased in losers from the pre- to the postresolution phase (losers: F1,20: 7.19, p < 0.05; winners: F1,20= 6.60, p < 0.05).
Since fish were matched for standard length within each dyad, size was also not a predictor of the fight outcome (SL of winners = 2.804 ± 0.035 mm; SL of losers = 2.800 ± 0.038 mm, t (1, 22) = −0.25, p = 0.80). Being the first to engage in the interaction, which could be seen as a proxy of aggressive motivation, was also similar between individuals that became winners and individuals that became losers (six winners vs. seven losers, qui-square = 0.077, P = 0.78).
Experience effects
Most of the winners of the first interaction also won the second interaction against a naïve individual (85.71%), whereas only a very small percentage of fish that lost the first interaction won the second fight (4.55%), suggesting the presence of both winner and loser effects in zebrafish (X 2 = 28.7, p < 0.0001; Fig. 3).
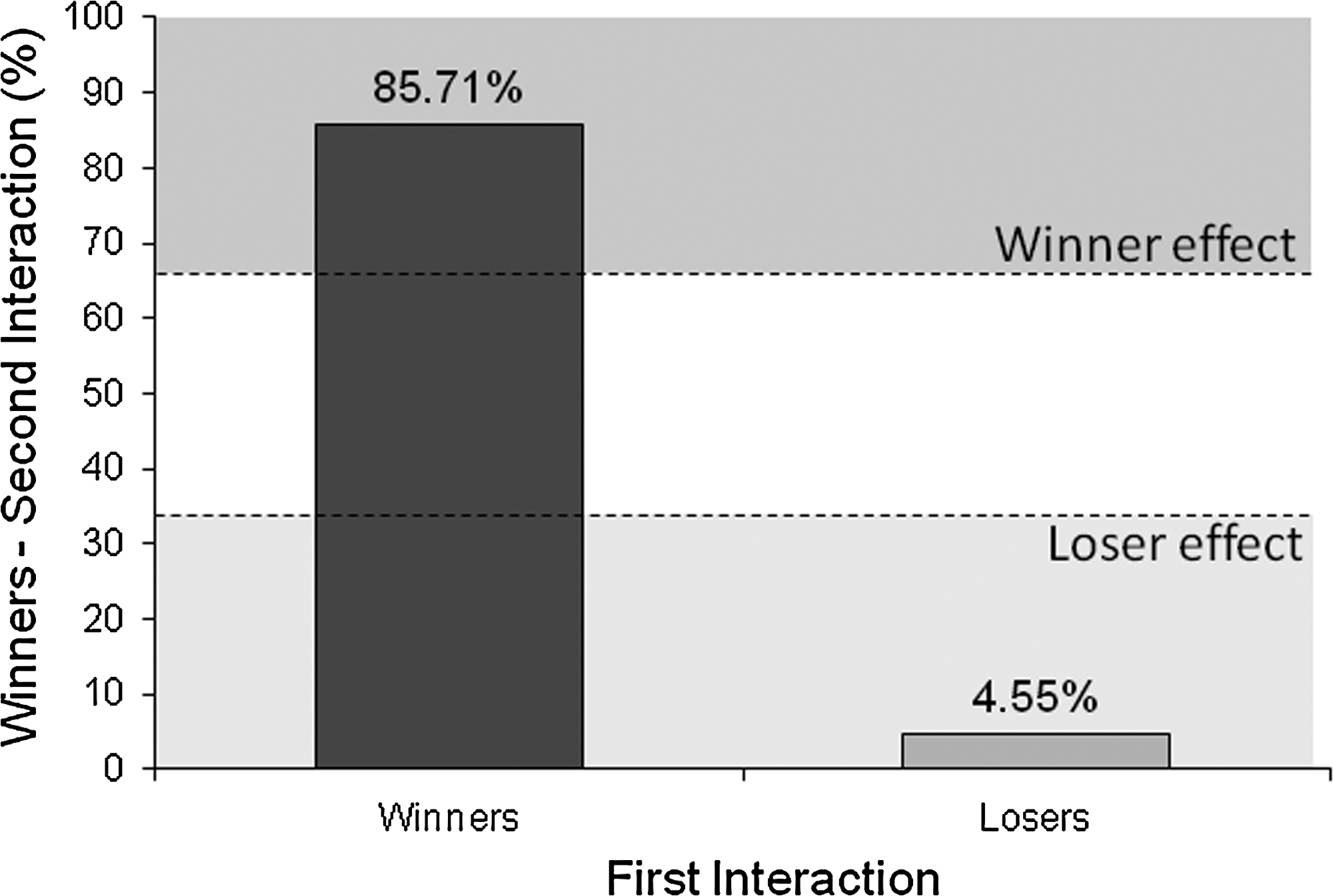
Experience effects in zebrafish as indicated by the percentage of victories/defeats of previous winners and losers in the second fights. Dashed lines indicate the cut-off values for the detection of winner/loser effects calculated considering the variation in intrinsic fighting ability according to ref. 44
To investigate the behavioral mechanisms that may account for these winner/loser effects, we investigated the variation in motivation and persistence induced by the previous fight, by comparing the first fights (i.e. WL) and second fights (i.e. winner vs. naïve [WN] and loser vs. naïve [LN]). As a proxy of fighting motivation of the dyad we compared the latency for the first interaction between WL and WN and between WL and LN and the identity of the initiator of the fight. As a proxy of fighting persistence in the dyads, we compared the latency for the resolution of the fight between WL and WN and between WL and LN. Although there is a trend for second fights to start sooner than the first fight, no significant differences were found for the latency for the first interaction for the planned comparisons described above (N = 11; WL/WN: t = 0.74, p = 0.47; WL/LN: t = 1.45, p = 0.17; WN/LN: t = −1.62, p = 0.13; Fig. 4a). There was no effect of prior experience on taking the initiative to start the second fight, since 3/11 previous winners and 3/11 previous losers initiated the second fights. The time needed to reach the resolution of the fight tends to be shorter in second fights (Fig. 4b), but the difference is only significant for the WL versus LN comparison (t = 2.55; p < 0.05). Moreover, the time needed for the resolution of the fights in the two second interactions is significantly shorter for those involving the previous loser than for those involving the previous winner (i.e., WN/LN: t = 2.86; p < 0.05). The escalation index does not change significantly in winners between the first and the second fight (ANOVA repeated measures, contrast effect: F1,20= 0.08, p = 0.77; Fig. 5), but it decreases significantly in losers (ANOVA repeated measures, contrast effect: F1,20= 5.84, p < 0.05; Fig. 5). As a consequence, the escalation index of previous winners and previous losers is significantly different in subsequent fights against naïve opponents (ANOVA repeated measures, contrast effect: F1,20= 9.49, p < 0.01).
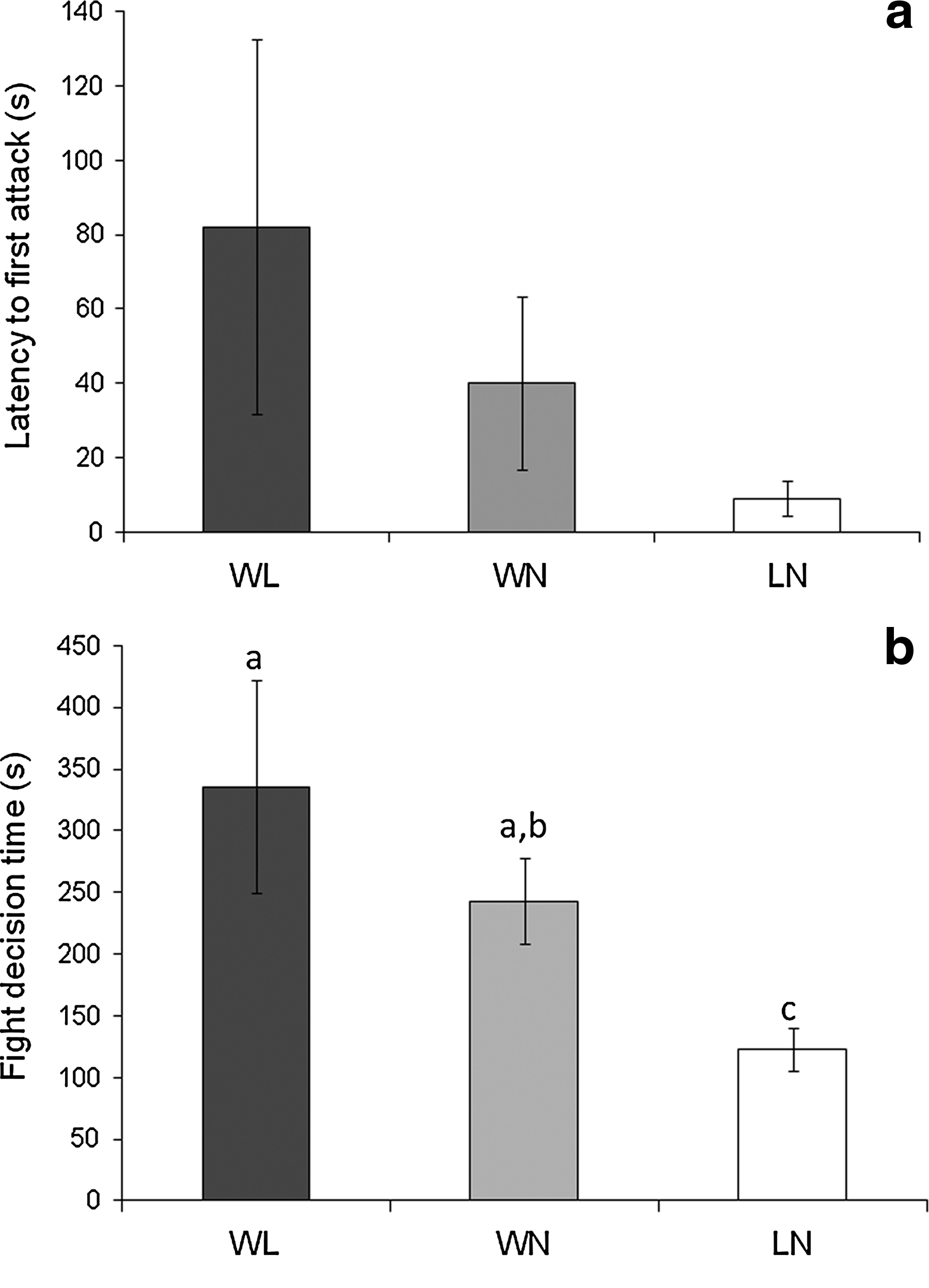
Comparison of latency to first attack
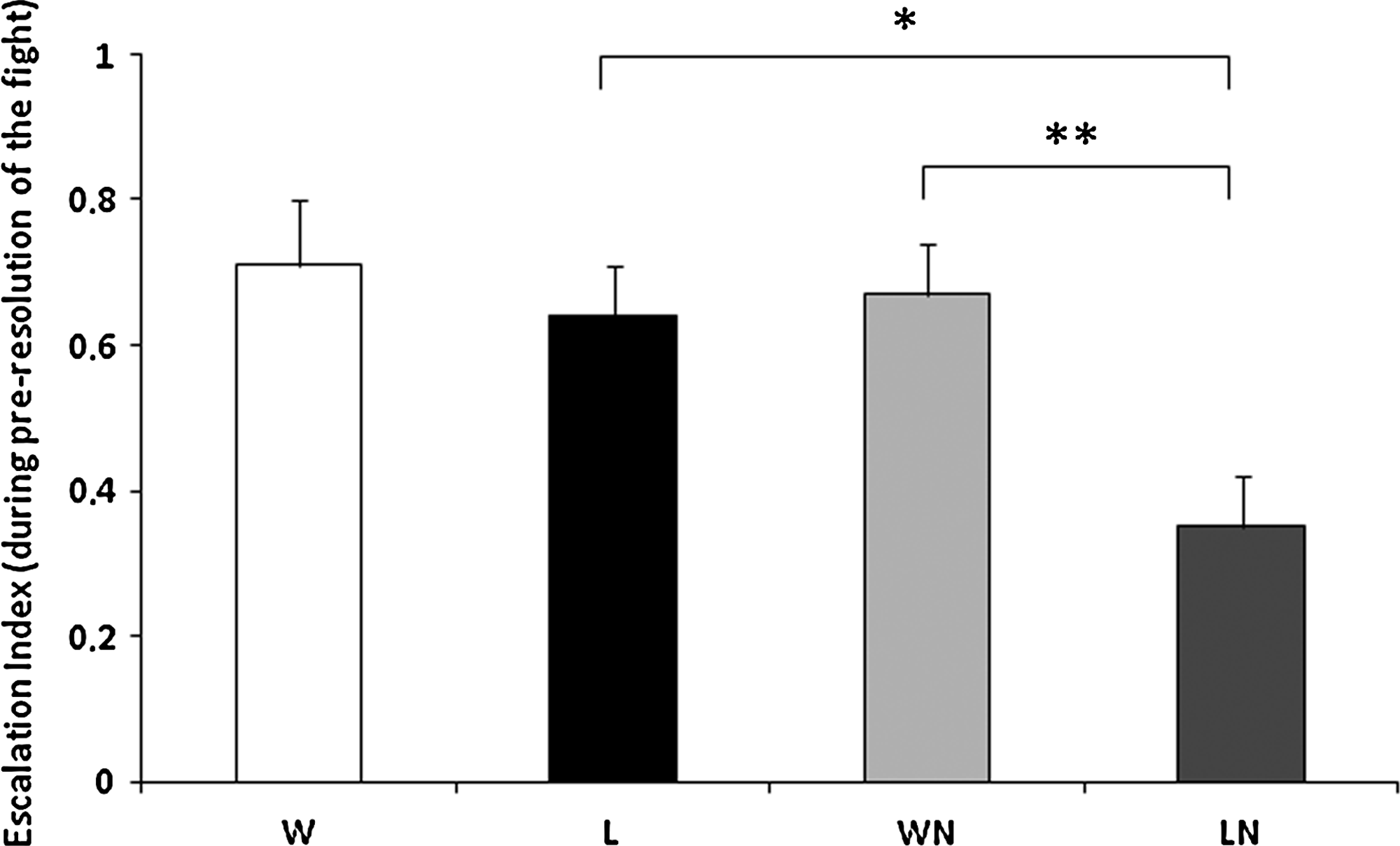
Comparison of escalation behavior in the preresolution phase between winners (W, white) and losers (L, black) of the first fight, and between the focal males in the second interaction (WN, prior-winner vs. naïve, light gray; LN, prior-loser vs. naïve, dark gray). * indicates p < 0.05; ** indicates p <0.01.
Discussion
In this article we have described a simple behavioral paradigm under which male zebrafish consistently expressed fighting behavior. Dyads of two males that have been previously isolated for 24 h consistently expressed fighting behavior when exposed to each other, even in the absence of a limited resource to promote competition, such as food, shelter, or a potential mate. In a previous study,
30
5 days of social isolation have been used to promote aggression in zebrafish. Our study indicates that 24 h of social isolation is enough to promote the expression of aggressive behavior in zebrafish. Social isolation has been reported to increase aggression in different species, including fish (e.g., ref.
38
). Different mechanisms may explain the effect of social isolation on aggressiveness, including an increased sensitivity to external stimuli that may act as releasers of aggressive behavior, or forgetting prior social experiences that are the basis for dominance hierarchies that regulate social interactions in social networks. Whatever the mechanisms involved, for the purpose of this study, the key result is that a short period of social isolation consistently promoted the expression of aggressive behavior in dyads of male zebrafish. So far the study of aggression in zebrafish has mainly used either standardized mirror image stimulation tests (e.g., refs.15,39–41) or groups of three or more individuals (e.g., refs.27–29,42–44). Although the mirror test can be seen as a standardized test that elicits heightened aggressive responses,45,46 it has recently been shown that it triggers different hormonal, and brain activation patterns from those elicited by a real opponent33–35
(RF Oliveira et al., unpublished data for zebrafish). Therefore, the use of mirror image stimulation as a behavioral paradigm to study aggression should be taken with caution, and the use of real opponent fights is advisable, especially when studying the proximate mechanism of aggressive behavior. It should also be mentioned here two major technical and analytical challenges of analyzing zebrafish aggression that we have faced:
Due to the high speed with which some of the behavioral patterns are performed by the fish, a frame-by-frame analysis was recurrently needed; this is very time-consuming and in some cases a higher time resolution would have been helpful. In this respect a high-speed camera (with image acquisition rates staring at ca. 200 frames per second) would be a major improvement when compared to regular video cameras (with acquisition rates of 25–30 frames per second depending on which video signal standard is being used, PAL or NTSC, respectively). The small size of zebrafish is a limitation to the identification of particular individuals during behavioral trials either in real time or in video-recordings. We have used fin clips to individually tag subjects. Despite being very efficient to identify individuals in stock tanks this method proved inefficient to identify individuals in the video-recordings, as illustrated by the fact that we had to drop the video-analysis of almost 50% of our behavioral trials due to the lack of a clear identification of each individual along the whole session. We have recently replaced with good results the use of fin clips by fluorescent elastomer tags (Northwest Marine Technology Inc.) that are implanted beneath transparent or translucent tissue and remain externally visible. Although this is a promising solution for individually tagging zebrafish in behavioral assays we still have to confirm if the different colors are having an effect on the behavior of the fish.
The qualitative aspects of the behavioral patterns observed in our experiment are consistent with other ethograms previously published for this species (e.g., refs.25,28–30). The sequence analysis based on transition matrices of behavioral patterns expressed by each contestant allowed to identify a complex and highly structured aggressive behavior in zebrafish, indicating that the expression of the different behavioral patterns that make up the agonistic repertoire is not random and that there are decision rules underlying their expression. This is a particularly relevant finding since it makes aggressive behavior in this species suitable for quantitative analysis and allows for the study of the impact of selected mutations or other genetically or pharmacologically induced changes in behavior.
Zebrafish fights lasted for ca. 5 min until a clear asymmetry was established and a clear winner and a clear loser could be established. Before fight resolution contestants mainly expressed mutual assessment behaviors such as displays and circling, and biting. After the resolution point, winners mainly expressed chase and bites and subordinates flee. Biting is present in both phases (i.e., before and after resolution but its frequency increases in the postresolution phase where physical aggression becomes more frequent. Contrary to other studies (e.g., ref. 47 ), the initiative to start the interaction was not a good predictor of fight outcome, and there was no behavior pattern whose expression in the preresolution phase was an indicator of who would win the contest. Therefore, we have failed to identify a key fight parameter used by fish to decide when to give up and retreat from fighting.
Data presented here on sequential fights show that a recent victory increases and a recent defeat reduces the probability of winning a subsequent fight, suggesting the occurrence of winner and loser effects in male zebrafish. This is an important result since it establishes that fight outcome has behavioral consequences that may impact in the individuals Darwinian fitness.
Since we have used a self-selection protocol and did not impose winning and losing experiences to our focal fish, it can be argued that the winner/loser effect observed is due to uncontrolled sources of interindividual variation in intrinsic fighting ability. To control for this possibility, we have used size-matched males, and a posteriori we also controlled for fighting motivation by registering, which male took the initiative to start the fight. Moreover, we have used a null hypothesis against which to test the effects of prior experience that is not equiprobability of winning/losing the second encounter, but having prior winners/losers winning/losing at least two-thirds of subsequent interactions against a size-matched naïve opponent, which is the probability estimated by ref. 48 of a random individual in a population to have higher/lower intrinsic fighting ability than neutral opponents.
Two behavioral mechanisms have been advanced to explain the effects of prior experience on future fighting success: (1) changes in self-assessment of fighting ability (i.e., resource holding power, sensu 49 ) induced by the fighting outcome (i.e., perceived increase in winners and perceived decrease in losers of own fighting ability), and (2) social cues that signal the winner/loser status to conspecifics allowing them to respond differentially to winners versus losers of a previous fight.50–52 These two mechanisms are not mutually exclusive and evidence for both is present in the literature. In support of the former, after a recent win or loss individuals change their contest behavior accordingly in standard aggression tests and increase the probability of initiating a new fight (see ref. 51 for a review). In support of the latter it is known that fish collect information from observed interactions between third parties and that they respond differentially to individuals that they have observed winning/losing a previous interaction. 53 Moreover, a number of social cues are known to signal social status. For example, both in crayfish and in cichlid fish social dominance is signaled through odorants present in the urine.54,55
In our study the likelihood of starting a second fight was not affected by the outcome of the first fight (i.e., same numbers of previous winners vs. losers took the initiative to start the second fight). On the other hand, the escalation index does not change significantly in winners between the first and the second fight, but it decreases in losers so that in second fights previous winners express more escalated fighting behavior than previous losers. Similarly, the fight resolution time decreased in the second fights and was significantly shorter in the LN fights than in WN fights. Together, these results suggest that the effects of previous experience might be different in winners and losers: while escalation decreases in losers in subsequent fights, suggesting an experience-driven change in the self-assessment of their own fighting ability, the behavior of winners does not seem to change significantly in the subsequent fight, and therefore experience effects in winners may be relying on social cues that signal a recent winning that naïve opponents in the second fights are responding to. These cues can be behavioral, pheromonal, or other. In another teleost fish (i.e., tilapia, Oreochromis mossambicus) it has been recently shown that dominant individuals release more urine than subordinates during agonistic encounters and that the urine of dominants can be discriminated from that of subordinates at the levels of the olfactory organ with that of the dominants eliciting a higher olfactory response.55,56 Since in zebrafish olfactory communication is also well developed and used in social context (e.g., refs.57,58), it is possible that experience-induced changes in social status or motivation to engage in a contest are signaled through olfactory cues.
The fact that second interactions involving the loser of the first interaction have a short latency for the first interaction and a shorter resolution time suggests that the loser effects has a higher impact that the winner effect. This is in accordance with the relative magnitude of winner and loser effects reported in the literature (for review see ref. 50 ). These results are also consistent with a heightened decision to retreat in previous losers, an effect that is consistent with previous work in other species.59–61 An increased likelihood to give up appears to be the real explanation for the behavioral changes; however, this is not spelled out specifically (see page 15, line 334).
In summary, in dyadic fights male zebrafish express highly structured behavior and the outcome of these fights has an impact on their subsequent behavior. Given the available genetic and genomic tools for this species, these results support the use of zebrafish as a neurogenetic model for the study of the neural and hormonal mechanisms of aggressive behavior in a vertebrate model.
Footnotes
Acknowledgments
The authors thank Magda Teles and Paulo Raimundo for helping with the care of the fish and with behavioral analysis, Robert Huber for assistance with the analysis of transition matrix, and two anonymous referees for constructive comments on a previous version of the article. This study was funded by a research grant from Fundação para a Ciência e a Tecnologia (FCT), Portugal (grant # PTDC/PSI/71811/2006), and by the Pluriannual FCT Programme (UI&D 331/2001). JMS is supported by an FCT Ph.D. Fellowship (SFRH/BD/40976/2007).
Disclosure Statement
No competing financial interests exist.