
Abstract
Abstract
Although zebrafish is used as a major model species for understanding a number of biological functions and mechanisms involved, there is practically no information on the nutritional requirements of this cyprinid. The rearing practices, especially with regard to feeds and feeding, also vary considerably. To obtain base line information on the nutrient requirements and to develop a standard diet, we initiated the present work of feeding zebrafish larvae with a formulated feed right from first feeding onward. Growth of zebrafish fed the compound feed was very good, reaching a total length of 23 ± 4 mm in 9 weeks with a survival rate of 89% ± 4%. We present the first ever published data on whole-body composition in terms of essential amino acids (ideal protein profile), total, neutral and polar fatty acid profiles, minerals, and trace elements. Results obtained here show clearly that zebrafish can be reared with formulated feed right from mouth opening without resorting to any live prey.
Introduction
Given the use of zebrafish as a model in developmental biology, especially during early ontogenesis, it is rather surprising to note that the larvae of zebrafish are reared and weaned using diverse protocols, going from paramecia to other live preys to weaning with feeds of undefined nutritional composition (http://zfin.org/zf_info/zfbook/zfbk.html; Fig. 1). Despite the interest for zebrafish as an interesting model organism in fish nutrition research, the potential has not yet been realized. 3 This is because our knowledge on the nutrient/energy requirements of zebrafish is largely inexistent. 4 The few studies undertaken so far are limited to those of a summary report on n-3/n-6 polyunsaturated fatty acids (PUFA), 5 possible supply of riboflavin through special delivery vectors, 6 the dietary essentiality for retinoids, 7 and on the need for water-borne boron for normal embryogenesis. 8 Other studies have looked into the possible effects of dietary nutrients on fatty acid (FA),9,10 carbohydrate, 11 or zinc 12 utilization and metabolism in juvenile fish. To undertake nutritional studies and to precisely define the nutrient requirements, and consequently work on the effects of nutrients on diverse metabolic functions, it is essential to develop formulated feeds of known composition that are well accepted by the target organism and in which the nutrient levels can be modulated. This can also eventually lead to a steady line of larval diets for zebrafish that would very much help basic studies undertaken with this model animal.

Rough scheme for the production of zebrafish (based on Westerfield 2 ). Age is given in days postfertilization at 28.5°C.
Onal and Langdon 13 observed that with 12-day-old zebrafish previously reared on Artemia or a formulated feed, between 20% and 40% of Artemia nauplii could be replaced by specially developed microparticle feeds (cross-linked protein-walled capsules or gelatin alginate beads) with equal growth and survival over the 8 days of the study, although the growth rates were small. More recently, Siccardi et al. 14 compared several commercial and experimental feeds with 28-day-old zebrafish previously fed over 9 weeks and found that juvenile zebrafish could be reared with success using such feeds.
Most of the morphological and functional changes occur during the first few days or weeks of exogenous feeding in larvae and the capacity for absorption of nutrients varies depending on the life history and the pattern of development of the digestive tract.15–18 Given the importance of zebrafish as a model for changes occurring during early ontogenesis, it is essential to exert close control on the qualitative and quantitative supply of nutrients during these early stages. As regard early weaning of zebrafish larvae with formulated feeds of known composition, despite several efforts, data available are also not conclusive. Carvalho et al. 19 undertook a comparative study by feeding zebrafish larvae with Artemia nauplii, a commercial starter feed, a semi-purified diet, and a practical diet during the first 21 days of exogenous feeding. Feeding dry feeds right from first feeding, they could achieve good growth of zebrafish, although much lower than that recorded with Artemia nauplii.
In several species of teleosts having very small egg and larvae at first-feeding, studies undertaken on the digestive physiology of larvae besides improved feed processing technologies and adequate feeding strategies have shown that formulated microparticulate feeds can totally replace live prey.20–22,77,79 This has also opened up possibilities for undertaking studies on nutrient requirements with first feeding larvae23–25 or for analysing the metabolic effects of specific nutrients during early development.26–30 The use of microdiets facilitates early weaning of marine finfish larvae and to reduce the reliance on live prey 31 requiring specific enrichment techniques to improve their nutritional values. The objective of this preliminary work was to appraise if microparticulate dry feeds can be used right from first feeding onward to rear zebrafish, D. rerio, under controlled conditions and to get the complete nutrient profile of juveniles produced to generate base line information useful for further studies on the nutrition of this laboratory model species.
Materials and Methods
Zebrafish eggs were obtained from wild-type breeders (ZF WT 2 F 5, Wageningen Agricultural University, Netherlands). Eggs were incubated and larvae were reared in the facilities of the University of Patras, in 10 L aquaria (30 L × 22 W × 18 H cm), which were connected to a 90 L water recirculation system. Initial stocking density was 350 fish per tank (35 individuals L−1); when they reached 15 mm total length (TL), some fish were removed to bring the density to 150 fish per tank (10 L−1). Two days after hatching, triplicate groups of fish started receiving a dry micro-particulate feed (50–100 microns) of commercial origin (Gemma micro®; Skretting). This feed is based on studies undertaken with European seabass. 18 The analytical composition of the feed is provided in Table 1. Feed was distributed by hand to fish four times daily to visual satiety during the larval period (up to circa 12 mm TL) and two to three times daily thereafter. Larvae were eating from both the surface and the water column. After ca 12–14 mm TL, zebrafish started feeding also from the bottom.
Data on vitamins as provided by the feed manufacturer.
DM, dry matter.
Temperature was maintained at 28.0°C ± 1.0°C, pH at 7.0–7.5, oxygen saturation at 70%–95%, and photoperiod at 14/10 h light/dark. Concentration of ammonia, nitrite, and nitrate was <0.1 mg L−1. Total length was measured periodically, in random samples of only 10–18 fish, to avoid stress to the whole population. Mortality was recorded daily. Data on growth at each sample point were drawn from a representative number of fish from each of the three tanks. Data on survival of fish are reported as the mean and SD of the triplicate groups. At the end of the growth trial, which lasted 9 weeks and after an overnight fast, 100 fish from each of the triplicate aquaria were sampled and kept frozen in liquid nitrogen for further analyses.
Whole-body samples of 300 fish were analyzed for dry matter (DM, 110°C, 24 h), ash (550°C, 18 h) crude protein (CP = Kjeldahl nitrogen × 6.25) were performed following AOAC (1995) 32 procedures. Total lipid was extracted according to Folch et al. 33 and quantified gravimetrically. Neutral lipid (NL) and polar lipid (PL) were separated as per Juaneda and Rocquelin, 34 using cartridges (Sep-Pac, Waters) and the two fractions were quantified gravimetrically.
FA composition of diets and of fish was determined on both the NL and phospholipid fractions. FA methyl esters (FAME) were prepared by acid-catalyzed transmethylation of total lipids for 16 h at 50°C, using tricosanoic acid (23:0) as an internal standard. 35 FAME were extracted, purified, and then separated by a Hewlett-Packard 5890A Series II gas chromatograph equipped with a chemically bonded (PEG) Supercowax-10 fused silica wall coated capillary column (30 m × 0.32 mm i.d.; Supelco Inc.), using an on- column injection system and flame ionisation detection. Injector and detector temperature was, respectively, 240°C and 260°C. Hydrogen was used as the carrier gas and the oven thermal gradient was 50°C to 180°C at 25°C/min, 180°C to 235°C at 3°C/min, and then a constant temperature of 235°C for 10 min. FAME were identified by comparison with known standards and quantified by means of a direct-linked computer and Hewlett-Packard ChemStation software.
The protein-bound amino acid (AA) composition was analyzed in samples after acid hydrolysis (6 M-HCl, 1108C, 23 h). After evaporation, samples were analyzed in an automatic amino acid analyzer (Biochrom-30; Biochrom Ltd) using a sodium high-resolution protein hydrolysate column (resin ultra pac 8). The AAs were derivatized with ninhydrin and quantified using norleucine as an internal standard. This method, however, does not allow us to detect the levels of tryptophan. Minerals and trace elements were analyzed after acid digestion of ash samples by the Institut Européen de l'Environnement de Bordeaux (IEEB, Bordeaux), using inductivity coupled plasma analytical procedures.
Results and Discussion
Data on the chemical composition of the dry micro-particulate diet used are reported in Table 1. Since this diet contained a high level of phospholipids, the NL and PL fractions were separated and the FA profiles of these fractions are also provided in Table 2. The feed used here has already been successfully used for rearing larvae of European sea bass 18 as well as in Asian sea bass. 31 Based on mainly marine ingredients and rich in soybean phospholipid, the diet was rich in 18:2 n-6 (41%), but contained close to 14% of n-3 FA, with 5% 18:3 n-3 (linolenic acid [LNA]), 3 and 4% each of 20:5n-3 (eicasopentanoid acid) and 22:6 n-3 (docosohexanoic acid [DHA]) respectively, resulting in a n-3/n-6 ratio of 0.3. Calculated per 100 g feed, the n-3 supply amounted to more than 1.3%.
During the whole growth trial, survival was 85%–92%. Data on growth measured as TL (mm) of zebrafish over the 9 weeks is reported in Figure 2. Compared to the data from the only previous study where zebrafish were fed a dry feed right from first feeding, 19 fish growth was much higher in the present study and comparable to that observed with Artemia nauplii. The growth of zebrafish here was also much higher than that reported by Siccardi et al., 14 who reared zebrafish for the first 28 days post-fertilisation with rotifers and fed different dry feeds. A reference growth curve of zebrafish reared on Artemia was recently provided by Gomes-Requeni et al. 36 When compared to this reference curve, our data are very similar during the first month post-hatch, but then the growth retarded after week 6. Such a reduction in growth with the onset of maturation has been reported 37 and also with significant differences between males and females.36,38 We did not make any distinction between males and females in our study. Our data also show a much higher growth rate for first feeding larvae than that reported for zebrafish or even for the giant danio by Biga et al. 38 reared with live organisms. Goolish et al. 39 compared different diets and found that the growth of zebrafish was lower with dry feeds as compared to live feeds. Although comparisons of growth rates between studies is difficult to make due to possible differences between strains, rearing conditions etc, this is, as far as we know, the first ever demonstration of such a high survival and the best growth performance recorded with first-feeding zebrafish reared only with a dry compound feed. The whole-body composition of zebrafish at the end of the growth trial (Table 3) showed that protein, fat, and ash contents were 16%, 10%, and 2.5% of wet weight, respectively. Data on whole-body composition of zebrafish as regard AAs, FA, mineral, and trace elements are reported in Tables 4 to 6.

Total length (mm) of zebrafish reared from first feeding onward with a dry formulated feed.
From Tacon et al. (1984).
Ideal AA profile
The AA composition of the whole body of zebrafish (Table 4) is similar to that recently reported for this species 36 as well as with those of other cyprinids. It is known that there is a good correlation between the whole-body AA profile and that of the requirements for the same species. 40 Based on an analysis of data available on essential AA (EAA) requirement of different species as well on the whole-body AA profiles of several species of fish, Akiyama et al. 41 found close similarities in the essential AA ratios (A/E ratios) of the whole-body AA profiles of closely related species. Gatlin 42 compared the EAA profiles of goldfish (Carassius auratus), golden shiners (Notemigonus crysoleucas), and fathead minnows (Pimephales promelas) and found that they had similar AA patterns in whole-body tissue. By computing the A/E ratios, he compared their data with the quantitative data on AA requirements of the common carp 43 and found them to be very similar. Similar observations were also made by Mohanty and Kaushik 44 when they did a comparison between another group of cyprinids, the Indian major carps. Based on data obtained here and those of Gomes-Requeni et al. 36 , an attempt has been made in Table 4 to provide the ideal protein profile suitable for zebrafish. As already stated for other species of fish,42,44 we should recognize that this can well serve only as a starting point, awaiting more precise quantitative data on the individual EAA requirements of zebrafish.
Essential FAs
Given the very good growth of the larvae of zebrafish, based on data available, we can speculate that the essential FA (EFA) requirements were met by this feed supplying about 1.3% of n-3 FA and about 4% of n-6 FA, in comparison to what is known for other cyprinids.45,46 In common carp, the very early works of Takeuchi and Watanabe 47 already showed that the dietary requirement of carp was around 1% both of linoleic (LA) and LNA, but that there was a supplemental effect of 0.5% ω3-LC PUFA on carp growth exceeding that of 1% 18:3ω3. In a study dealing with the EFA requirements of first feeding larvae using semi-purified dry feeds, Radunz-Neto et al. 24 observed that a dietary level of total n-6 FA level of 0.9 to 1% was sufficient and that the n-3 FA requirement would possibly be extremely low (<0.1%). They also found that the FA composition of carp larvae reflected that of the diets and that carp larvae were capable of elongating and desaturating LNA and LA to longer chain FAs. In zebrafish, we do not have any information on the quantitative EFA requirements. Meinelt et al. 5 found that in young and old larvae, the greatest length was attained by those fed the diet with an n-3/n-6 ratio of 0.4 to 0.6, corresponding to an n-3 levels of 1.5% to 2% and n-6 levels of 3.3% to 4.1% of the diet. The diet used here had about 4% n-6 FA and that of n-3 PUFA was 1.4% (with LNA, eicasopentanoid acid, and DHA being 0.6%, 0.3%, and 0.4%, respectively).
Data on the FA profiles of the NL and PL fractions of the whole body of zebrafish are reported in Table 5. The PL fraction represented about 15% of the total lipids. Saturated and monounsaturated FAs represented about 30% each of the total FA in the total and NLs. The proportion of LA (18:2 n-6) was higher in the NL fraction than in the PL of zebrafish whole body. The levels of monoenes were reduced (24%) in the PL fraction. Arachidonic acid (20:4 n-6) and DHA levels in the PL fraction was almost two-fold higher than in the NL. Dietary FA profiles and the FA profiles of NL and PL fractions were computed per unit 100 g feed or 100 g wet weight and the relations analyzed. As presented in Figure 3, there was a very good and significant correlation between dietary FA profiles and the whole-body FA profiles. The correlation coefficient was, however, slightly reduced for the relation between dietary FA and whole-body PL FA profile. A rough analysis suggests that the relatively high level of DHA in the PL fraction reflected specific retention of this EFA in the PL. This is in conformity with the aforementioned observations of Radunz-Neto et al. 24 in the common carp larvae.
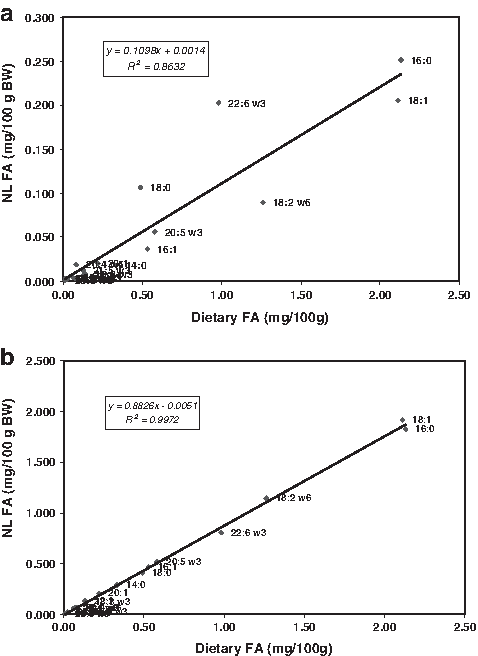
Relation between dietary FA profile and whole-body FA profile of neutral lipids
From a metabolic point of view, FA bioconversion capacities have been analyzed to some extent. Zebrafish have been shown to possess an unique desaturase possessing both delta-6 and delta-5 activity along with an elongase with a very high C18-20 activity. 48 Jaya-Ram et al. 9 found that dietary FA profiles affected different (liver, muscle, ovary, and eggs) tissue FA profiles; there was increased expression of hepatic desaturase and elongase genes in zebrafish fed diets with low levels of long-chain PUFAs (LC-PUFAs). Similarly, data from Karanth et al. 10 in adult zebrafish show that dietary FA profiles, besides affecting tissue FA profiles, had an effect also on the expression of genes of FA binding proteins. Monroig et al. 49 found that the genes encoding enzymes involved in LC-PUFA biosynthesis are expressed right from the beginning of embryogenesis, possibly implying maternal mRNA transfer to the embryo. This implies that care should also be taken toward reproductive performance as affected by dietary factors.
Minerals and trace elements
There is increasing interest for studying skeletal development or bone mineralization, morphology, and structure 50 or on specific trace element metabolism 51 in zebrafish, but the nutritional background of these animals is generally not clearly defined. Although it is known that dietary mineral levels affect bone formation and growth in almost all teleosts, there is no information available on the whole-body mineral or trace element concentrations of zebrafish or to the dietary supply and availability. The effects of dietary versus water-borne zinc on developmental pathways in the gill were followed by Zheng et al., 12 who also reportedly measured tissue trace element concentrations (data not provided). Adult zebrafish are capable of accumulating water-borne iron 52 and dietary iron reduces water-borne copper-induced tissue copper load and several genes involved in copper transporters. 78 Goldsmith et al. 53 showed that the overall nutritional status (fasted vs. fed) affected caudal fin development in zebrafish, without any specific consideration to the dietary supply of minerals or trace elements. The involvement of dietary strontium on increasing bone mineral density has also been shown by Siccardi et al. 54
Shearer 55 stated that in the absence of sufficient quantitative data on mineral or trace element requirements, the ability of a diet to supply essential elements can be assessed by comparing the whole-body elemental levels of fish fed a given diet to that reported as normal for a fish of equivalent size and that if the whole-body concentration of a particular element is less than normal, fortification of the diet will be required. The whole-body mineral and trace element composition of zebrafish fed the dry micro-particulate diet over 9 weeks is reported in Table 6. For comparative purposes, data on the composition of juvenile common carp 56 are also provided in this table. Satoh et al. 57 reported that the whole-body mineral composition of developing fish changes markedly over the first weeks of life and becomes stable later on. The comparison made in the table between adult zebrafish and juvenile common carp, however, shows that although there is reasonably good similarities as regard the whole-body macro-mineral contents, the trace element concentrations vary much between these two cyprinids. The tissue concentrations of trace elements are very much influenced by dietary factors as well as by possible uptake from the aquatic environment. 58 By giving diets containing different levels of mineral or vitamin premixes to first-feeding common carp larvae, Alami-Durante et al. 59 found that larvae fed a non-supplemented diet had a lower survival rate and final mean body weight than those fed supplemented feeds. Similar work with zebrafish fed graded levels of a given mineral or trace element in a complete diet would strongly help studies devoted to developmental biology. Nevertheless, a point of concern with regard to current husbandry practices with zebrafish is that they are generally reared in recirculating culture systems where there is a possibility of accumulation of water-borne minerals and trace elements, affecting osmotic balance 58 as well as embryogenesis and early development, as already shown in the common carp larvae. 60 Indeed, dissolved minerals in water can form complexes with inorganic and organic ligands with possible adverse effects of extremely high levels of even essential trace elements on aquatic organisms need our attention. 61
Vitamins
We did not measure the individual vitamin levels in the feeds or in the fish. According to the manufacturer's specifications and to our own personal knowledge, the diet used contained all the different fat-soluble and water-soluble vitamins at levels beyond the recommendations of NRC 45 for rainbow trout. These recommendations are based on a series of studies as summarized by Woodward, 62 who also compared the water soluble vitamin requirements of fish, chicken and mammals showing great similarities even between phylogenetically distant species. These recommendations have further been confirmed to be valid for other finfish species as well. 63 The good growth rate with no deformities (data not shown) strongly indicate that the dietary vitamin supply was adequate even for the first-feeding zebrafish larvae. In first-feeding common carp larvae, a total lack of vitamin supply has been shown to lead to high mortalities and low growth. 59 Similar findings were made by Mazurais et al. 64 in first-feeding European seabass (Dicentrarchus labrax) larvae fed a diet totally devoid of a vitamin mixture. They found that with increasing the dietary vitamin mixture levels in the feeds, there was improved survival, skeletal development and morphogenesis of the larvae. More work is needed to precisely define the optimum levels of the different vitamins and also to address issues such as whether zebrafish like carp larvae require a dietary supply of vitamin C or not25,65 and what would be the optimal dietary level of retinoids for improved reproductive performance of zebrafish. 7
Carbohydrates
We also did not give any specific attention to carbohydrate fraction of the feeds since in almost all teleosts, there is no defined dietary carbohydrate requirements. 45 Studies already undertaken with broodstock zebrafish also show that there is possibly sexual dimorphism in carbohydrate utilization. 11 There are indications that we can possibly induce long-term metabolic changes in carbohydrate utilization in teleosts through early feeding high carbohydrate diets. 66
General implications
Lieschke and Currie 1 listed a number of characteristics in zebrafish having similarities as well as differences to humans. In fact, in that list, almost all the points can also be attributed to other teleosts and not specific only to zebrafish: Whether knowledge gained with zebrafish can be applied to other teleosts, especially the hundreds of species of finfish that are farmed, is also a bit questionable. For instance, zebrafish, like other cyprinids, are functionally stomachless and our knowledge on the distribution of digestive enzymes, their activities, and functional roles in zebrafish is also limited. Nevertheless, given the particularities of early development and facility with which they can be reared or maintained and with which we can intervene at the cellular or molecular levels within the whole organism, zebrafish remains a model of choice for basic research. Developing a standard diet for zebrafish will largely expand the horizon of actions as regard the analysis of the effects of potential factors under controlled nutritional conditions.
Meinelt et al. 67 observed that the fertilization rate of zebrafish eggs was reduced with increased dietary n-3/n-6 ratios. Markovich et al. 68 found that performance of zebrafish broodstock (7-month-old fish, 20 days of feeding) was linked with the FA profiles of the feeds, as in most teleosts. By feeding 22-week-old female zebrafish (mean individual mass: 260 mg), semi-purified diets containing different oils over 10 weeks, Jaya-Ram et al. 9 showed that dietary LC-PUFAs improved the reproductive performance of zebrafish. An important part of analyzing any diet is to assay reproductive performance of fish. Although this was not formally done, preliminary observations (data not shown) indicate that this diet supports good reproductive performance, showing that it is possible to complete the life cycle with zebrafish fed a formulated feed. However, further work is needed to tailor the nutritional status of male and female zebrafish, given the possible sexual dimorphism in carbohydrate utilization in broodstock zebrafish 11 and given the increasing interest toward nutritional programming.30,66
Despite the interest for zebrafish as a model for studies on nutrition or nutrigenomics of other farmed finfish species, 69 the potential has not yet been realized. 3 The first bottleneck is that we do not have any solid information on the nutrient requirements of this cyprinid.4,69 Our knowledge in this area limited to a summary report showing a correlation between dietary n-6 LC-PUFA and growth rate. 5 From a qualitative point of view, a dietary requirement for retinoids has been reported. 7 The dietary supply of riboflavin, a water-soluble vitamin, can be improved through special delivery vectors. 6 There is a need for water-borne boron for normal embryogenesis of zebrafish 8 as in rainbow trout 70 or higher animals, including humans. 71 Since zebrafish is a cyprinid, one can possibly assume that the requirements for macronutrients of zebrafish can be drawn from what is known for the common carp or other cyprinids. Data on the quantitative requirements of common carp, a cyprinid, are relatively well known for most of the nutrients43,72 and, even then, mostly for juvenile stages. Nevertheless, most of the basic work undertaken with zebrafish deals with the early developmental stages, for which zebrafish is duly recognized as a model for exploring metabolic pathways or physiological consequences as affected by environmental factors, including feed. Even with larvae of other finfish, especially those producing small eggs and consequently small larvae, from a practical point of view, the production of juveniles in hatcheries is still very much based on the use of a sequence of live preys as feed, with or without specific enrichment procedures of the prey items. Recognizing that the most challenging period of larviculture of zebrafish is the first-feeding phase, Best et al. 73 have recently shown that, by using specific rotifers, zebrafish can be reared with high rates of growth and survival, but the majority of live prey used for rearing larval fish are deficient in one or several essential nutrients and they require different enrichment techniques to improve their nutritional values. 74 Hamre et al. 75 showed that rotifers might not be able to meet the requirements of marine fish larvae for several micronutrients, including vitamins and trace elements. It was recognized that the real breakthrough for precisely defining the nutrient needs during the vital developmental stages will be the formulation of nutritionally complete, palatable particulate diets for larvae, 76 with the production of a steady line of larval diets for basic research as well for simplifying hatchery operations.
Most finfish larvae have the ability to digest and thrive on compound dry feed if this diet has the correct physical and palatability attributes and is nutritionally well balanced. 22 Data obtained here show that this is also possible with zebrafish showing excellent growth and survival fed a compound feed all through the life cycle. We initiated this preliminary work to provide some base line information on the nutrient requirements. Further work should focus on developing a standard semi-purified feed for controlled nutrient deliveries in studies with zebrafish.
Footnotes
Disclosure Statement
No competing financial interests exist.