
Abstract
Abstract
Lead (Pb2+) affects neuronal and endocrine systems that influence social interactions. By providing potential hiding locations, spatial heterogeneity may affect Pb2+-induced behavioral outcomes. Therefore, a test chamber was designed into which a refuge could be inserted. The refuge allowed test subjects to escape from the mirror image that stimulated agonistic interactions. Behaviors with a mirror were compared with baseline activity patterns without a mirror. Adult (12-month old) male and female zebrafish, exposed to Pb2+ (0–10 μM) as embryos (2–24 hours post fertilization), were tested individually for 5 min in each chamber design within 2 h of feeding. Behaviors were evaluated for % time in mirror zone, distance traveled (=activity level), and attacks on the mirror image. When there was no refuge, significant concentration-dependent increases occurred in male % time in mirror zone, activity level, and number of attacks. Increases in these variables were less pronounced in females. When there was a refuge, there were significant differences for males only in activity level and attacks at the higher developmental exposure concentrations; % time in mirror zone followed a similar pattern and level as without refuge. Females displayed Pb2+-induced behavioral changes only for attacks on mirror. Since the presence of refuges that is, environmental enrichment, reduced Pb2+-induced agonistic behavior in both sexes, experimental spatial design can be considered an important factor when interpreting behavioral outcomes.
Introduction
Social behavior, particularly agonistic interactions, has a strong sex-linked basis in mammals29,30 but is more complicated in fishes because sex determination is not purely based on the presence of sex-linked chromosomes, for example, temperature, sex ratios, and pheromonal cues may be determinants of individual sex status. 31 Thus, it is possible that changes in social behavior as a result of exposure to environmental contaminants such as Pb2+ may not be as straightforward. A further complication, and one that is investigated in the studies described in this article, is the experimental apparatus used to elicit specific behavioral outcomes. It has been observed that environmental enrichment can mitigate the learning deficits caused by developmental Pb2+ exposure, 32 suggesting that the design of the testing chamber or protocol may affect behavioral outcomes. From this perspective, mirrors have been used as means to stimulate social interactions in many vertebrate species, including fishes.33–39 This experimental design is effective because of the strength of visual sensory inputs to stimulate exploratory and social behaviors. Specifically, mirrors allow a test subject to use vision alone to guide its use of space and spatial preferences inside a closed arena within a short time frame. 40
Since environmental stimuli produce specific behavioral responses, it is possible that constant stimulation, at least over the short time scales used in this study, induces greater behavioral intensity. Conversely, if there is a location within the test chamber in which the test subject can find “respite” from the source of stimulation, that is, the mirror, it is possible that the break in stimulation, regardless of however short a time frame, alters the response intensity toward the mirror image. This is important, because Pb2+ has been associated with aggressive behavior. 41 Therefore, a separate goal of this study was to determine whether a change in experimental design alters the behavioral outcomes that are usually associated with this contaminant. This is a particularly important question, as it affects how researchers compare data from other laboratories and our understanding of the mechanisms of behavioral responses to environmental contaminants in nature outside the confines of a controlled, experimental laboratory aquarium or cage.
Materials and Methods
Treatment of glassware and plasticware
All laboratory materials made of plastic were washed thoroughly in a 10% solution of a nontoxic, biodegradable detergent (Simple Green™; Sunshine Makers, Inc., Huntington Harbour, CA), rinsed repeatedly in ultrapure Milli-Q™ water (Millipore Corp., Medford, MA), and immersed in a 30 mM Na4EDTA (Fisher Scientific, Hanover Park, IL) solution overnight to remove all surface adsorbed metal ions; glassware was washed and rinsed similarly but immersed in a 10% HNO3 (Fisher Scientific) solution overnight. Glass and plasticware were then rinsed in ultra-pure Milli-Q water.
Breeding and egg collection
Adult zebrafish were from a wild-type laboratory strain originally acquired as aquarium fish from EkkWill Waterlife Resources (Gibsonton, FL). Fish were maintained at 26–28°C on a 14-h light and 10-h dark cycle in a flow-through, dechlorinated water system at the Aquatic Animal Facility of the University of Wisconsin-Milwaukee Children's Environmental Health Sciences Core Center. All experimental procedures were approved by the University of Wisconsin–Milwaukee Animal Care and Use Committee. Zebrafish were bred in 2-L plastic aquaria with a 1/8′′ nylon mesh false bottom to protect fertilized eggs from being consumed by the adults.
Eggs were collected ≤2 hours post fertilization (hpf), counted, and placed in metal-free, glass culture dishes (100 mm diameter×50 mm depth; N=100 eggs/dish) in E2 medium (each liter contains 0.875 g NaCl, 0.038 g KCl, 0.120 g MgSO4, 0.021 g KH2PO4, and 0.006 g Na2HPO4).
Exposure regimen
A stock solution of 1 mM PbCl2 in ultrapure water (MilliQ Synthesis A10; Millipore Corp.) was prepared at pH 2 until fully dissolved. Using 1.0 M KOH, pH was adjusted to 6.5. Eggs were rinsed twice in Pb2+-free E2 medium [as determined by inductively coupled plasma mass spectrometry (ICP/MS) analysis] and transferred (N=100 eggs/dish; <2 hpf) to a metal-free glass dish (100 mm diameter×50 mm depth) containing 100 mL of E2 medium with Pb2+ at 0.0, 0.01, or 0.03 μM. At 24 hpf, eggs were rinsed thrice with Pb2+-free E2 medium and placed in clean, metal-free glass dishes with Pb2+-free E2 medium. After 1 month, larvae were placed in 1-L plastic, flow-through tanks using dechlorinated Lake Michigan tap water. At 4 months of age, fish were transferred to 3-L plastic, flow-through tanks at a density of 12 fish/tank. Previous studies demonstrated Pb2+ uptake values in 24 hpf embryos. 3 One month before testing, fish were separated by sex by feeding them Hikari Micro Pellets (Kyorin Co., Ltd, Himeji, Japan); with this protocol, the ventral side of the males produce an orange hue.
Testing chamber
A white, plastic box (16 cm width×25 cm length×10 cm height) was constructed with interchangeable panels to allow for multiple testing designs. In the first test (Fig. 1A), a white plastic wall was inserted to split the chamber lengthwise so that each section of equal area would hold a single fish, and each fish was unable to detect the presence of the other visually or by odor. The 10 cm height was necessary to prevent the fish from jumping out of the chamber. At one end of each section, a mirror was inserted; the opposite end remained without a mirror. An individual male was placed in one section and a female was placed in the other so that each sex was tested together within the same time frame. Each section was divided into three zones: 0–1 cm from the mirror (=mirror), 1–12.5 cm from the mirror (=transition), and 12.5–25 cm from the mirror (=opaque). These three zones represent different levels of relationship to the mirror image.

Social behavior test chambers. Male and female fish are separated by white, opaque barrier (——). Zone boundaries (——) are computer generated and are used only for spatiotemporal analyses. Gray boxes=mirror.
In the second test (Fig. 1B), a white plastic wall was inserted to split the chamber widthwise so that each section of equal area would hold a single fish, and each fish was unable to detect the presence of the other visually or by odor. As done earlier, an individual male was placed in one section and a female was placed in the other so that each sex was tested together within the same time frame. In each section, a refuge with no mirror was created by a partial wall (5 cm length×10 cm height) that prevented the fish from seeing its image in the mirror. Each section was divided into four zones of equal area: on the side with the mirror 0–5 cm from the mirror (=mirror) and 5–12.5 cm from the mirror (=mirror-away); on the side without the mirror 0–5 cm from the end (=refuge) and 5–12.5 cm from the end (=refuge-away). To test the effect of the mirror itself on behavior, the experiment was repeated except that the mirror was not inserted. Therefore, fish activity was not responding to the visual stimulation provided by the mirror.
Fish movements were recorded at a rate of 30 frames/s with an infrared-sensitive digital camera (Ikegami Model ICD-49; Ikegami Tsushinki, Co., Ltd, Tokyo, Japan) placed 45 cm above the bottom of the test chamber. Lighting was provided by a ring (diam. 10 cm) of 18 white light LEDs (EnvironmentalLights.com, San Diego, CA) placed above the test chamber to provide even lighting intensity (200 lux) throughout the chamber to allow the fish to see the mirror image. Lighting for the camera was provided by an infrared lamp (IR—ROOM Ultra-Covert 940 nm Infrared Illuminator; Night Vision Experts.com, Buffalo, NY) placed 45 cm above the bottom of the test chamber.
Testing protocol
All tests used dechlorinated Lake Michigan water warmed to 28°C in an incubator. This represents the same water conditions used for maintaining the adult fish. Water (1 L, 2.5 cm depth) was replaced after each trial to maintain equal temperatures between trials and to remove any olfactory clues left by the previous fish. Before replacing with fresh water, the chamber was rinsed thrice with distilled water. To reduce the potential effects of circadian activity rhythms affecting results, all tests were conducted within 2 h of feeding time (0930 h).
Mirror test without refuge
Individual male and female adult zebrafish (12-months old; N=12 of each sex) were placed into separate sections of the testing chamber and allowed to acclimate for 1 min before recording started. Fish movements were monitored for 5 min. Using image analysis software (EthoVision v8.5; Noldus Information Technology, EthoVision XT Reference manual, Version 8.0, Wageningen, The Netherlands, 661 pp. Wageningin, The Netherlands), time spent in each zone (mirror, transition, and opaque), number of attacks on the mirror image, and total distance traveled (cm) were recorded.
Mirror test with refuge
Individual male and female adult zebrafish (12-months old; N=12 of each sex) were placed into separate sections of the testing chamber and allowed to acclimate for 1 min before recording started. Fish movements were monitored for 5 min. Using the image analysis software, time spent in each zone (mirror, mirror-away, refuge, and refuge-away), as well as all of the variables mentioned earlier, were recorded.
Statistical analysis
Since two fish died during the experimental protocol, a regression approach (concentration×presence of mirror×sex) was used to perform the statistical analysis for each chamber design (refuge vs. no refuge) as opposed to a three-way analysis of variance. Interactions, and within and between treatment differences for presence of mirror, concentration, and presence of mirror×concentration were subjected to regression analysis with t-test with significance established at p<0.05. Since the level of social interaction was best defined by the degree of association with the mirror (time spent, distance traveled, and number of attacks), these analyses were confined to the mirror zone only, that is, analyses for zones not near the mirror (transition and opaque zones for experiments without a refuge and mirror-away, refuge-away, and refuge for experiments with a refuge; Fig. 1) were not included. Correlation coefficients (r2) of the regression equation were calculated to determine the percent of total variation attributable to each variable under conditions of either with or without the refuge.
Results
Since data were analyzed using several comparisons, for example, within and between sexes, between concentrations, with versus without mirror, and with versus without refuge, statistical significance of these comparisons is summarized in Tables 1 and 2 rather than using a confusing array of symbols in each graph. Correlation coefficients for each variable are found in Table 3.
Level of significant set at p<0.05. Bold numbers indicate statistical significance.
NS, treatment group not significantly different from control group; T, t statistic; df, degrees of freedom.
Level of significant set at p<0.05. Bold numbers indicate statistical significance.
NS, male behavior not significantly different from female behavior; T, t statistic; df, degrees of freedom.
Mirror test without refuge
No mirror
In tests without a refuge, there were no significant differences between exposure regimens for time spent in the mirror zone regardless of sex when no mirror or refuge was present with the exception of an increase among females at the lowest exposure concentration (p<0.05, T=−2.73, df=21; Table 1; Figs. 2 and 3). A similar lack of significant differences were observed in both male and female zebrafish for distance traveled (Table 1; Figs. 4 and 5), and number of attacks into the zone where the mirror would be placed (Table 1; Figs. 6 and 7). The only exception to this result occurred at the lowest developmental exposure concentration (0.01 μM Pb2+) in which females were in the mirror zone for more time than males and males had more attacks into the mirror zone than females (Table 2).
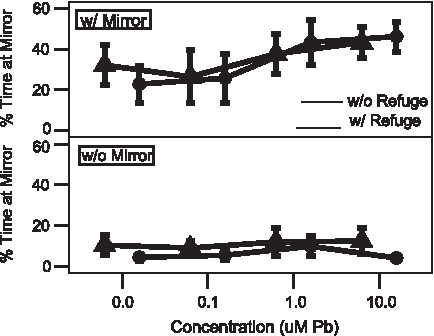
Time spent by adult male zebrafish in mirror zone with and without a refuge, and with and without a mirror in the test chamber as a function of developmental Pb2+ exposure. The spatiotemporal distribution of adult (1-year old) male zebrafish is dependent on the presence of a mirror and developmental exposure to various concentrations of Pb2+ (0–10.0 μM). The presence of a refuge does not change that behavior. Mirror at top, without mirror at bottom. ▴——▴=Without refuge in test chamber. ●——●=With refuge in test chamber. Values=mean±2 standard error (SE).

Time spent by adult female zebrafish in mirror zone with and without a refuge, and with and without a mirror in the test chamber as a function of developmental Pb2+ exposure. The spatiotemporal distribution of adult (1-year old) female zebrafish is dependent on the presence of a mirror and developmental exposure to various concentrations of Pb2+ (0–10.0 μM). The presence of a refuge does not change that behavior. Mirror at top, without mirror at bottom. ▴——▴=without refuge in test chamber. ●——●=with refuge in test chamber. Values=mean±2 SE.
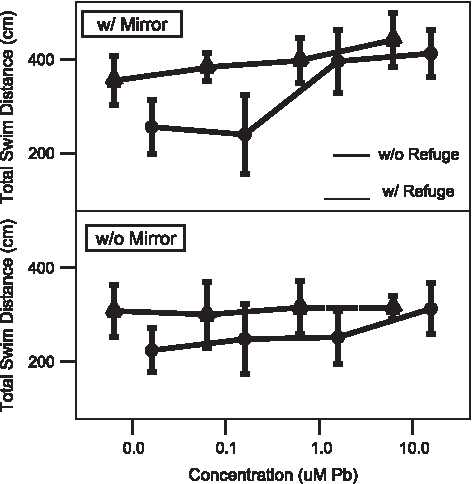
Activity level in test chamber of adult male zebrafish as measured by distance traveled during observation period with and without a refuge, and with and without a mirror in the test chamber as a function of developmental Pb2+ exposure. The total distance traveled by an adult (1-year old) male zebrafish is dependent on the presence of a mirror and developmental exposure to various concentrations of Pb2+ (0–10.0 μM). The presence of a refuge changes the expression of that behavior. Mirror at top, without mirror at bottom. ▴——-▴=without refuge in test chamber. ●——●=with refuge in test chamber. Values=mean±2 SE.

Activity level in test chamber in adult female zebrafish as measured by distance traveled during observation period with and without a refuge, and with and without a mirror in the test chamber as a function of developmental Pb2+ exposure. The total distance traveled by an adult (1-year old) female zebrafish is dependent on the presence of a mirror and developmental exposure to various concentrations of Pb2+ (0–10.0 μM). The presence of a refuge does not change the expression of that behavior. Mirror at top, without mirror at bottom. ▴——▴=without refuge in test chamber. ●——●=with refuge in test chamber. Values=mean±2 SE.

Agonistic activity of adult male zebrafish as measured by number of attacks on the mirror image with and without a refuge, and with and without a mirror in the test chamber as a function of developmental Pb2+ exposure. The number of attacks on the mirror by an adult (1-year old) male zebrafish is dependent on the presence of a mirror and developmental exposure to various concentrations of Pb2+ (0–10.0 μM). The presence of a refuge decreases the expression of that behavior at all exposure regimens. Mirror at top, without mirror at bottom. ▴——▴=without refuge in test chamber. ●——●=with refuge in test chamber. Values=mean±2 SE.

Agonistic activity of adult female zebrafish as measured by number of attacks on the mirror image with and without a refuge, and with and without a mirror in the test chamber as a function of developmental Pb2+ exposure. The number of attacks on the mirror by an adult (1-year old) female zebrafish is dependent on the presence of a mirror and developmental exposure to various concentrations of Pb2+ (0–10.0 μM). The presence of a refuge decreases the expression of that behavior at all exposure regimens. Mirror at top, without mirror at bottom. ▴——▴=without refuge in test chamber. ●——●=with refuge in test chamber. Values=mean±2 SE.
With mirror
Males spent a greater amount of time in the mirror zone when the mirror was present; this increase was also affected by concentration level (t-test, p<0.001, T=−2.87, df=21; Table 1; Fig. 2), particularly at the highest exposure levels (1 and 10 μM Pb2+). Males increased the distance traveled during the observation period (∼400 cm) compared with tests without the mirror (∼300 cm) with significant, concentration-dependent differences at the highest exposure regimen (t-test, p<0.05, T=−4.06, df=20; Table 1; Fig. 4); females displayed no differences between mirror and no mirror or between exposure regimens (∼300 cm in both cases; Fig. 5). The number of attacks on the mirror was significantly higher when the mirror was present for both sexes and showed a concentration-dependent increase (t-test, p<0.005, T=−3.86, df=13; Table 1; Figs. 6 and 7), although males displayed greater increases versus females (Table 2).
Mirror test with refuge
No mirror
In tests with a refuge, there were no significant differences between exposure regimens for time spent in the mirror zone with a refuge zone when no mirror was present (Figs. 2 and 3), distance traveled (Figs. 4 and 5), or number of attacks at the site where the mirror would be placed (Figs. 6 and 7).
With mirror
When the experimental design was altered so that there was a place within the testing chamber where the fish could not see its image in the mirror (Fig. 1B), there no significant, treatment-dependent differences observed for percent time in the mirror zone for females (Table 1, Fig. 3); males displayed a concentration-dependent increase (t-test, p<0.001, T=−4.00, df=20; Table 1, Fig. 2). There were no significant, treatment-dependent differences observed for total swim distance for females (Table 1). Male zebrafish showed significant increases in swimming distance at the highest exposure regimen (p<0.05, T=−2.22, df=21; Table 1). Inclusion of a refuge resulted in a decrease in the number of attacks on the mirror image for both males and females (Figs. 6 and 7). Although females displayed no differences between Pb2+ exposure regimens (Table 1; Fig. 7), males showed a greater increase versus females in number of attacks with increasing developmental Pb2+ (t-test, p<0.005, T=−3.86, df=11; Table 2; Fig. 6 vs. Fig. 7) and for distance traveled (t-test, p<0.05, T=2.74, df=20; Table 2; Fig. 4 vs. Fig. 5).
Discussion
Two fundamental questions were posed in this study: (1) do developmental exposures to an environmental contaminant induce life-long changes in behavioral outcomes and (2) to what extent can environmental enrichment, as created through spatial heterogeneity, mitigate those effects? Since critical neural wiring occurs early in zebrafish embryo development, and male and female brain organization is influenced by sex steroids, it was hypothesized that contact with potential endocrine disruptors such as Pb2+ at this early life history stage would result in sex-related changes in adult social interactions, specifically agonistic behaviors that include aggression as well as ritual, threat, or submissive displays. Pb2+ was examined because of its role in altering social behaviors 26 and previous work conducted in our laboratory identifying behavioral changes due to low-level (nM–μM) exposures to this metal.3,21,42,43
In adult zebrafish used for this study, the presence of a mirror alone stimulated increased activity for both males and females when compared with a test chamber with no mirror. We used this baseline of no mirror in the test chamber to investigate the degree to which the behavioral changes observed were due to an interaction between toxicant-induced hypersensitive responses and the stimulation caused by the mirror image. Mirrors have been shown to alter anxiolytic activity in mice. 37 Importantly, there were clear sex-related differences in the patterns of developmental Pb2+ exposure-produced responses in the adult zebrafish, although the specific patterns varied depending on whether a refuge was provided, paralleling studies conducted using rats.19,44
Due to its association with aggressive behavior, we were particularly interested in potential differences between males and females with the number of attacks on the mirror; Pb2+ has been linked to increased aggression in experimental animals and humans.19,21,44,45 Males showed a greater number of attacks versus females at higher exposure regimens, (1.0 and 10.0 μM Pb2+), while females displayed a greater number of attacks versus males only at the lower treatment level (0.1 μM Pb2+).
The presence of a refuge, that is, spatial heterogeneity, represented a section of the test chamber in which the fish can escape visual contact with the mirror (Fig. 1B). This environmental enrichment design provides four spatially distinct zones that the fish is able to choose: direct close encounters with an “opponent” (=mirror zone), less intimate association but still within visual contact, no visual contact but within a close range of re-establishing such interaction, and no visual contact and requiring greater swimming activity to enter the zone with such stimulation.
While females displayed no significant differences in time spent in the mirror zone, number of attacks on the mirror image, or distance traveled during the trial as a function of developmental Pb2+ exposure (Figs. 3, 5, 7), males demonstrated significant shifts in all these variables (Figs. 2, 4, 6), suggesting that the intensity of the attacks on the mirror image was mitigated by the presence of the refuge. By providing areas in which there were sections free from any visual stimulation by the mirror image, there was a reduction in attention given and level of response to the source of stimulation. Not all studies, however, indicate that environmental enrichment decreases aggression. Using Nile tilapia, Barreto et al. 46 found that environmental enrichment, by providing more resources worth defending, actually increased aggression. However, spatial heterogeneity that is not resource based may create a different framework for agonistic behaviors. The ability of spatial variation to mitigate adverse behavioral outcomes was also observed in two separate studies with rats exposed to significantly higher gestational concentrations of Pb2+ than the embryonic exposures used in this study. In each experiment, an enriched environment reversed Pb2+-induced long-term potentiation impairment or spatial learning deficits.9,32
The low correlation coefficients (Table 3) suggest that additional confounding factors were responsible for causing the observed variation in our data. First, individual position within the social hierarchy was not tracked. Not only do physiological, morphological, and behavioral measures affect social status in fishes but the reverse is also true.47–49 Furthermore, the behavioral effects of toxic exposure will affect individual fish differently depending on their social rank. 50 Without correlating the social status of individual fish with their behavioral responses, we likely grouped individuals with inherently different relationships to the apparent competitor. Second, since circadian rhythms of agonistic behavior are regulated by feeding schedules,51,52 all testing was within 2 h of feeding time. To assure that all fish from a specific experimental exposure were tested on the same day, observation times were only 5 min. Dahlbom et al. 48 observed that over the course of a 15 min observation period, the first 5 min did not always result in significant differences in social contest outcomes between dominant and subordinate male zebrafish; the last 10 min did. This may imply that establishing social relationships with what is essentially, from the fish's point of view, a similarly sized and sexed opponent takes longer than 5 min and that the variation we observed was a function of that process. Third, our study used adult fish that were developmentally exposed to Pb2+. It is possible that not all fish are equally sensitive to this procedure and that, as adults, their behavior will vary accordingly. Fourth, the highest concentration used in this study was 10 μM Pb2+. It is possible that higher concentrations for an equal exposure time (2–24 hpf) or identical concentrations for longer exposures times, for example, until hatching, may have yielded more significant differences in the results. Longer exposure times may indicate that the period of development most sensitive to alterations of social behavior occurs at a later embryonic stage. Alternatively, prolonged exposures throughout development may be necessary to induce changes in the multiple mechanisms that control this specific behavior but develop at different times during embryogenesis. In spite of all these potential confounders, differences were still observed. It appears that the existence of environmental enrichment indeed has a role to play in decreasing the intensity of concentration-dependent changes in behavior. This seems to apply even to developmental exposures to Pb2+ and long-term behavioral effects.
Lead exposure in humans is associated with irritability (or hypersensitivity to environmental stimuli), hyperactivity, and psychomotor disturbance. All three of these behavioral changes have been observed in our laboratory and can have profound effects on the social interactions observed in this study: Irritability presented itself as greater attraction to the mirror, hyperactivity presented itself as higher activity levels (less time spent immobile and greater distances traveled), and psychomotor disturbance presented itself as altered startle response behaviors (e.g., Rice et al. 3 ). Hyperactivity is also a known risk factor for antisocial and aggressive behavior. 41 While it cannot be claimed that Pb2+ specifically caused increased aggression, it clearly was a driver for increases in the more global behavioral category of agonistic interactions.
The hypothesis that developmental Pb2+ exposure affects adult social behavior by altering sex steroid levels and/or activity is partially supported by research examining the role of estrogen during development and Pb2+ interactions with that process. In fish, both estrogen and progesterone are important in embryonic development and stimulation of the brain-pituitary axis during reproductive maturation.53–55 Assuming that the social behaviors evaluated in this study are hormonally based, the differential responses between adult male and female zebrafish as they interact with their mirror image, that is, another “animal” of same sex and size, suggest that developmental Pb2+ exposure may act as an endocrine disruptor of adult social behavior.
Each of the two estradiol receptor (ER) subtypes, ERα and ERβ, play a role in controlling aggressive behavior. While a lack of the ERα gene results in decreased aggression, a lack of the ERβ gene leads to increased aggression. 56 Lead-gene interactions during early embryonic development have been previously identified26–28 and a lack of gene expression due to a mutation can be related to blocked gene expression due to Pb2+ binding. Therefore, it should be noted that human granulosa cells cultured with 10 μM Pb2+, the high concentration level for the zebrafish study reported here, had decreased cytochrome p450 aromatase and ERβ activity presumably through down-regulation of both P450 aromatase and ERβ gene transcription. 57 Although mature, cultured gonadal tissue may react differently to Pb2+ than the embryonic brain, these observations on Pb2+ effects on sex steroid gene expression provide a working hypothesis as to why adult zebrafish display sex-specific agonistic behavioral responses as a result of developmental Pb2+ exposure.
As a practical application of this study, the design of a behavioral test chamber can affect the outcomes and, therefore, interpretations of an experiment. It is, however, in the comparison of both experimental designs that new insights into potential mechanisms of toxicity may be gleaned.
Footnotes
Acknowledgments
This project was funded by the Children's Environmental Health Sciences Core Center (NIEHS grant ES04184, David Petering, director). Randall Metzger of the UW-Milwaukee Great lakes WATER Institute Machine Shop is gratefully acknowledged for his assistance in improving the original design and in constructing the behavioral apparatus used for this study.
Disclosure Statement
No competing financial interests exist.