
Abstract
Abstract
The infraorbital (IO) bone series, a component of the circumorbital series, makes up five of the eight dermal bones found in the orbital region of the zebrafish skull. Ossifying in a set sequence, the IOs are closely associated with the cranial lateral line system as they house neuromast sensory receptors in bony canals. We conducted a detailed analysis of the condensation to mineralization phases of development of these bones. Our analyses involved both bone and osteoblast staining of zebrafish at 20 different time points. IO bone condensations are shaped as templates for the final bone shape, and they mineralize at one or more centers of ossification. Initially, mineralization is closely associated with the lateral line canals and/or foramen, and the onset of mineralization is temporally variable. Canal wall mineralization is a process that continues into adulthood and completely mineralized canal roofs were not found. Our comprehensive growth series detailing the ossification of each IO bone provides important insight into the growth and development of this series of neural crest-derived flat bones in the zebrafish craniofacial skeleton.
Introduction
T
The zebrafish (Danio rerio) 3 circumorbitals make up six of the eight dermal bones found in the orbital region of the skull. 4 The remaining two dermal orbital bones are the median parasphenoid and the paired frontals. 4 The circumorbital bones comprised different groups named according to their location around the orbit. The supraorbital bone is located dorsally above the orbit, the antorbital is located anterior to the orbit, the suborbital is located ventral to the orbit, and the postorbitals are located posterior to the orbit.5,6 Infraorbital one (IO1; antorbital), infraorbital two (IO2; suborbital), and infraorbitals three to five (IO3–IO5) (postorbitals) make up the IO bone series (Fig. 1).
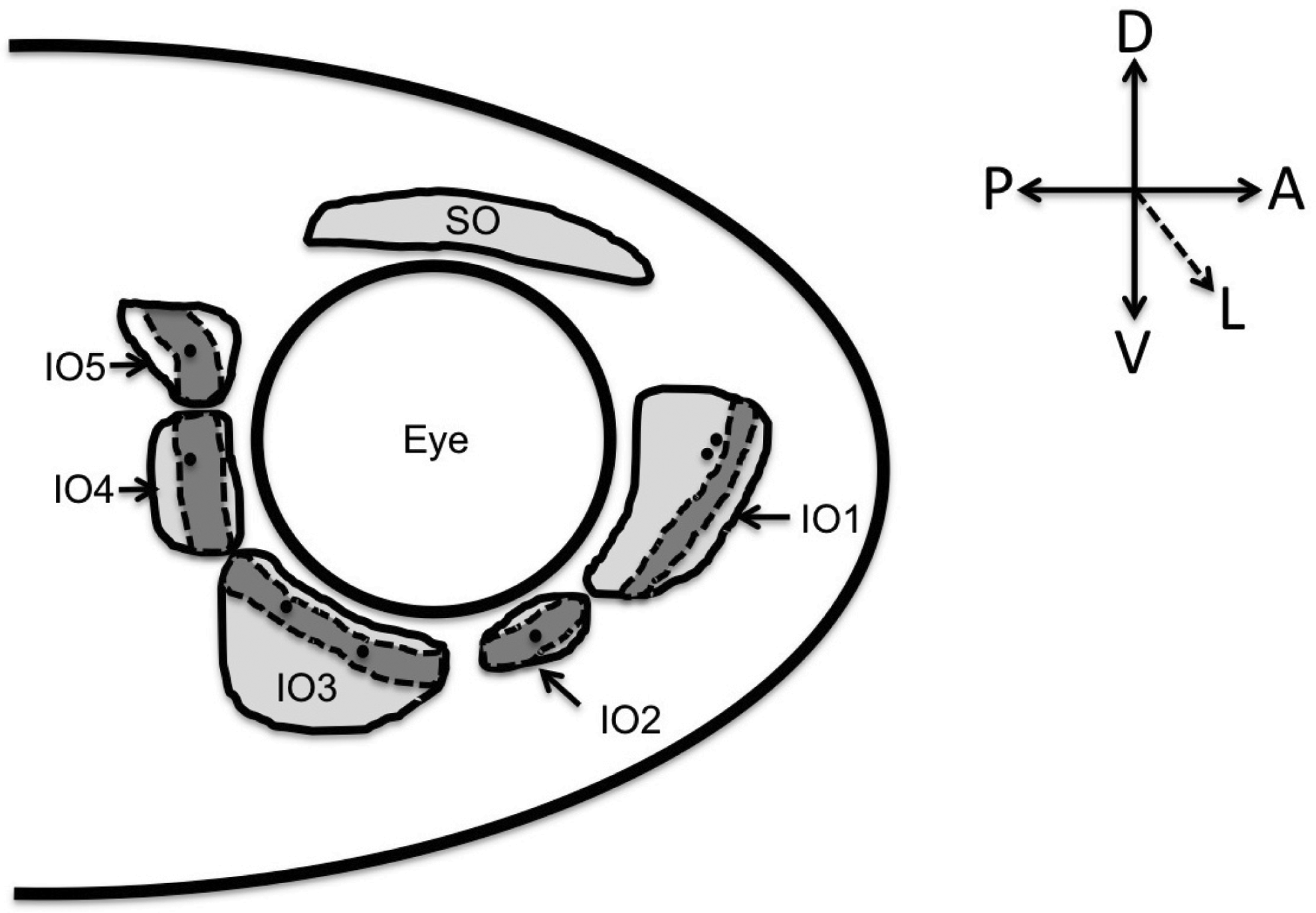
A schematic of the zebrafish circumorbital bones composed of the dorsal supraorbital (SO) bone and the infraorbital (IO) bone series. Infraorbital one (IO1) (i.e., the lachrymal bone) is in the antorbital position, IO2 (i.e., the jugal) is found ventral to the orbit in the suborbital position, and IO3 through IO5 are found as the three posterior postorbital bones. The IO series of bones consists of the antorbital, suborbital, and postorbital bones only. The canal walls (dashed line) and canal portion (dark shaded area) are distinguished from the flat bone component (light shading). Neuromast foramen are indicated by dark circles.
The IO series of bones are found in generally the same position in all teleosts. 1 Similarities are found in the arrangement and/or patterning of the IO bones, as well as in the distribution of neuromast sensory receptors that are found in lateral line canals associated with these bones. 1 The primitive teleost condition is to have seven IO canal bones beginning anteriorly with the antorbital and continuing posteriorly to the dermosphenotic. 1 A reduction in the number of IO bones occurred during the evolution of Teleostei; 1 within Cypriniformes, the family which includes the zebrafish, the reduced number of IO bones is likely due to fusion of the suborbital bones. 1
All of the zebrafish IO bones are neural crest derived 7 and develop intramembranously. 4 They develop late within the craniofacial skeleton, and ossify in a set sequence. 4 The first IO bone to mineralize is IO1, followed by IO3 and IO5 concurrently, and then IO4 and finally IO2, which is also the last bone in the zebrafish skull to ossify. 4 The IO bones are sequentially labeled according to the anatomical position: starting anteriorly and moving posteriorly, 4 this numbering therefore does not match their temporal sequence of development.
The zebrafish IO bone series is a complex and interesting set of neural crest-derived dermal bones. These bones share a close association with one another, developing in a set temporal order and eventually form a connective tissue attachment to one another. How does each IO bone acquire its adult morphology? How many centers of ossification do IO bones possess? Are all IO bones the same? Does the cranial lateral line system play a role in IO bone development? These are some of the questions we sought to answer.
Cubbage and Mabee's highly cited description of the zebrafish skull currently provides the most comprehensive description of the development of the zebrafish IO bones to date. Detailed descriptions of the timing of IO condensations and the growth and shape change of developing IOs have yet to be described. In this study, a detailed analysis of the condensation (premineralization) to mineralization phases of development of these bones is conducted. Osteoblast and bone staining of zebrafish at 20 different time points during zebrafish ontogeny were used to assess the mineralization and growth of these bones as well as the ossification of the lateral line canal roof.
Materials and Methods
Wild-type zebrafish (D. rerio) were bred in our laboratory from AB stocks from the Zebrafish International Research Center (ZIRC) and reared following standard conditions. Specimens were collected at 20 developmental time points from ∼21 days postfertilization up to 4 months of age (n=5 for each stage). These developmental time points were based on the specimen size rather than age, as size has been previously shown to be a more reliable indicator of developmental stage in fish. 8 The developmental time points used for the growth series range from 6.0 to 25.0 mm standard length (SL), and were based on the timing of IO development observed by Cubbage and Mabee. In addition, zebrafish that had survived up to 3 years of age were also examined (30 mm SL, n=5). Animal care protocols were approved annually according to the guidelines of the Canadian Council on Animal Care.
Samples were anesthetized with 0.01% MS222 and measured to the nearest 0.5 mm using a Nikon SMZ1500 microscope. Following SL measurements, samples were euthanized in 0.1% MS222. Samples for osteoblast (alkaline phosphatase [AP]) staining were fixed in 4% paraformaldehyde (PFA) in phosphate-buffered saline (PBS), pH 7.4, overnight at 4°C and samples for bone staining were fixed in 10% neutral buffered formalin (NBF) overnight at room temperature. Following fixation, the specimens fixed in 4% PFA were transferred to 0.01 M PBS for storage at 4°C and the specimens fixed in 10% NBF were processed to 70% ethanol for storage at room temperature.
For analysis of IO bone condensations, AP staining was utilized to identify active osteoblasts. 9 A more concentrated, AP substrate solution (2%) was used instead of the recommended 1%. For analysis of the growth and shape change involved in IO mineralization, bone staining was conducted using the standard protocol for Alizarin Red.10,11 To analyze the mature IO canal structure, 3-year-old zebrafish specimens were double stained with Alizarin Red and Alcian Blue following standard procedures 11 with the adjustment of the staining procedure being split into two separate steps. 12
IO condensations and bones were analyzed both in situ and following dissections using a Nikon SMZ1500 stereomicroscope and Nikon 50i eclipse compound microscope. IOs were photographed using a Nikon DXM 1200C camera and measurements were taken using Nikon NIS elements software.
Results
Here, a detailed description of the development and growth of each of the five IO bones follows, starting at its first detection with whole-mount staining and ending with a description of the element in adult fish at 4 months and at 3 years of age.
Infraorbital one
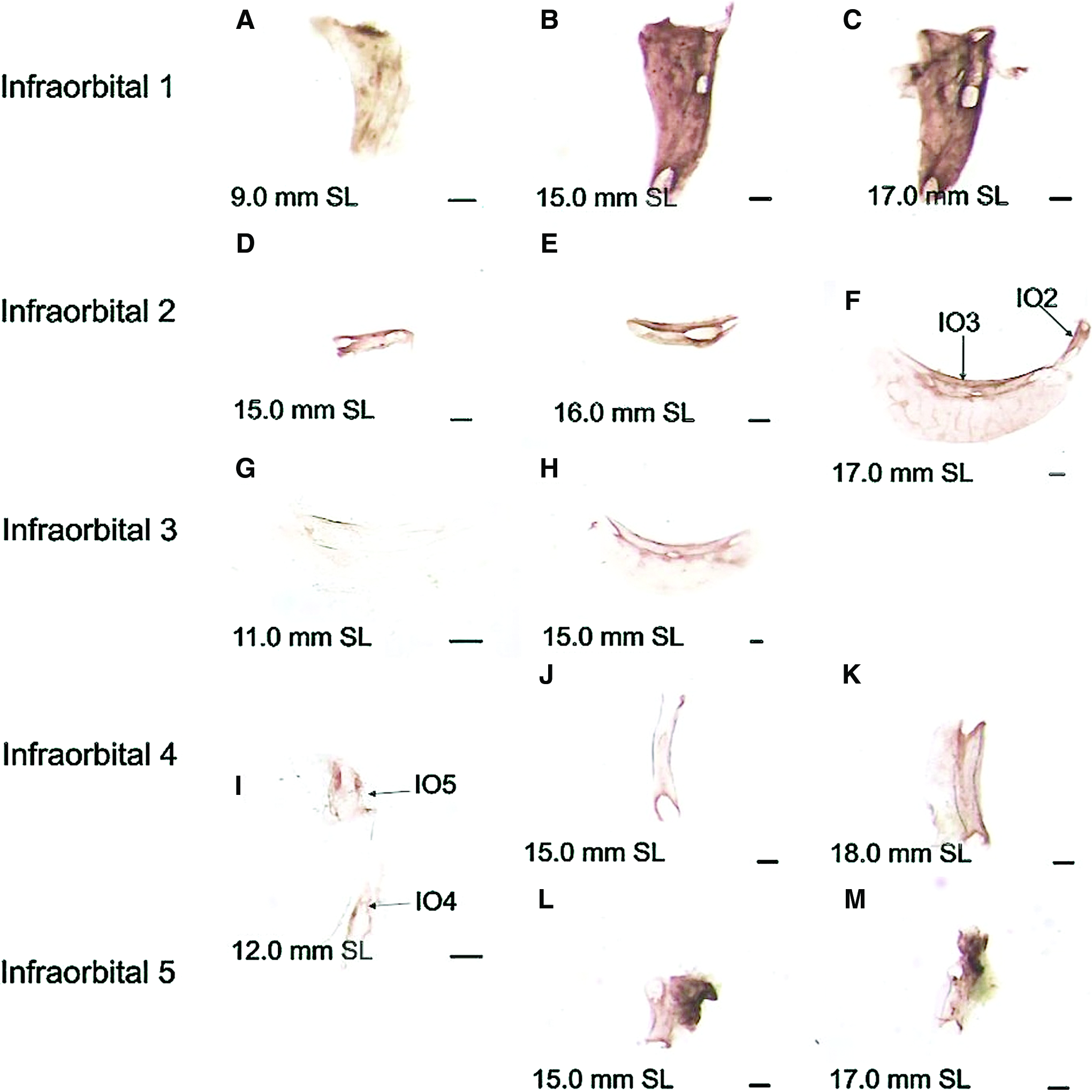
IO1 is first detected at 9.0 mm SL using AP staining; the staining is present along the posterior side of the lateral line canal and along the dorsal edge of the presumptive IO1 bone (Fig. 2A). At the same stage, this element does not stain with Alizarin Red (Fig. 3A), however, the shape of the IO1 bone can be deduced based on the background of AP staining. This presumptive IO bone has a similar, although miniaturized, morphology to that of the mature mineralized IO1 bone (compare Figs. 2A and 3E). Mineralization of IO1 is first observed at 10.0–11.0 mm SL as a single sliver of bone along the posterior side of the lateral line canal and extending around a central foramen (Fig. 3B). With continued growth, the zone of AP staining (Fig. 2B, C) and mineralization (Fig. 3C, D) broadens, extending to the edges of the presumptive IO1 bone. The dorsal portion of the bone mineralizes before ossification and is present in the ventral half (Fig. 3C). At 15.0 mm SL, the lateral line canal walls begin to mineralize as the posterior canal wall extends out (laterally) and eventually closes over toward the anterior portion of the canal to form the canal roof (Fig. 3D). Canal closure continues throughout IO1 mineralization. Initially, the canal roof closure occurs only in the central portion of the canal length. In mature 3-year-old zebrafish, the lateral line canal of IO1 is still not completely closed along the canal length (Fig. 4A, B).
Alkaline phosphatase (AP) staining in the developing zebrafish IO bones.

Alizarin Red staining shows mineralization in the developing zebrafish IO bones.

The IO bones in a mature 3-year-old zebrafish.
Infraorbital two
IO2 first appears as a short canal-shaped structure posteroventral to IO1; it can be detected with AP activity at 15.0 mm SL (Fig. 2D). Beginning at 16.5–18.0 mm SL, mineralization of IO2 can be observed with Alizarin Red and begins ventrally (Fig. 3F). Interestingly, IO2 can have one (n=3) or two (n=2) ossification centers (Fig. 3F, G); these two centers enlarge and eventually fuse with one another. The location of the singular ossification center when compared with bones of two centers of ossification is located in the midpoint between the two centers. At 20.0 mm SL, the canal walls begin to ossify outward (dorsolaterally) with the ventral canal wall mineralizing faster than the dorsal wall; both the flat and canal portion of IO2 are stained at this stage (Fig. 3J). In mature 3-year-old zebrafish, the IO2 canal is still not mineralized to closure and the canal walls are not completely fused (Fig. 4C, D).
Infraorbital three
IO3 is first observed using AP staining, which is present in the area of the presumptive lateral line canal, at 11.0 mm SL (Fig. 2G). AP staining then expands along the length of the presumptive canal (Fig. 2H); later, osteoblast activity is observed in the flat portion of the presumptive IO3 (Fig. 2F). This area of AP activity corresponds to the mineralization pattern observed in the IO3 bone stained growth series (Fig. 3K–O). Ossification first occurs along the lateral line canal of the presumptive IO3 bone with mineralization subsequently expanding into the flat portion of IO3 (Fig. 3K–M). Eventually, the canal walls begin to mineralize outward (laterally) (Fig. 3M–O). The dorsal canal wall grows out as a single unit, whereas the ventral canal wall expands in several segments. The dorsal canal wall also mineralizes before the ventral wall. In mature specimens, the canal walls approach one another, but never meet and canal roof mineralization is incomplete (Fig. 4E–F and K–L).
Infraorbital four
IO4 is first observed at 12.0 mm SL as a canal-shaped element that stains positive for AP (Fig. 2I). This AP activity expands along the length of the canal-shaped element and then extends as a flat projection on the posterior side of the element at 18.0 mm SL (Fig. 2J–K). The AP staining corresponds to the pattern of IO4 mineralization (Fig. 3P–T). At 11.0–16.0 mm SL, mineralization begins at the base of the canal-shaped element and expands along the length of the presumptive IO4 bone (Fig. 3P–R). Canal walls are observed at 16.0 mm SL and ossify outward (laterally) in segments starting at 16.5 mm SL (Fig. 3S, T). The onset of ossification of the flat wing-like extension of IO4 is variable and was observed as early as 16.0 mm SL. The canal walls begin to form the roof of the canal beginning at 18.0 mm SL. In mature specimens, fusion of the canal walls in the roof has not occurred (Fig. 4G, H).
Infraorbital five
The IO5 is first observed at 12.0 mm SL as a short canal-like AP-positive stained element (Fig. 2I). At 15.0 mm SL, this AP staining is intense and has expanded in a flat anterodorsal direction (Fig. 2L). Once again, the AP activity corresponds to the mineralization pattern observed in the Alizarin Red stained specimens. IO5 is an unossified element in specimens ranging from 11.5 to 13.0 mm SL. Ossification of IO5 starts as a single ossification center surrounding a canal foramen (Fig. 3U). By 14.0 mm SL, mineralization expands along the entire presumptive canal length (Fig. 3V). The flat wing-like extension of the IO5 bone is apparent by 17.0 mm SL (Fig. 3W–X). At 22.0–25.0 mm SL, the lateral line canal walls begin ossifying outward in adjacent segments and over the canal roof, such that the anterior wall precedes the posterior wall (Fig. 3Y). In mature 3-year-old zebrafish, the canal walls have ossified toward each other, but are still not fused (Fig. 4I, J).
Discussion
As previously reported, the onset of ossification in the IO bones is specific to each bone in the series. 4 Our data show a similar sequence of ossification to that previously reported, however, the precise timing of ossification differs, most likely due to differences in growth rates of our fish.
IO bones exhibit two mineralization patterns
In all the bones examined, AP staining was observed before staining with Alizarin Red. Interestingly, after our observations of AP expressing skeletogenic condensations, but before Alizarin Red staining of mineralized bones, an unstained presumptive IO bone element that is similar, but smaller, in shape and size to that of the mineralized structure is visible and easily dissected (for all IO bones). The nature of this unstained tissue is likely to be osteoid, since AP staining detects active osteoblasts and preosteoblasts. 12 Skeletogenic condensations are known to be the building blocks on which the skeleton is built13,14 and these skeletogenic condensations must reach a critical size before ossification, a process that requires both genetic and mechanical regulation.13,15 Interestingly, in IO2–5, the initial area of AP activity only includes the canal portion and excludes the flat wing-like extension of the bone. That is, in these bones, subsequent mineralization begins in the canals before the AP activity is observed in the flat wing-like extensions. The IO1, however, is different. It has AP staining and mineralization in the flat component of the bone before the onset of canal formation. Thus, two patterns of ossification for IO bones are observed and emphasize the complexity of IO bone development as well as the importance of the two different roles that these parts of the IO play in the zebrafish: namely, the canals enclose the sensory lateral line system, while the flat wing-like extension provides structural support and protection to the orbit and face. In addition, the functional demands of IO1 are different from the other IO bones as the first IO bone to mineralize; it first serves a protective and structural role.
IO bones develop in a robust sequence with temporal variability
The observed sequence of IO bone mineralization agrees with the order previously described by Cubbage and Mabee, namely, infraorbital one, infraorbital three and five concurrently, infraorbital four, and infraorbital two. Previous work on the sequence of ossification of the zebrafish skull shows that there is a low level of intraspecific variation in skull ontogeny consisting of small sequence shifts; however, the sequence shifts observed did not affect the order of IO development. 16
Overall, the onset of IO ossification occurred later in zebrafish development in our study when compared with Cubbage and Mabee. However, this inconsistency in the timing of IO ossification between studies is most likely due to the differences in the wild-type zebrafish strains used by each research group. Additionally, specimens in the previous study were measured postfixation and thus have experienced shrinkage during fixation. 17
The temporal onset of mineralization of each IO bone differed between specimens. IO1, IO2, and IO4 display the most variability in the timing of mineralization onset. Variability in the timing of developmental events (i.e., ossification) is not uncommon and has previously been reported in fish development studies. The onset, duration, or offset of developmental events in fish can be influenced by nutrition,18,19 water velocity, 20 and temperature. 21 Of these, the studies that investigated the development of the endoskeleton found that the sequence of ossification was robust, but the timing was variable.20,21 Additionally, bones that mineralize later in craniofacial development were more affected by external environmental factors (temperature and mechanical loading) than those that develop earlier.20,21 The variable IO mineralization onset observed in this study could similarly be a consequence of environmental factors; even zebrafish reared in the same tank experience variable environments due to individual differences in food intake and/or preferred location in the water column.
Further investigation into the numerous environmental factors affecting the development of the zebrafish skull is required to fully understand the complexities of craniofacial growth and development in teleosts.
IO bones begin mineralizing at one or more centers of ossification
In this study, we found that IO1 and IO3–5 ossify from a single ossification center, whereas IO2 can have one or two ossification centers. This is in contrast to Cubbage and Mabee who reported two centers of ossification for IO1 and one ossification center for IO2–5. These findings suggest that there is some variability in how IO1 and IO2 ossify in zebrafish. It is interesting that the two IO bones that have the potential to ossify from two centers are the first and last in the IO series to mineralize; they are also the largest (IO1) and smallest (IO2) bones in the series. One would expect more than one ossification center in larger bones or in bones that need to ossify quickly (where the rate of matrix production cannot be increased or is at a maximum). IO1 mineralizes early and forms a flat-shaped bone in the antorbital cavity that overlies the nasal gland and maxilla. One could speculate that IO1 has an important function to protect these structures requiring its growth to occur quickly. IO2 is the last bone in the skull to form and two ossification centers would allow the bone to mineralize quickly at the end of zebrafish skull development.
The cranial lateral line is closely associated with IO bone development
There has been a large informal debate regarding whether the cranial lateral line plays a role in the development of the dermal bones in the teleost skull, specifically the development of the lateral line canal walls.6,14,22–26 Studies agree that the sensory components of the IO lateral line (i.e., the neuromasts) are present well before the IO bones ossify.24–27 Furthermore, the enclosure of these neuromasts into bony canals occurs during IO bone development. 26 Neuromast induction of the lateral line canal wall ossification is thought to be caused by either an epithelial tissue interaction,14,26 a biochemical environmental change, 23 or a mechanical trigger. 22 In this study, we show an association between the lateral line sensory system and IO bone ossification. With the exception of IO1, both the osteoblast activity and bone mineralization within the canal component of IO bones precede the presence of the flat wing-like extension of each bone. Interestingly, the IO lateral line sensory system lies adjacent to or passes directly through the ossification centers of mineralizing IO bones (Fig. 3) such that there is always a close association between the two. In addition, IO ossification first begins around canal foramen in the case of IO5. The role the lateral line plays in IO development requires more resolution and is currently under investigation.
The IO canal roof is incomplete
Although it has been suggested that the neuromasts are enclosed in a mineralized bony canal,4,5,26 we never observed the complete ossification of the canal roof in any of the IO bones, even in 3-year-old zebrafish.
In all these bones, the pattern of canal roof mineralization included the raising of canal walls. During this wall rising, the canal walls closest to the orbit ossified before the other side. Only after the walls reach a certain height do they begin to fold toward the contralateral wall to form the canal roof. Interestingly, we observed that the tissue between the two edges of these approaching canal walls (i.e., the canal roof region) stains prominently with Alcian Blue (Fig. 4K, L) suggesting that the tissue is rich in glycosamingylcans or muccopolysaccharides. 14 Detailed histological analysis is required to fully determine the composition of this tissue within the roof of the IO canals of zebrafish.
In summary, the zebrafish IO bones are a complex series of neural crest-derived dermal bones. Located around the underside of the zebrafish orbit, the IO bones house the cranial lateral line system and play a structural, supportive and protective role as a component of the skull. We describe important aspects of IO development through detailed observation. These include the temporal variation in ossification both between IO bones and between different specimens, the presence of multiple ossification centers in the largest IO bones, the formation of the lateral line canals, and the canal roof. Our study highlights the need for further investigation exploring the signaling events preceding and leading up to condensation formation in this series, and the role, if any, of the lateral line system in the development of these bones.
The IO bone series is a constituent of the circumorbital bones surrounding the orbit, and because of their close association with the lateral line system and the eye, their development is of interest to researchers investigating each of these tissues and also those interested in the interactions between these hard and soft tissues during development. Additionally, zebrafish IO bones can now be compared with those of other teleosts (e.g., Astyanax mexicanus 28 ) in the study of skull changes over evolution. More detailed descriptive analyses, however, are required to determine if similar patterns of mineralization and lateral line canal wall formation are observed in other teleost orders.
Footnotes
Acknowledgments
The authors would like to thank the Nova Scotia Health Research Foundation (to T.F.O.) and the Natural Sciences and Engineering Research Council of Canada for funding (to T.F.O.). Finally, we would like to thank the Mount Saint Vincent University and Saint Mary's University, the fish care personnel and members of the Franz-Odendaal Bone Development Lab for their ongoing support.
Disclosure Statement
No competing financial interests exist.