
Abstract
Abstract
The inhibitory avoidance paradigm allows the study of mechanisms underlying learning and memory formation in zebrafish (Danio rerio Hamilton). For zebrafish, the physiology and behavior associated with this paradigm are as yet poorly understood. We therefore assessed the effects of environmental enrichment and fish age on inhibitory avoidance learning. Fish raised in an environmentally enriched tank showed decreased anxiety-like behavior and increased exploration. Enrichment greatly reduced inhibitory avoidance in 6-month (6M)- and 12-month (12M)-old fish. Following inhibitory avoidance, telencephalic mRNA levels of proliferating cell nuclear antigen (pcna), neurogenic differentiation (neurod), cocaine- and amphetamine-regulated transcript 4 (cart4), and cannabinoid receptor 1 (cnr1) were lower in enriched-housed fish, while the ratios of mineralocorticoid receptor (nr3c2)/glucocorticoid receptor α [nr3c1(α)] and glucocorticoid receptor β [nr3c1(β)]/glucocorticoid receptor α [nr3c1(α)] were higher. This was observed for 6M-old fish only, not for 24-month (24M) old fish. Instead, 24M-old fish showed delayed inhibitory avoidance, no effects of enrichment, and reduced expression of neuroplasticity genes. Overall, our data show strong differences in inhibitory avoidance behavior between zebrafish of different ages and a clear reduction in avoidance behavior following housing under environmental enrichment.
Introduction
T
Experiment 1: environmental enrichment
Laboratory housing of fish all too often is designed to standardize conditions between experiments and therefore typically consists of barren aquaria. However, housing rodents under such conditions is considered to lead to poor welfare: animals are often confined to small cages, isolated, and kept from performing highly motivated behaviors and may therefore more likely develop stereotypies. 11 There is consensus that enrichment improves not only animal welfare12,13 but also the quality of research data. 14 Enrichment can be defined as “a combination of complex inanimate and social stimulation” 15 and in this context refers to an environment richer than barren housing conditions. 16 When rodents are housed under enriched conditions, they play more, 17 are less anxious, 18 and perform better in spatial learning and memory tasks.19,20
Typically, zebrafish in laboratory settings are kept in stimulus-poor environments, that is, a tank without bottom substrate, plants, rocks, or other shelter objects. We do know, however, that enrichment affects brain morphology and gene expression 21 and behavior22–27 of fish. Effects of environmental enrichment on inhibitory avoidance behavior in zebrafish have to our knowledge as yet not been studied. We therefore housed zebrafish under enriched or barren conditions from larval stage onward. At 6 months, we first assessed anxiety-like behavior (in a dark–white preference box) followed by the inhibitory avoidance protocol at the age of 12 months. Remarkably, in mice, environmental enrichment increases inhibitory avoidance behavior,6,7 while in rats, enrichment reduces such.8,9 It follows that caution should be taken to predict the effects of environmental enrichment on inhibitory avoidance behavior in zebrafish.
Experiment 2: age
Rats have a window where passive avoidance learning performance is better in adult animals (120–365 days) compared with both young (20–30 days) and aged animals (540 days).28–30 To assess whether age also affects inhibitory avoidance behavior in zebrafish, we studied 6-month (6M), 12-month (12M), and 24-month (24M) old zebrafish in the inhibitory avoidance paradigm. The 24M-old zebrafish are generally classified as “aged” and both the 6M- and 12M-old zebrafish are classified as “adults.” 31 However, at the age of 6 months, zebrafish are still growing in size, 32 and as yet, it is unclear till what age the brain is still developing. We assessed the telencephalic expression of genes related to the stress response, individual differences in coping style, anxiety and fear-conditioning after inhibitory avoidance learning.33–39 We focused on the telencephalon, not the whole brain, as the telencephalon houses centers involved in learning, memory, and fear-conditioning (lateral and medial zone of the dorsal pallium, respectively).40–42 We predict that inhibitory avoidance behavior is reduced in 24M-old zebrafish compared with both 12M- and 6M-old fish.
Experiment 3: interaction of age and environmental enrichment
In aged rats (25 months old), environmental enrichment improved learning up to the level of younger animals (2.5 and 15 months). 43 We hypothesized (experiment 2) that 24M-old zebrafish would have lower brain plasticity and that this would impair inhibitory avoidance learning. Exposure to environmental enrichment could counter this reduction in brain plasticity and thus improve task performance. We housed 24M-old zebrafish under enriched conditions for 4 weeks. As a control, we housed a group of 6M-old zebrafish under similar conditions and assessed their respective inhibitory avoidance behavior.
We hypothesize that enrichment increases inhibitory avoidance behaviour in 24M-old zebrafish, while we cannot make a clear prediction for 6M-old zebrafish (see experiment 1).
Materials and Methods
Ethical approval
All experiments were approved by the Animal Ethics Committee of the Radboud University Nijmegen (RU-DEC 2012-106) and were conducted in agreement with Dutch law (Wet op de Dierproeven 1996) and European regulations (Directive 86/609/EEC).
Housing
Depending on experimental and life stage needs, fish were either housed in small aquaria or large aquaria. Whereas the barren environment consisted of the aquaria as they are, the enriched tanks received sand as bottom substrate and the aquarium was decorated with plastic plants and an artificial rock formation. Enrichment was kept constant (no changes in the amount, type, or location of enrichment) over the course of the experiment.
Small aquaria
Two-liter zebrafish tanks were used to house zebrafish from fertilization up till 1 month. One group was housed under barren conditions, while the other was housed under enriched conditions.
Large aquaria
A single aquarium (100×50×60 cm) was split in two compartments by addition of a black glass plate, creating two equally sized compartments (50×50×60 cm): one barren and one enriched. In each aquarium, the water level was brought to 30 cm, creating a total water volume of 150 L (or 75 L on either side). Both compartments were equipped with their own water in- and outflow (26°C; pH=8.0) and linked to a single biological filter (300 L volume). In addition, each compartment received an aeration stone, through which air was pumped, creating air bubbles and surface motion. The photoperiod was kept at 12-h light:12-h dark (lights on from 07:00 to 19:00 h) with feeding moments at 09:00 h (Artemia salina) and 15:00 h (TetraMin; Tetra, Melle, Germany).
Animals
An in-house-bred Tupfel long fin (also known as Tübingen long fin) of mixed sexes were used. Experiments were carried out at the Radboud University Nijmegen, the Netherlands.
Experiment 1 (enrichment)
A batch of freshly spawned and fertilized eggs was randomly divided into two groups of roughly equal size. Once hatched, fish were either placed in the barren (n=20) or enriched (n=24) environment. Zebrafish larvae were kept in small aquaria until the age of 1 month, after which they were transferred to larger aquaria (see small aquarium and large aquaria section). After 6 months, fish were exposed to a black and white preference test (for details, see black and white preference test section in the Materials and Methods section). After this test, the animals were returned to their housing tanks and left undisturbed for 6 months. At the age of 12 months, we assessed inhibitory avoidance behavior (for details, see inhibitory avoidance behavior section), an age that we used in our earlier study on chronic stress. 5
Experiment 2 (age)
After 6 months (n=22), 12 months (n=35), or 24 months (n=9) of housing under barren (standard) conditions, zebrafish were placed in one compartment of a large aquarium for 4 weeks. After this acclimation period, inhibitory avoidance behavior was assessed.
Experiment 3 (age and enrichment)
Zebrafish, at the age of 6 months (n=16) or 24 months (n=17), were placed in one compartment of a large aquarium for 4 weeks. After this acclimation period, inhibitory avoidance behavior was assessed.
Black and White Preference Protocol
To assess anxiety-like behavior, an aquarium (60×30×30 cm; 10 cm water level) was split in two equal compartments. The walls and bottom of one compartment were covered with white plastic; walls and bottom of the other compartment with black plastic. No physical barrier separated the two compartments so the fish could cross to the other compartment freely.
Zebrafish were placed individually in the white compartment and were allowed to swim around freely for 6 min, after which they were returned to their home-tank. Swimming behavior was recorded by camera and analyzed using EthoVision XT v8.0 tracking software (Noldus, Wageningen, the Netherlands). More time spent in the white compartment is indicative that animals are less anxious. A higher number of crossings between compartments are indicative for more exploratory behavior.
Inhibitory avoidance protocol
To evaluate inhibitory fear avoidance, a protocol was used as previously described. 5 In short, an aquarium (60×30×30 cm; 10 cm water level) was split into two equal compartments, separated by a manually operated sliding door. Surfaces of one compartment and sliding door were white, whereas surfaces of the other compartment and other side of the sliding door were black. Compartments were not covered by a lid. The black compartment contained two metal plates (covered with black sound box mesh to prevent light reflection by the metal) that covered two opposite walls completely. Both plates were wired to a power source, which allowed for 3 V AC (measured at the middle of the tank; across a distance of 15 cm between electrodes) to be put between the plates in the water. Assessment of inhibitory avoidance learning started at 11:00 h and was done for each fish individually (i.e., not as a group).
On day 1, the fish were placed (individually) in the white compartment with the sliding door closed. After 60 s of acclimation, the sliding door was lifted, giving the fish free access to the black compartment. Their innate response, when anxious, is to readily enter the black compartment when given this option.44,45 Once the fish had completely entered the black compartment, the sliding door was closed and an electric shock (3 V AC) was given for 5 s, after which fish were netted and returned to their home tank. Fish that would not enter the black compartment within 180 s on the first day of the paradigm were excluded from further experimentation, as their innate responses were not compatible with the inhibitory avoidance paradigm. For each fish, the latency time to enter the black compartment was recorded. Higher latencies indicate that animals are less anxious in the white compartment.
On day 2, the procedure was repeated. Once the sliding door had been opened after 60 s, the fish was given 180 s to make a choice to enter the black compartment. When the fish entered the black compartment within this time frame, a second shock was given. To give all fish the same number of shocks, fish that did not enter within 180 s were carefully driven into the black compartment with a small net and given the second shock. For each fish, the latency to enter the black compartment was recorded. Higher latencies are indicative for associating the black compartment with the shock and thus increased avoidance behavior.
On day 3, fish were placed within the white compartment once again. After 60 s, the sliding door was opened and the fish were given 180 s to make a choice to enter the black compartment. For each subject, the latency to enter the black compartment was recorded. This time, when the fish entered the black compartment, no shock was given and the fish that did not enter within 180 s were not driven into the black compartment. Rather, fish were netted and kept in a holding tank to be euthanized 2 h after the test (see tissue collection and preparation section).
Operation of the sliding door was done manually, and visual observations were done in real-time and recorded on-site. Recording of the latency time was done by hand using a stopwatch. One experimenter (R.M.) recorded all latencies. All procedures were carried out in a manner that caused the least possible disturbance of fish during the experiment.
Tissue collection and preparation
On the third day of the inhibitory avoidance protocol, fish were euthanized in water containing 0.5% (v/v) 2-phenoxyethanol 2 h after testing (between 14:00 and 15:00 h) in line with our earlier studies (see Manuel et al.4,5 for an extensive discussion of this 2-h time-window and pros and cons of post-task sampling). From each fish, the telencephalon was dissected, snap-frozen in liquid nitrogen, and stored at −80°C until further analysis.
Analysis of gene expression
Genes of interest selected were brain-derived neurotrophic factor (bdnf), proliferating cell nuclear antigen (pcna), neurogenic differentiation (neurod), cocaine- and amphetamine-regulated transcript 4 (cart4), cannabinoid receptor 1 (cnr1), serotonin receptor 1ab (htr1ab), corticotropin-releasing factor (crf) binding protein (crf-bp), glucocorticoid receptor α [nr3c1(α)], glucocorticoid receptor β [nr3c1(β)], and the mineralocorticoid receptor (nr3c2).33–39 Two internal standards (ribosomal protein L13 [rpl13] and elongation factor 1α [elf1α]) were used for normalization. Primer sequences can be found in Table 1.
Elongation factor 1α (elf1α) and ribosomal protein L13 (rpl13) were used as housekeeping genes. Genes of interest were brain-derived neurotrophic factor (bdnf), proliferating cell nuclear antigen (pcna), neurogenic differentiation (neurod), cocaine- and amphetamine-regulated transcript 4 (cart4), cannabinoid receptor 1 (cnr1), serotonin receptor 1ab (htr1ab), corticotropin-releasing factor (crf) binding protein (crf-bp), glucocorticoid receptor α [nr3c1(α)], glucocorticoid receptor β [nr3c1(β)], and the mineralocorticoid receptor (nr3c2).
qPCR, quantitative polymerase chain reaction.
Total RNA from individual telencephalons was isolated using Trizol (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions, including an additional precipitation step with ethanol and sodium acetate. RNA concentration and purity was measured by nanodrop spectrophotometry (Nanodrop; Thermo Fisher Scientific, Waltham, MA). Possible present genomic DNA was removed by treatment with DNase I (Invitrogen) as follows: 1 μL of DNase I (1 U/μL) and 1 μL of DNase I reaction buffer (10×) were added to 8 μL of total RNA (250 ng) and incubated in a total volume of 10 μL at room temperature for 15 min. DNase was inactivated by addition of 1 μL of ethylenediaminetetraacetic acid (25 mM) and incubated for 10 min at 65°C. Subsequently, each sample received 1 μL of random primers (250 ng/μL), 4 μL of first-strand buffer (5×), 1 μL of dNTP mix (10 mM), 1 μL of dithiothreitol (DTT) (0.1 M), 1 μL of RNaseOUT (40 U), and 0.5 μL of superscript II reverse transcriptase (200 U/μL) (all from Invitrogen) and incubated for 10 min at 25°C, followed by 50 min at 42°C and finally 15 min at 70°C. Afterward, cDNA was diluted five times with ultra-pure water and stored at −20°C until analysis. Relative gene expression was assessed by real-time quantitative polymerase chain reaction (qPCR). Briefly, 4 μL of cDNA was used as template in a reaction with 10 μL iQ SYBR Green Supermmix (BioRad, Hercules, CA), 0.7 μL of forward and reverse primer (10 μM each; Table 1), and 4.6 μL of ultra-pure water. qPCR (3 min 95°C, 40 cycles of 15 s 95°C, and 1 min 60°C) was carried out on a CFX 96 (BioRad). Data were analyzed according to Vandesompele et al. 46
Statistical analyses
Statistical analyses were performed with GraphPad Prism 5.0 for Mac (GraphPad Software, Inc., La Jolla, CA).
Experiment 1 (enrichment)
We used a two-way analysis of variance (ANOVA) (with the factors time and enrichment) to test for significant differences in the black and white preference test. In case of a significant interaction, a Bonferroni post hoc test was used. A D'Agostino and Pearson omnibus normality test revealed that the latency times did not conform to Gaussian distribution. Therefore, differences in inhibitory avoidance behavior were assessed by the Kruskal–Wallis test (followed by Dunn's multiple comparison post hoc tests) to test for differences across days and the Mann–Whitney U tests to test for differences between groups on each day.
Experiment 2 (age)
A D'Agostino and Pearson omnibus normality test revealed that the latency times did not conform to Gaussian distribution. Therefore, differences in inhibitory avoidance behavior were assessed by the Kruskal–Wallis test (followed by Dunn's multiple comparison post hoc tests) to test for differences across days and the Mann–Whitney U tests to test for differences between groups on each day. Gene expression data were analyzed for Gaussian distribution using the D'Agostino and Pearson omnibus normality test. In case of a Gaussian distribution, significance between barren and enriched fish was assessed using a one-way ANOVA (with the Student–Newman–Keuls post hoc test); in case of a non-Gaussian distribution, the Kruskal–Wallis test (followed by Dunn's multiple comparison post hoc tests) was used to assess differences between groups.
Experiment 3 (age and enrichment)
A D'Agostino and Pearson omnibus normality test revealed that the latency times did not conform to Gaussian distribution. Therefore, differences in inhibitory avoidance behavior were assessed by the Kruskal–Wallis test (followed by Dunn's multiple comparison post hoc tests) to test for differences across days and the Mann–Whitney U tests to test for differences between groups on each day. Gene expression data were analyzed for Gaussian distribution using the D'Agostino and Pearson omnibus normality test. In case of a Gaussian distribution, significance between barren and enriched fish was assessed using the Student's t-test (with a Welch's correction in case of unequal variances); in case of a non-Gaussian distribution, the Mann–Whitney U test was used to assess differences between groups. In all cases, significance was accepted when p≤0.05 (two-tailed).
Results
Experiment 1: environmental enrichment
Enrichment: black and white preference test
We excluded the first minute from our data analysis, as we believe that this minute reflects a habituation period in which fish recover from entering the experimental tank (Fig. 1A, B).
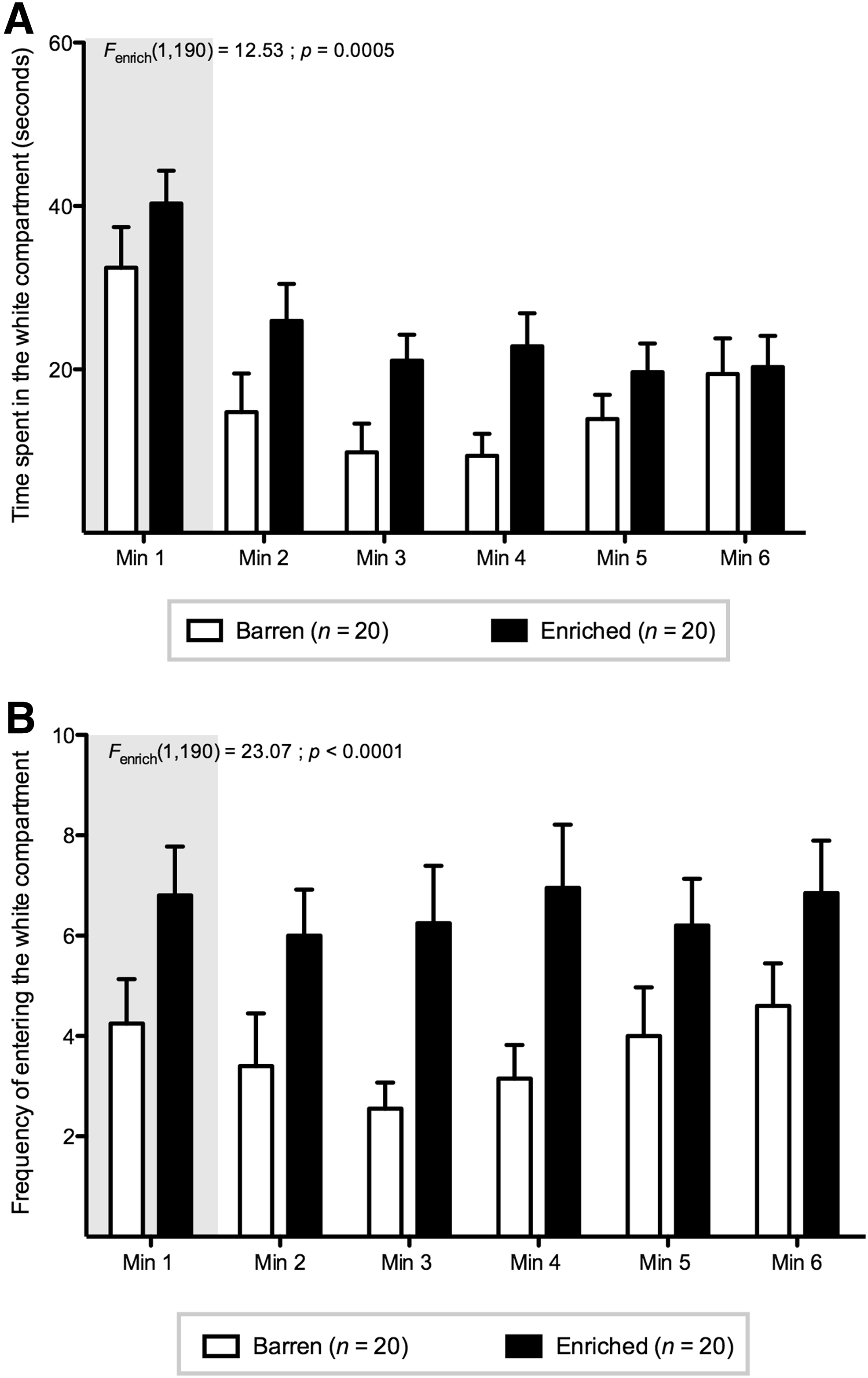
Black and white preference test.
We found that enriched-housed fish spent more time in the white compartment compared with barren-housed fish [Fenrich(1,190)=12.53; p=0.0005]. Furthermore, the number of crossings between compartments was significantly higher for enriched-housed fish compared with the barren-housed fish [Fenrich(1,190)=23.07; p<0.0001]. No other statistically significant differences were found.
Enrichment: inhibitory avoidance paradigm
All fish entered the black compartment within 60 s on the first day, except one individual in the barren-housed group (80 s) and two in the enriched-housed group (90 and 130 s). Median values of groups were 14 and 8 s for barren and enriched conditions, respectively (Fig. 2).

Inhibitory avoidance learning in lifelong barren and enriched house zebrafish. Day 1 (no training) shows no significant differences in the initial latencies between barren-housed (white) and enriched-housed (black) zebrafish. Day 2 (one shock) and day 3 (two shocks) show a significant lower latency for fish housed under enriched conditions compared with those from a barren environment: bars indicate median values. Capital letters above groups indicate significant differences assessed by the Mann–Whitney U test: groups that do not share corresponding letters are significantly different from each other. Triple asterisks (p≤0.001) indicate a significant increase in latency from days 1 to 2 or from days 2 to 3 within a single housing conditions. n.s.=not significant, n=the number of fish.
The latencies of barren-housed fish and enriched-housed fish were not significantly different on day 1 (U=69.50; p=0.52). However, we observed significantly higher latencies for fish raised in a barren environment compared with those raised in an enriched environment on day 2 (U=38.50; p=0.02) and day 3 (U=35.00; p=0.01).
The latencies for barren-housed fish on days 2 and 3 were significantly higher compared with the latencies on day 1 (H=18.25; p=0.0001), but there was no significant difference between days 2 and 3. For enriched-housed fish, we did not observe significant differences (H=5.250; p=0.07).
Experiment 2: age
Age: inhibitory avoidance behavior
All fish entered the black compartment within 60 s on the first day, except two individuals: one in the 6M-old group (95 s) and one in the 12M-old group (115 s). Median values of groups ranged from 4 to 9 s (Fig. 3). We observed no significant differences between groups on day 1 (H=4.884; p=0.09).
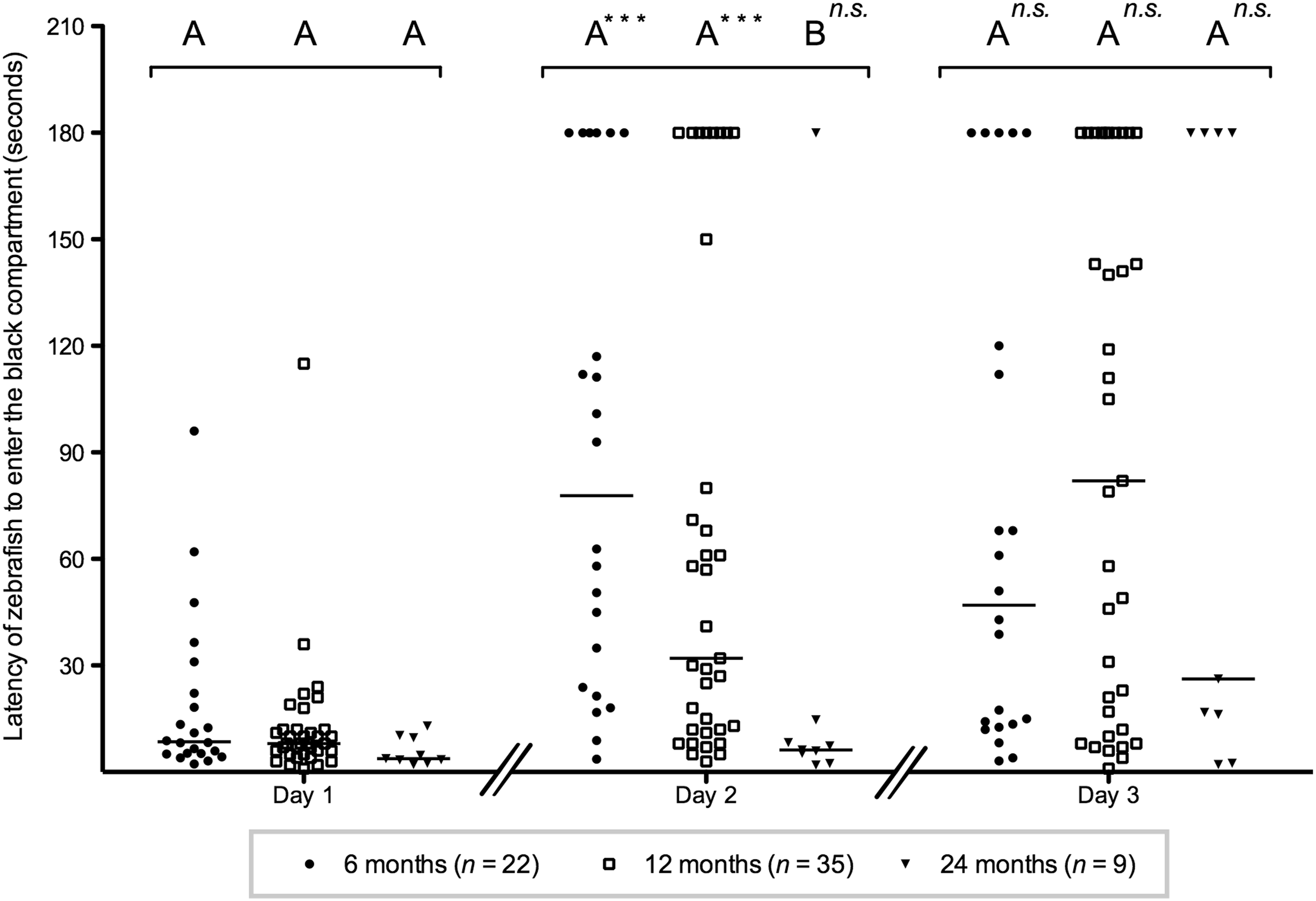
Inhibitory avoidance learning in 6M-, 12M-, and 24M-old zebrafish. Day 1 (no training) shows no significant differences in the initial latencies between 6M-old (black dots), 12M-old (open squares), and 24M-old (black triangles) zebrafish. Day 2 (one shock) shows a significant lower latency for the 24M-old zebrafish compared with the 6M- and 12M-old zebrafish. Day 3 (two shocks) shows no significant differences in latencies between 6M-, 12M-, and 24M-old zebrafish: bars indicate median values. Capital letters above groups indicate significant differences assessed by the Dunn's multiple comparison test following the Kruskal–Wallis test: groups that do not share corresponding letters are significantly different from each other. Triple asterisks (p≤0.001) indicate a significant increase in latency from days 1 to 2 or from days 2 to 3 within a single age category. n.s.=not significant, n=the number of fish.
On day 2, the latencies to enter the dark compartment of the tank of the 24M-old fish were significantly lower compared with the latencies of both the 6M- and 12M-old fish (H=11.96; p=0.003). On day 3, we did not observe significant differences between the three age groups (H=0.4284; p=0.81).
The latencies on days 2 and 3 were significantly higher compared with the latencies on day 1 for both the 6M-old fish (H=18.58; p<0.0001) and the 12M-old fish (H=29.15; p<0.0001), but there was no significant difference between days 2 and 3 for either age. For 24M old fish, we did not observe significant differences across days (H=5.785; p=0.06).
Age: gene expression analysis
Statistics on the data are shown in Figure 4A–M. Following the inhibitory avoidance paradigm, expression levels of bdnf, pcna, and cart4 were significantly lower in 24M-old zebrafish compared with both the 6M- and 12M-old zebrafish.
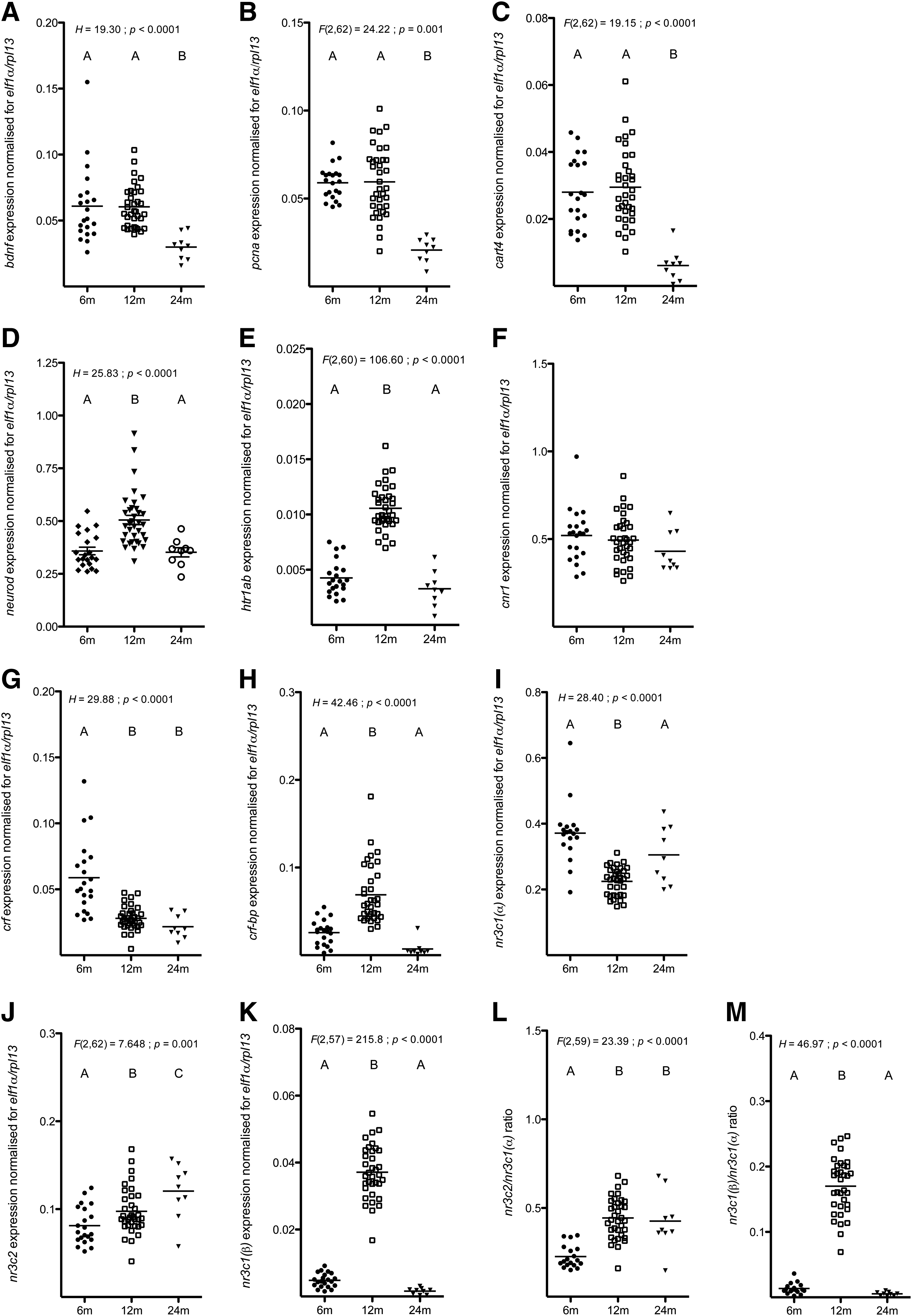
Gene expression levels of neurod, htr1ab, crf-bp, nr3c1(β), and the nr3c1(β)/nr3c1(α) ratio were significantly higher in the 12M-old fish compared with both the 6M- and 24M-old zebrafish, while the expression of nr3c1(α) was significantly lower.
The expression of crf was higher in the 6M-old group compared with both the 12M- and 24M-old zebrafish, whereas the nr3c2/nr3c1(α) ratio was significantly lower.
Only for the expression of nr3c2, we observed significant differences between all age groups, where the expression levels increased with age. We found no significant differences for cnr1 expression.
Experiment 3: age and enrichment
Age and enrichment: inhibitory avoidance behavior: 6M-old zebrafish
All fish entered the black compartment within 60 s on the first day, except three individuals: two in the barren-housed group (62 and 97 s) and one in the enriched-housed group (118 s) (Fig. 5A).

Inhibitory avoidance learning following 4 weeks of enrichment in 6M- and 24M-old zebrafish.
On day 1, median values of both barren-housed and enriched-housed groups were 9 s, and there was no significant difference between the groups (U=165.0; p=0.76). However, we observed significantly higher latencies for barren-housed fish compared with enriched-housed fish on day 2 (U=48.00; p=0.0002) and day 3 (U=90.00; p=0.01).
In addition, we found that latencies for barren-housed fish on days 2 and 3 were significantly higher compared with the latencies on day 1 (H=18.58; p=0.0001), but there was no significant difference between days 2 and 3. For enriched-housed fish, we did not observe significant differences (H=1.088; p=0.58).
Age and enrichment: inhibitory avoidance behavior: 24M-old zebrafish
All fish entered the black compartment within 60 s on the first day (Fig. 5B).
On day 1, median values of groups were 4 and 6 s for barren-housed and enriched-housed fish, respectively, and the Mann–Whitney U test did not reveal significant differences between rearing conditions (U=65.0; p=0.42). We observed no significant differences between the latencies of barren-housed fish and enriched-housed fish on day 2 (U=47.00; p=0.12) and day 3 (U=66.00; p=0.58).
In addition, we found no significant difference in latencies across days for both barren-housed fish (H=5.785; p=0.06) and enriched-housed fish (H=5.689; p=0.06).
Age and enrichment: gene expression analysis
Statistically significant differences in gene expression between barren-housed and enriched-housed fish following inhibitory avoidance were found (Table 2). We observed the following significant differences for the 6M-old fish: compared with barren-housed fish, enriched-housed fish had significantly lower gene expression levels of pcna, neurod, cart4, and cnr1, but the ratios of nr3c2/nr3c1(α) and nr3c1(β)/nr3c1(α) were significantly higher.
Values listed are means±1 SD of the relative and normalized (to elf1α and rpl13) expression for each group. Please note that these data are not presented such that the different age groups can be compared.
M, month.
In contrast, for the 24M-old fish, we only observed a significantly lower nr3c1(β)/nr3c1(α) ratio for enriched-housed fish. No other significant differences were observed.
Discussion
In this study, we report a decrease in anxiety-like behavior and an increase in exploratory behavior following long-term housing under enriched conditions. In addition, exposure to environmental enrichment reduced inhibitory avoidance behavior in 6M- and 12M-old fish, but not in 24M-old fish. Gene expression analysis following the task revealed that enriched-housed 6M-old fish had lower levels of pcna, neurod, cart4, and cnr1 in the telencephalon and higher telencephalic ratios of nr3c2/nr3c1(α) and nr3c1(β)/nr3c1(α). No such changes were observed for enriched-housed 24M-old fish. We further observed a delay in inhibitory avoidance behavior of 24M-old fish compared with 6M- and 12M-old fish, which was associated with lower telencephalic expression levels of bdnf, pcna, and cart4.
Experiment 1: enrichment
In contrast to previous studies,23,47 where environmental enrichment was either offered for a shorter period (<6 months) or the position of items was changed weekly, we here raised zebrafish under enriched conditions starting from the time they were hatched and aimed to keep the position of enrichment items constant and unchanged. Fish housed under these enriched conditions displayed reduced anxiety-like behavior (more time spent in the white compartment) and increased exploration (more transitions between the white and black compartment), which is in line with previous experiments.23,47 Although we did not observe a significant interaction term, our data do suggest that barren-housed fish become more explorative and less anxious toward the end of the experiment (i.e., minutes 5 and 6). Indeed, separate t-tests show a significant difference between enriched- and barren-housed fish for minutes 2, 3, and 4, but not for minutes 5 and 6.
In rats, environmental enrichment reduced inhibitory avoidance learning,8,9 while mice raised in an enriched environment had increased inhibitory avoidance behavior.6,7 In our study, we observed that zebrafish from an enriched environment displayed reduced inhibitory avoidance behavior. Interestingly, not all fish in the enriched-housed group seemed to have lowered avoidance behavior, as we still observed fish that learned to refrain from swimming to the dark compartment where they experienced a shock before. However, the number of such fish in the enriched-housed group was clearly lower compared with the barren-housed group, and this resulted in an overall lower latency time.
Impaired associative learning or a different appraisal of the shock by the subject could be at the basis of this observed reduction in avoidance behavior. However, studies in several fish species have demonstrated that environmental enrichment promotes learning and memory in fish24–27 and suggests that impaired associative learning in all likelihood does not underlie the reduction observed. It is thus more likely that the electric shock had a weaker impact for fish from an enriched environment. In rodents, environmental enrichment attenuated the need for pain relief in animals submitted to invasive procedures 48 and reduced the injury-induced nerve hypersensitivity to both mechanical and cold stimuli, 49 that is, environmental enrichment is accompanied by a potentially higher pain threshold. Thus, an increased pain threshold due to environmental enrichment would offer an explanation for the reduction in avoider fish. It is clear that this warrants further studies (see concluding remarks below).
Experiment 2: age
Inhibitory avoidance behavior in 6M-old zebrafish was not significantly different from that of 12M-old zebrafish in our Kruskal–Wallis test. However, our data suggest that there might be a difference in learning curves between these age groups. And, indeed, when comparing the 6M-old group with the 12M-old group only, we observed no difference on day 2 (p=0.07), but a significant difference on day 3 (p=0.02). Where the 12M-old group showed a steady increase in latency after each shock, the 6M-old group had a strong increase after a single shock but did not improve performance following the second shock. The expression levels of neurod, htr1ab, crf-bp, nr3c1(β), and nr3c2 following the inhibitory avoidance paradigm were found to be significantly lower in 6M-old zebrafish, whereas the expression of crf and nr3c1(α) was significantly higher compared with the 12M-old group. In addition, the nr3c2/nr3c1(α) ratio was found to be significantly lower. As the ratio between mineralocorticoid receptor (MR) and glucocorticoid receptor (GR) activation is crucial for learning and memory,33,50 differences in the ratio of the expression of these genes observed between 6M- and 12M-old fish may have contributed to the observed differences in latency on day 3. The observed higher levels of crf and lower levels of crf-bp mRNA are likely related to age differences: in rats, levels of Crf increased with age while the expression of Crf-bp decreased. 51 Corticotropin-releasing factor (CRF)-family peptides within the brain modulate learning and memory,52–54 and differences in expression levels may cause differences in learning and memory. Although dependent on the region where neurod is expressed, expression levels in the brain change as an individual ages. In the cerebellum of rodents for instance, an increased expression was observed followed by a decline at older age.55,56 The serotonergic system also undergoes changes during development and aging57,58; in rats, serotonin transporters increased up to the adult phase after which they decreased again with aging. 59 Although little is known about the expression of htr1ab specifically during aging, our expression data (showing a hyperbolic relationship) suggest that the expression of htr1ab may follow a similar pattern.
Although 6M- and 12M-old zebrafish have been classified as “adults,” 31 our data suggest that differences between 6M- and 12M-old zebrafish exist in avoidance learning. We observed minor, but significant, differences in inhibitory avoidance performance, and telencephalic gene expression showed strong differences following the task, most of which are linked to the endocrine stress system. Further characterization of the maturation of the zebrafish brain seems warranted.
Twenty-four-month-old zebrafish appear to have delayed inhibitory avoidance behavior compared with 12M-old zebrafish, in the sense that they required an extra training day (day 3) to reach a similar latency time as 12M-old zebrafish (day 2). This reduced avoidance behavior might relate to the vast reduction in telencephalic gene expression related to plasticity and learning observed. Compared with 12M-old zebrafish, the 24M-old group had reduced levels of bdnf, pcna, cart4, neurod, htr1ab, and crf-bp and a higher expression of nr3c1(α) and nr3c2 in the telencephalon. The overall reduced expression of genes suggests a lower brain activity and neural plasticity. Lower expression of bdnf is associated with aging 58 and may lead to reduced fear learning and lower ability to consolidate fear memories.60,61 Higher expression of pcna has been directly linked to increased cell proliferation in the brain of fish62,63 and may be considered as an indicator for brain plasticity. Higher concentrations of cocaine and amphetamine-regulated transcript (CART) peptide lead to enhanced learning and memory39,64 and a reduction in the expression of cart4 likely results in reduced peptide concentrations. In line with these observations, age-related spatial changes in the expression of neurod have been observed in the rodent brain55,56 and similarly may hold true for the fish brain. As discussed earlier, the serotonergic system changes during aging and the lower expression of htr1ab fits in the overall reduction of the system as seen at later age.57–59 In contrast to the reported increase of Crf-bp expression in aged rats, 51 we observed a reduction in the crf-bp expression in older zebrafish. This reduction might result from an interaction of the aging effect with the effect of our inhibitory avoidance task, which lowers crf-bp expression (unpublished data). CRF peptides modulate learning and memory formation52–54 and are bound by corticotropin-releasing factor-binding protein (CRF-BP). By doing so, CRF-BP inhibits CRF-mediated activation of CRF receptors. 65 A reduction in crf-bp expression may thus result in more CRF-mediated activity in the brain. Higher expression of nr3c2 may indicate a higher need for MR activation; for under basal conditions, the MR is fully occupied to maintain basal neuronal activity. 66 Both MR and GR are involved in learning and memory,67–70 and the ratio between MR and GR activation is crucial.33,50 This may explain the higher nr3c1(α) expression, as it may maintain a proper MR/GRα ratio. Indeed, the nr3c2/nr3c1(α) ratio does not differ between 12M- and 24M-old fish. The need for more GRα activation is also reflected by the decrease in nr3c1(β), which is a splice variant that gives rise to GRβ. In zebrafish, the GRβ is thought to act as a decoy receptor, which inhibits the activation of GRα (but not MR due to its much higher affinity for steroids) by binding circulating glucocorticoids. 71 A reduction in GRβ seems thus an adaptive mechanism to ensure activation of GRα.
Experiment 3: age and enrichment
Similar to the effects observed in our 12M-old zebrafish, inhibitory avoidance behavior in 6M-old zebrafish housed in an enriched environment was reduced. Gene expression analysis following inhibitory avoidance revealed a lower expression level of pcna, neurod, cart4, and cnr1 in 6M-old fish from an enriched environment. These observations are in contrast with previous studies, where pcna and neurod were upregulated following housing under enriched conditions.21,72 Yet, the decrease in pcna, neurod and cnr1 is in line with a decrease in inhibitory avoidance as indicated above and as revealed by comparison with unpublished baseline data from our laboratory. Thus, the higher expression of these genes in the barren-housed group may reflect the acquisition of avoidance behavior, which was not observed in the enriched group. The lower expression of cart4 found in the enriched group may also be related to avoidance learning rather than housing condition. While gene expression levels of cart4 are decreased when measured after the task (Manuel et al. 4 ), we reported previously that cart4 expression was higher in avoider fish (latencies equal to or larger than 180 s) than in nonavoider fish (latencies smaller than 60 s) when a 3 V shock was given. 4 Our enriched-housed group consisted nearly only of nonavoider fish, which corresponds to lower cart4 expression, whereas the barren-housed group contained both nonavoider and avoider fish, which corresponds to higher levels of cart4. CART is involved in learning and memory,39,64 but higher levels of CART are also known to increase fear and anxiety-like behavior.39,73 The lower expression of cart4 in the enriched-housed fish not only relates to the observed behavior in our current study: reduced anxiety-like behavior (6 months) and reduced inhibitory avoidance (12 months) but also corroborates previously reported inhibitory avoidance behavior and associated cart4 expression. 4 Thus, next to possible changes in pain as discussed above, enrichment may affect levels of fear, lowering inhibitory avoidance behavior.
In 24M-old zebrafish, we saw no clear effect of enrichment with regard to inhibitory avoidance behavior or gene expression analysis. We only observed a reduction in the nr3c1(β)/nr3c1(α) ratio, which suggests a higher potential for cortisol to activate GRα. Possibly, 4 weeks of enrichment is too short a period, or the enrichment came too late in the life history for 24M-old fish to benefit. In rats, lifelong enrichment reduced cognitive dysfunction at an older age. 43 Whether lifelong enrichment has similar effects on zebrafish warrants further studies. However, we would suggest to expand the behavioral assessment paradigms to include a spatial learning task, as performance in such a task improved with enrichment. 72 Here, both environmental enrichment and old age reduced inhibitory avoidance behavior, making it difficult to properly assess the effects of environmental enrichment on the cognitive abilities of aged fish.
Limitations
It should be noted that anxiety-like behavior could also have been assessed using the novel tank test 74 rather than the black and white preference test. While both tests may be used to assess anxiety-like behavior, data suggest that they may not assess the same phenomenon as pharmacological profiles are not always identical. 74 Therefore, our conclusions only relate to anxiety-like behavior (and the modulation thereof) as assessed through a black and white preference test. In addition, as we tested animals individually, this may have an effect on the outcome of the tests. 75 This also applies to our inhibitory avoidance protocol. This warrants further studies.
Given that we sampled post-task, differences in gene expression between experimental groups are closely associated with differences in inhibitory avoidance behavior, as discussed earlier. 5 We do not as yet know whether differences in telencephalic gene expression between age and/or enrichment conditions were already present before the task and how the task alters them. However, data from our earlier experiment in barren-housed 6M-old zebrafish 4 using a 1-day shock paradigm showed only little differences between pre-task (baseline) and post-task gene expression levels. As indicated above, preliminary data comparing baseline and task-related gene expression levels in 12M-old barren-housed fish revealed changes in levels of gene expression, which may help explaining our findings. We are currently studying this in more detail.
Concluding Remarks
Our data show that environmental enrichment reduces inhibitory avoidance behavior in 6M- and 12M-old zebrafish. For 6M-old zebrafish, this enrichment-induced reduction in performance was associated by differences in the mRNA levels of several genes in the telencephalon indicative for neuroplasticity. In contrast, 24M-old fish showed poor/delayed inhibitory avoidance behavior without any noticeable effects of environmental enrichment on task performance. This poor task performance was associated with strongly reduced levels of genes indicative for neuroplasticity as well as the endocrine stress system. Where we observed minor differences in avoidance behavior between 6M- and 12M-old zebrafish, post-task gene expression analysis revealed clear differences in the expression level of genes involved in the endocrine stress system. For example, both the 6M and 12M age groups are often categorized as “adult” based on external morphological characteristics, yet our study indicates that there are age-related differences in behavior, cognition, and gene expression profiles.
Footnotes
Acknowledgments
The authors thank Mr. F.A. Tom Spanings for excellent organization of fish husbandry and expert technical assistance throughout the experiment. This study was sponsored by the Dutch Science Organization (NWO; programme: Value of Animal Welfare; project no. 827.09.040). Further support came from the Norwegian Research Council (grant no. 190469) (L.O.E.E.) and by the EU Seventh Framework Programme (FP7/2010–2014) under grant agreement no. 265957–COPEWELL (L.O.E.E., M.G., and G.F.)
Disclosure Statement
No competing financial interests exist.