
Abstract
Abstract
Astyanax taeniatus occurs in coastal areas of southeastern Brazil, and it is very abundant in the Upper Doce River Basin. Our objective was to study C-, argyrophilic nucleolar organizer region (Ag-NOR) and fluorescent in situ hybridization (FISH) banding patterns using 5S, 18S, CA(15), and GA(15) repetitive DNA probes on a population of A. taeniatus present in the Piranga River, in the Doce Basin. Two syntopic cytotypes were found, both with 2n = 50: cytotype A (14m + 12sm + 16st + 8t) and cytotype B (10m + 14sm + 18st + 8t). In both cytotypes, heterochromatic blocks occurred in all the chromosomes; Ag-NOR sites were multiple, ranging from four to eight. The 5S rDNA probe marked eight chromosomes in both cytotypes, a unique condition within Astyanax, suggesting a recent divergence between these cytotypes. The 18S rDNA probe differed between the cytotypes, marking 10 and 8 chromosomes in cytotypes A and B, respectively. CA(15) and GA(15) FISH patterns were mainly subtelomeric, but CA(15) showed centromeric markings that were diagnostic for each cytotype. Although overall cytogenetic evidence suggests that these cytotypes are closely related, morphological and molecular data in progress will provide further hypothesis test on their phylogenetic relationship.
Introduction
I
With 154 valid species, Astyanax is the most species-rich genus in the order Characiformes. Most of them are widely distributed in the Neotropical region from southern United States to northern Argentina. 7 Cytogenetic research shows high levels of variation in this genus, and it also suggests that some nominal species such as Astyanax altiparanae Garutti & Britski, 2000,8,9 Astyanax fasciatus (Cuvier, 1819),10,11 Astyanax scabripinnis (Jenyns, 1842),12,13 and Astyanax hastatus Myers, 1928 14 are actually species complex, and several good species have been validated within these complexes. Astyanax species are morphologically similar, and their taxonomic classification has been historically difficult; the last revision in this genus was made almost a hundred years ago.15–18 Moreover, a complicating factor for understanding the group's systematics is the occurrence of hybridization in some of its taxons.10,11,19
Astyanax taeniatus was described in 1842 by Jenyns from the type locality Socego, Rio de Janeiro State. This species also occurs in the Paraíba do Sul River Basin, in the drainage of coastal rivers of Rio de Janeiro and Espírito Santo, 2 in the Doce River Basin, 20 and in the São Mateus and Mucuri river basins. 21 This species is the most abundant characin present during the annual reproductive migration in the Risoleta Neves Dam, in the Upper Doce River. Although the diploid number and chromosomal formula have been characterized from specimens of A. taeniatus belonging to the isolated coastal basins such as Jucu and Benevente, 22 the cytogenetic banding and fluorescent in situ hybridization (FISH) patterns from Upper Doce River Basin population are still unknown.
Karyotypic comparisons between species, associated with taxonomic and molecular biological studies, provide useful information for studying evolution, phylogeny, and understanding of mechanisms of speciation. 23 In this study, we aimed to analyze a population of A. taeniatus (Jenyns, 1842) present in the Piranga River, using various cytogenetic tools. The study area is located in the Doce River Basin. Like many other Neotropical drainages, the Doce River Basin is small and isolated and is characterized by low levels of species richness and high levels of endemism. 24
Materials and Methods
Specimen collection
The specimens of A. taeniatus were collected in the Hydroelectric Risoleta Neves Reservoir fish scale (20°11′41.92"S 42°51′04.95"W), in the Piranga River, Upper Doce River Basin, located in the municipalities of Rio Doce and Santa Cruz do Escalvado, Minas Gerais, Brazil. Collection permit of the Instituto Chico Mendes de Biodiversidade (ICMBio) (SISBIO14975-1) was issued to Jorge Abdala Dergam. Species identification followed Eigenmann. 17 Voucher specimens were deposited in the scientific collections of the Museu de Zoologia João Moojen in Viçosa, Minas Gerais, Brazil (voucher MZUFV3992) and the Universidade Federal do Rio Grande do Sul, Rio Grande do Sul, Brazil.
Karyological analyses
The specimens of A. taeniatus were anesthetized with clove oil 25 as approved by the Universidade Federal de Viçosa Animal Welfare Committee (CEUA authorization 58/2013). Mitotic chromosomes were obtained from kidney following Bertollo et al. 26 The following cytogenetic techniques were used: Giemsa conventional staining for basic karyotypic analysis; C-banding 27 stained with fluorescent 4′,6-diamidino-2-phenylindole (DAPI) dye 28 to identify blocks of constitutive heterochromatin; argyrophilic nucleolar organizer region (Ag-NOR) banding for their identification 29 ; and FISH 30 using 5S and 18S rDNA probes and CA(15) and GA(15) repetitive DNA probes.
Ribosomal DNA probes 18S and 5S were labeled using PCR with digoxigenin-11-dUTP (Roche Applied Science) and the signal was detected with anti-digoxigenin-rhodamine (Roche Applied Science). DNA was denatured with 70%/2× SSC formamide solution at 72°C for 3 min; each slide was treated with 20 μL of hybridization mix containing the following: 200 ng of labeled probe, 50% formamide, 2× SSC, and 10% dextrane sulfate. This hybridization mix was heated for 10 min at 85°C; slides were kept in a moist chamber at 37°C overnight; and after the hybridization step, the slides were washed twice in 2× SSC for 5 min and incubated in a 3% NFDM 4× SSC solution for 10 min and placed in a moist chamber at 37°C for 1 h with a detection solution using anti-digoxigenin-rhodamine (Roche Applied Science).
Finally, the slides were washed thrice in a 4× SSC/0.5% Tween solution for 5 min at room temperature; they were dehydrated in an alcoholic series and counterstained with DAPI. As for the probes CA(15) and GA(15), they were synthesized and labeled with Cy3 fluorochrome at the 5′ end (Sigma). Digital images were captured in a BX53F Olympus microscope equipped with DP73 and MX10 Olympus camera for colored and fluorescent techniques, respectively, using the CellSens imaging software. The karyotypes were measured with the Image-Pro Plus® software and classified according to arm ratios as metacentric (m), submetacentric (sm), subtelocentric (st), and telocentric (t). 31
Results
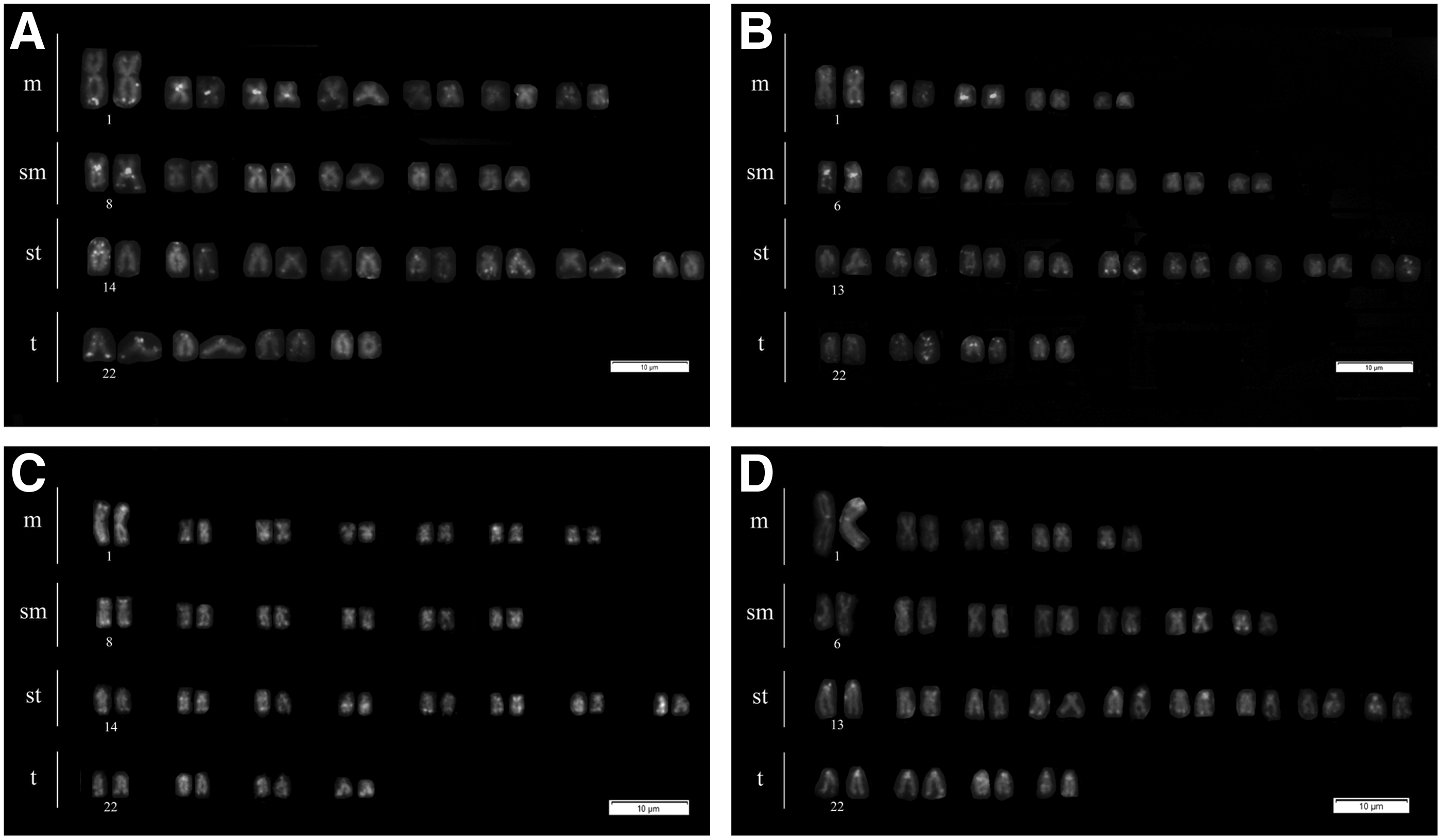
A total of 77 individuals were cytogenetically analyzed, of which, 28 were males, 44 were females, and 5 were juveniles. The diploid chromosome number was 2n = 50, without sex chromosomes. Two karyotypes were found: 30 specimens showed cytotype A: 2n = 14m + 12sm + 16st + 8t (Fig. 1A), whereas 47 specimens showed cytotype B: 2n = 10m + 14sm + 18st + 8t (Fig. 1B).

Giemsa-stained karyotype of Astyanax taeniatus:
In both cytotypes, all chromosomes showed centromeric heterochromatin blocks with additional pericentromeric marks in most submetacentric and subtelocentric chromosomes (Fig. 1C, D). The Ag-NOR banding showed multiple marks in both cytotypes varying within and among individuals from four to eight subtelocentric chromosomes located on their short arms; ranging from chromosome pairs 14 to 17 in cytotype A (Fig. 1E) and chromosome pairs 13 to 16 in cytotype B (Fig. 1F).
The 5S rDNA probe marked four chromosome pairs 19, 20, 21, and 25 in both cytotypes (Fig. 1G, H green probe), while the 18S rDNA probe marked five chromosome pairs 9, 14, 15, 16, and 17 in cytotype A, chromosome pair number nine lacked Ag-NOR markings, and only four chromosome pairs 13, 14, 15, and 16 in cytotype B (Fig. 1G, H red probe). Repetitive FISH patterns obtained with CA(15) varied between both cytotypes: cytotype A had three pairs of chromosomes with strong centromeric marks (two metacentric pairs and one submetacentric pair), while cytotype B had only two pairs with strong centromeric marks (one metacentric pair and one submetacentric pair) (Fig. 2A, B). Repetitive FISH patterns obtained with GA(15) probe marked the subtelomeric regions of all chromosomes in both cytotypes (Fig. 2C, D).
FISH patterns obtained with repetitive DNA probes in A. taeniatus:
Discussion
A. taeniatus has 50 chromosomes as well as most species of the genus Astyanax; a ploidy considered a primitive character state within the family Characidae. 32 This is the first report of two distinct syntopic karyotypes in this species. These cytotypes differed from the Jucu and Benevente rivers population, with its karyotypic formula = 12m + 2sm + 24st + 12t. 22 The Piranga River cytotypes also differed from each other by non-Robertsonian chromosome rearrangements, suggesting that chromosomal divergence involved pericentromeric inversions that changed the number of chromosomes assigned to each morphological class.8,33
The C- and Ag-NOR bandings showed similar results in both cytotypes suggesting a recent karyotypic divergence between these lineages. Furthermore, all chromosomes showed conspicuous heterochromatic blocks that were apparently more intense than the ones observed in other Astyanax species9,34–36 and provide further evidence of close phylogenetic relationships between cytotypes A and B. High level of intraindividual and intrapopulational variation of multiple Ag-NORs are frequently observed in the genus Astyanax. 37 Up to 10 Ag-NOR-bearing chromosomes have been reported in A. altiparanae and A. fasciatus species complex, with a maximum number of 15 in the A. scabripinnis complex (reviewed in Abelini et al. 38 ). In other Astyanax species, Ag-NORs cistrons occur in all chromosome morphological classes except telocentrics and are located on either the short or long chromosome's arms.38,39
The 5S rDNA probe showed a conserved pattern in both cytotypes. Within the genus Astyanax, this feature can be considered a derived condition since all species already studied with this probe showed fewer markers than the ones observed in this study (revised in Kavalco et al. 9 ), further suggesting a common origin of both cytotypes. The 18S rDNA probe also showed a large number of sites. When compared with the Ag-NOR banding, it suggests that most ribosomal sites are active in these cytotypes. Moreover, other species of the genus Astyanax also show small discrepancies between the number of Ag-NOR and 18S rDNA sites.40–43
Our study is the first that used CA(15) and GA(15) repetitive DNA probes to characterize an Astyanax species. In other species, such as Hoplias malabaricus 44 and Mystus bocourti, 45 these probes hybridized on subtelomeric chromosome regions. In Eremias velox, 46 Leporinus species, 47 and Triportheus trifurcatus, 48 these probes showed additional interstitial marks in their sex chromosomes. In this study, the CA(15) and GA(15) probes were mainly subtelomeric, but the CA(15) pattern included additional centromeric markings that were specific for each cytotype.
Our results indicate that the species A. taeniatus collected in the Upper Doce River exhibits two syntopic cytotypes. If ongoing molecular data confirm the specific status of specimens bearing different karyotypes, it would be interesting to understand the reproductive isolation mechanism between these cytotypes. In A. scabripinnis, large water bodies apparently act as physical barriers for the species. This species is restricted to headwaters or small springs, which hinders sympatry of different lineages, precluding hybridization between them.12,13 However, A. taeniatus occurs in the river channel and is a very active and opportunistic species with regard to habitat and feeding aspects, 49 suggesting reproductive barriers that may be supported by some behavioral/genetic mechanisms.
In the A. fasciatus species complex, karyotypic variations have been interpreted as evidence for the existence of viable hybrids that withstand a set of chromosomal combinations,10,11 while this condition may not apply to other sympatric populations of this species complex.35,36,50 Another widely studied species complex is A. scabripinnis that shows high levels of karyotypic diversity among allopatric 51 and sympatric 52 populations; in this case, however, no intermediate hybrid forms have ever been found.12,13
Although sympatric speciation cannot be ruled out, its occurrence is difficult to test.53,54 Within the Doce River Basin, vicariance between the river bed and a quaternary lacustrine system may have caused speciation between the small predator species Oligosarcus argenteus and Oligosarcus solitarius (Dergam, unpublished) and has also caused deep genetic differentiation within Geophagus brasiliensis populations. 55 Even if the two cytotypes of A. taeniatus are proven sister species, they may have evolved separately by allopatric speciation and became sympatric as a secondary contact.
Footnotes
Acknowledgments
The authors thank “Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq),” “Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)” and “Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG)” for their financial support.
Disclosure Statement
No competing financial interests exist.