
Abstract
Abstract
Bisphenol A (BPA) acts as an endocrine-disrupting compound even at a low concentration. Degradation of BPA could lead to the formation of toxic products. In this study, we compare the toxicity of BPA and seven intermediate products of its degradation. The accuracy of three molecular docking programs (Surflex, Autodock, and Autodock Vina) in predicting the binding affinities of selected compounds to human (ERα, ERβ, and ERRγ) and zebrafish (ERα, ERRγA, and ERRγB) estrogen and estrogen-related receptors was evaluated. The docking experiments showed that 4-isopropylphenol could have similar toxicity to that of BPA due to its high affinity to ERRγ and ERRγB and high octanol–water partitioning coefficient. The least toxic compounds were hydroquinone and phenol. Those compounds as well as BPA were screened in the zebrafish (Danio rerio) embryo test. 4-isopropylphenol had the strongest toxic effect on zebrafish embryos and caused 100% lethality shortly after exposure. BPA caused the delay in development, multiple deformations, and low heartbeats (30 bps), whereas hydroquinone had no impact on the development of the zebrafish embryo. Thus, the results of zebrafish screening are in good agreement with our docking experiment. The molecular docking could be used to screen the toxicity of other xenoestrogens and their products of degradation.
Introduction
B
BPA is known to be an endocrine- disrupting compound, that is, it shows an estrogenic activity even though it is not structurally related to estrogen. Multiple studies report that the ng/L to mg/L BPA concentration is toxic for reproductive health, obesity, and early development. 6 However, the toxicity and estrogenicity of BPA degradation products have not been studied widely enough to make sure that the degradation leads to less toxic products.
The mammal tests are usually used to evaluate toxicity, however, they are very time-consuming and expensive. As a good alternative, zebrafish is used to screen the endocrine-disrupting properties and toxicity of various natural and synthetic compounds. 7 Zebrafish becomes a good model organism due to fast development of the embryo, transparent eggs, and good standardization of each stage of the embryo development. 8 The toxicological effects observed in the zebrafish embryo development test could be approximated to those of higher vertebrates, 9 however, some assumption should be made about a similar mechanism of action. It was shown that the zebrafish embryo development test predicts well the toxic effect of model toxic compound, whereas prediction of the results for less toxic compounds could be of about 60%–70%.10,11
The effects of BPA9,12 and its intermediate products of degradation such as phenol, hydroquinone, 13 and 4-isopropylphenol 14 were screened with the zebrafish embryo toxicity test. However, some studies report only the mortality rate or concentrate on several features of the development. That makes the comparison between the compounds hard or even impossible.
Some attempts were made to evaluate the toxicity of BPA and compare it with other compounds. For example, the octanol–water partitioning coefficient was suggested as a toxicity marker. 9 Molecular docking of the compounds to appropriate receptors could be a complementary approach to the octanol–water coefficient. In the case of BPA and its degradation product molecular docking to its main target receptor, that is, human estrogen-related receptor gamma (ERRγ) or zebrafish ERRγB, 15 allows one to compare the estrogenicity of degradation products with the parent compound.
In this work, we performed the toxicity screening of BPA and its seven intermediate products of degradation, namely hydroquinone, 4-isopropylphenol, 4-(2-hydroxy)isopropylphenol, 1,4-benzoquinone, 3-methylbenzo[b]tetrahydrofuran, 2-methylbenzo[b]tetrahydrofuran, and phenol. The aim of the study was to identify (i) whether or not the BPA degradation leads to less toxic and less estrogenic compounds; and (ii) if the products of degradation show similar mechanism of toxicity to that of BPA. In this work, we calculated the octanol–water coefficient, as well as performed molecular docking of the BPA and selected products of its degradation, 17β-estradiol and 4-hydroxytamoxifen to human receptors ERα, ERβ, and ERRγ and zebrafish (Danio rerio) ERα, ERRγA, and ERRγB. We also did the zebrafish embryo development toxicity test. Our results suggest that 4-isopropylphenol has a similar mechanism of toxicity with BPA. However, further investigations of 4-isopropylphenol toxicity are required. Moreover, our results suggest that 4-(2-hydroxy)isopropylphenol, 3-methylbenzo[b]tetrahydrofuran, and 2-methylbenzo[b]tetrahydrofuran could also reveal some similar mechanisms of toxicity that are similar to BPA and should be screened with the zebrafish embryo test. Such an approach could be used for screening other products of BPA degradation that could occur when other AOPs are used, for example ultrasound-assisted degradation.
Material and Methods
Chemicals
BPA, phenol, and 4-isopropylphenol were purchased from Sigma-Aldrich. Hydroquinone, calcium chloride, magnesium sulfate, sodium bicarbonate, and potassium chloride were supplied by Avantor Performance Materials Poland S.A. All reagents were of analytical grade and were used as received without any further purification. Test solutions were freshly prepared for each independent experiment by dissolving BPA, phenol, 4-isopropylphenol, and hydroquinone in the REKO buffer (0.294 g CaCl2, 0.123 g MgSO4, 0.065 g NaHCO3, and 0.006 g KCl, per liter at pH 7.0, in ultrapure water). The concentrations of the studied compounds were as follows: BPA 20 mg/L, phenol 20 mg/L, 4-isopropylphenol 5, 10, and 20 mg/L, and hydroquinone 20 and 40 mg/L.
Fish care
Wild-type adult zebrafish (D. rerio) line ABxTL were kept in system water (pH 7.0, 600 μS) at a temperature of 28°C with maximal density up to five fish per liter of water. The 14-h day/10-h night regime was used. The fish were fed four times a day with dry feeds or artemia. The embryos were obtained in natural crosses. The eggs were rinsed with REKO to remove debris, sorted, and kept on a Petri dish at 28.5°C in an incubator with the light cycle set at 14/10 and humidity 60%.
The zebrafish embryo toxicity test
The zebrafish embryo toxicity test was performed according to published guidelines. 16 For each experiment, 20 fertilized eggs at the beginning of epiboly stage (4.7 hours postfertilization [hpf]) were used for subsequent experiments. The selected eggs were exposed in 6 mL of BPA, phenol, hydroquinone, and 4-isopropylphenol dissolved in REKO or in REKO for controls. All experiments were run in duplicates. Control in REKO was performed with each set of experiments. The samples were incubated at 28.5°C for 48 h exposure time. Embryonic development was observed with a Leica M165 microscope at 24 and 48 hpf posttreatment. No chorions were removed during the experiment. The observations were made at room temperature (20°C). The somites, pericardial edema, hematoma, heartbeats, blood circulation, pigmentation, yolk and yolk extension deformities, delay in development, and otolith malformation were evaluated. The heart rate was measured only for nonhatched embryos. No anesthetic agent was used. To measure the heart rate, individual embryos were placed under the microscope, and then a 10 s lapse video was recorded. The number of heartbeats was recalculated to the beats per minute. Delay in the development was evaluated based on the percent of embryos not developed to the stage as one in REKO. The somites, pericardial edema, hematoma, blood circulation, significant lack of pigmentation, yolk and yolk extension deformities, and otolith malformation were evaluated visually. The four lethal endpoints, that is, mortality of the embryo, nondetachment of the tail, lack of somite formation, and lack of heartbeat, were used.
Statistics
Statistical analyses were performed using StatSoft, Inc. (2014). STATISTICA (data analysis software system), version 12. One-way ANOVA (Kruskal–Wallis) with post hoc test was employed for heart frequency (beats per minute). To characterize the considered parameter, the expected value (mean), standard deviation, and relative standard deviation were computed. Toxic effects on the parameters of embryo development in control (REKO) and experimental groups were expressed as percentage values, and the statistically significant difference between groups was checked using the directional (one-tailed) test for proportions and the chi-square test. All computations were applied at significance level 0.05.
Molecular docking
Human estrogen receptor α (ERα), estrogen receptor β (ERβ), estrogen-related receptor γ (ERRγ), and zebrafish ERα, ERRγA, and ERRγB were selected for molecular docking. The structures of human receptors were taken from RCSB Protein Data Bank (PDB) (ERα id: 1XPC, ERβ id: 1QKM, ERRγ id: 2E2R). The zebrafish receptor was modeled using SWISS-MODEL. 17 The zebrafish ERα, ERRγA, and ERRγB protein sequences were retrieved from National Center for Biotechnology Information (respectively accession no. AAK16740, 569 amino acid residues; accession no. 998119, 435 amino acid residues; accession no. 001122150, 420 amino acid residues).
The structure of human ERα (PDB id: 2YJA, resolution 1.82Å) was selected as a template. The ligands set consisted of 10 compounds, including BPA, 7 products of its degradation, 17β-estradiol, and 4-OH-tamoxifen. The structures were drawn in Sybyl-X 1.2 Sketch, then hydrogens were added, and finally the structures were optimized (Tripos force field, gradient 0.05 kcal/mol*A). Docking was carried out using Surflex-Dock interfaced with Sybyl-X 1.2 program (Tripos International), AutoDock 4, and AutoDock Vina.18,19 Surflex protomol (an idealized active site ligand) was defined based on the ligand position in the crystal structure from PDB. The AutoDock grid box was centered in the same active site with the size set at 60 × 60 × 60 Å (x, y and z) and spacing between grid points was 0.269 Å. The other parameters of Surflex and AutoDock were used as default values. The best docking pose was chosen according to total scores. The LogP (octanol–water coefficient) values were calculated in HyperChem 7.5 for each ligand.
Results
Hydroquinone, 4-isopropylphenol, 4-(2-hydroxy)isopropylphenol, 1,4-benzoquinone, 3-methylbenzo[b]tetrahydrofuran, 2-methylbenzo[b]tetrahydrofuran, and phenol were proposed as the intermediate degradation products of BPA under photo-Fenton reaction (Fig. 1). 5 Even though the degradation of BPA in this reaction goes fast (9 min for 10 mg/L of BPA), the intermediate products could be present up to 36 h before final degradation to CO2. Thus, during the BPA degradation process the formed intermediate products could influence the environment.

Chemical structure of bisphenol A
Molecular docking
In this work, we screened the estrogenic potential of six products of BPA degradation and compared it with BPA, 17β-estradiol, and 4-OH-tamoxifen. As a target, we selected human estrogen receptor alpha (ERα), estrogen receptor beta (ERβ), ERRγ, and zebrafish (D. rerio) ERα, ERRγA, and ERRγB. The scoring function, which reflects the binding affinity, was calculated using Surflex, Autodock, and Autodock Vina programs (Tables 1 and 2).
The highest affinity of the compound to selected receptor is highlighted in bold (Surflex–maximum positive value, Autodock, Vina–the lowest values), the weakest affinity is in italic.
The highest affinity of the compound to selected receptor is highlighted in bold (Surflex–maximum positive value, Autodock, Vina–the lowest values), the weakest affinity is in italic.
Generally, all products of BPA degradation were found to have lower binding affinity to human ERα, ERβ, and zebrafish ERα than BPA and 17β-estradiol (Table 1). The compound with the second highest binding affinity was 4-isopropylphenol, whereas the lowest binding affinity was found for 1,4-benzoquinone, phenol, and hydroquinone. Only in the case of ERβ, Surflex predicted a higher score for 3-methylbenzo[b]tetrahydrofuran compared with BPA.
In case of estrogen-related receptors, that is, ERRγ and zebrafish ERRγA, 4-isopropylphenol has higher binding affinity than BPA with human receptor (Surflex), and slightly lower than BPA with zebrafish ERRγA (Table 2) (all programs).
In the case of ERRγB, Surflex predicted high hydroquinone binding affinity with ERRγB, which was not reflected in Autodock and Autodock Vina. Also, in Surflex, 3-methylbenzo[b]tetrahydrofuran and phenol had a high score function, whereas the binding affinities determined by Autodock and Vina for those compounds were the lowest.
As the score function values from different programs cannot be compared directly, we inspected the correlation of the studied compounds' ranks between human and the corresponding zebrafish receptor. Good correlations were found for the ERα and zebrafish ERα (0.96 Surflex, 0.96 Autodock, and 0.87 Autodock Vina), as well as human ERRγ and zebrafish ERRγB (0.9 Surflex, 0.91 Autodock, and 0.9 Autodock Vina).
Molecular docking experiments also provided information about pose of the molecule and hydrogen bonds formed. In the case of human ERRγ, 4-OH-Tamoxifen (reference compound binding to human ERRγ) was docked to the pocket formed by Phe435 and Phe450 and it formed hydrogen bonds with Arg316 and Glu275 (Fig. 2). Similar space (Phe435 and Phe450 pocket) was occupied by two methyl groups of BPA and 4-isopropylphenol. For those two compounds, we also observed the interaction with Asn346. Hydroquinone, 4-(2-hydroxy)isopropylphenol, 1,4-benzoquinone, 3-methylbenzo[b]tetrahydrofuran, and 2-methylbenzo[b]tetrahydrofuran were docked in the same cavity limited by Phe435, Phe450, and Tyr326. The hydrogen bonds were formed only by 4-(2-hydroxy)isopropylphenol (Tyr326 and Asn346), hydroquinone (Arg316 and Glu275), and phenol (Glu275).
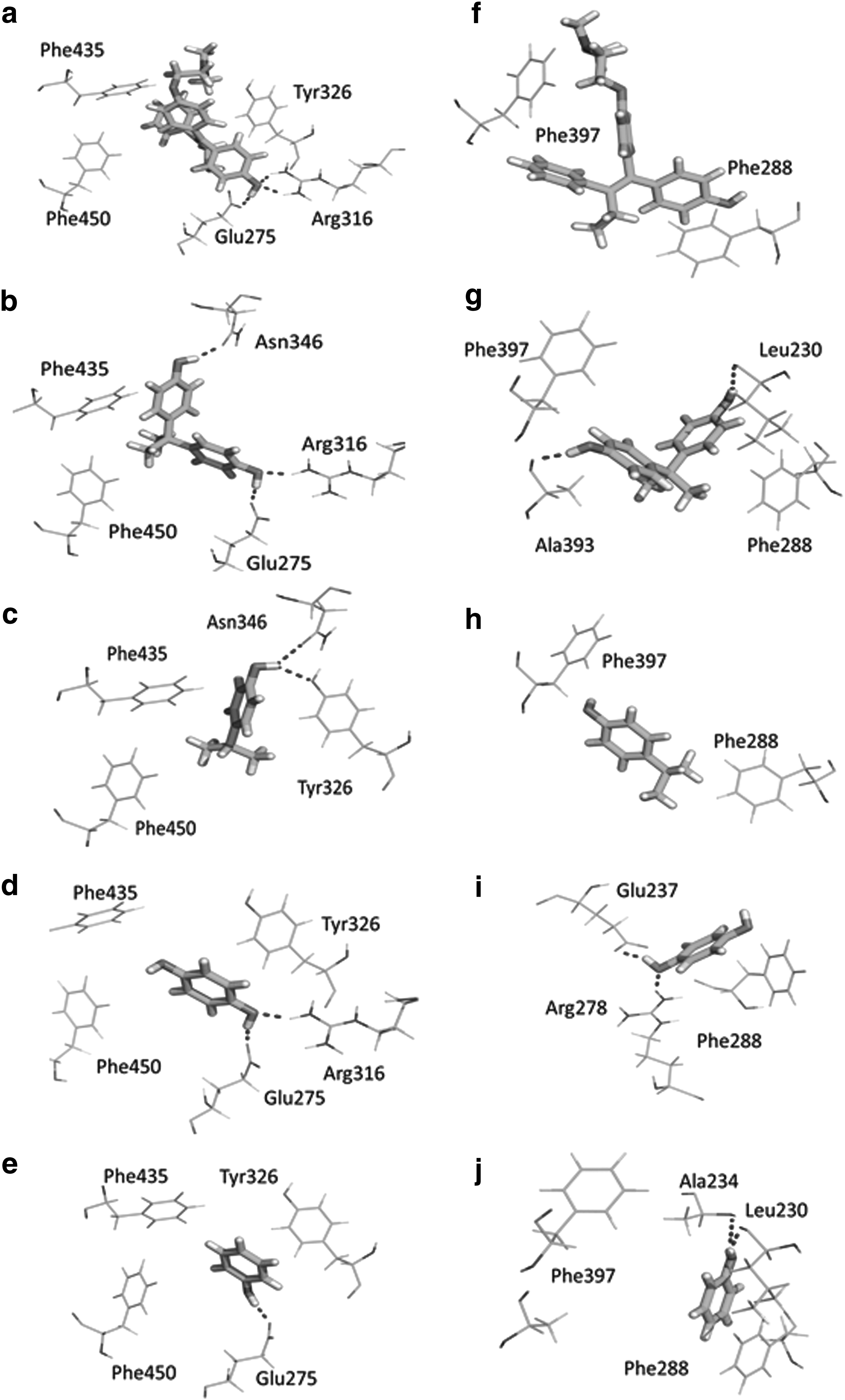
Binding modes of 4-OH tamoxifen
In zebrafish ERRγB, the phenol moiety of 4-OH Tamoxifen occupied the same space formed by Phe288 and Phe397 (corresponding to Phe435 and Phe450 in human ERRγ) and did not form hydrogen bonds (Fig. 2). In contrast to human ERRγ, BPA phenol groups formed only two hydrogen bonds, that is, one with Ala393 (corresponding to Glu275 in human ERRγ) and the other with Leu230 (corresponding to Asn346 in human ERRγ). The rest of the compounds occupied the space defined by Phe288 and Phe397. Similar to human ERRγ, hydrogen bonds were observed for 4-(2-hydroxy)isopropylphenol (Glu237), hydroquinone (Glu237, Arg278), and phenol (Leu230 and Ala234).
Octanol–water partitioning coefficients
Octanol–water partitioning coefficients reflect the ability of the compound to permeate through the biological membrane and combine with the target receptor, thus they are used as another parameter explaining the toxicity of the compound. 9 The highest ability to permeate the membrane was calculated for BPA (4.32), followed by 4-isoprophylphenol (2.96), 2-methylbenzo[b]tetrahydrofuran (2.19), and 3-methylbenzo[b]tetrahydrofuran (2.18). Among the less permeable compounds were phenol (1.76), 4-(2-hydroxy)isopropylphenol (1.72), 1,4-benzoquinone (1.52), and hydroquinone (1.48).
Based on the scoring functions and octanol–water partitioning coefficient, we chose 4-isoprophylphenol as the most toxic product of BPA degradation, whereas phenol and hydroquinone as some of the most nontoxic compounds for zebrafish embryo toxicity test.
Zebrafish development
The effect of environmental concentrations (μg/L) of BPA, phenol, 4-isoprophylpenol, and hydroquinone on the zebrafish embryos were studied previously.9,12,14,20 In this work, the high 5–40 mg/L concentrations of selected compounds were tested since such concentrations of BPA are typically used in wastewater treatment studies,5,21,22 and also, they can be found near the production and wastewater treatment plants. For example, the concentration of BPA up to 17.2 mg/L was found in hazardous waste landfill leachates from Japan. 23 High levels (>100 mg/kg DW) were found in the sludge of wastewater treatment plants that received elevated industrial effluent. 24
Twenty milligrams per liter of BPA at 24 hpf provoked yolk extension deformations (100%), pericardial edema (56%), retardation in development (78%), and yolk defects (30%) (Fig. 3 and Table 3). At that time point, 33% of the embryos did not develop eyes and about 5% of the embryos did not have separation of the tail. At 48 hpf, BPA provoked the 65% lethality. The heartbeat rate for the embryos exposed to BPA was significantly lower than for the control (30 and 145 bps, respectively). About 25% of the embryos had otolith malformations, 11% had weaker pigmentation, and 79% developed pericardial edema.
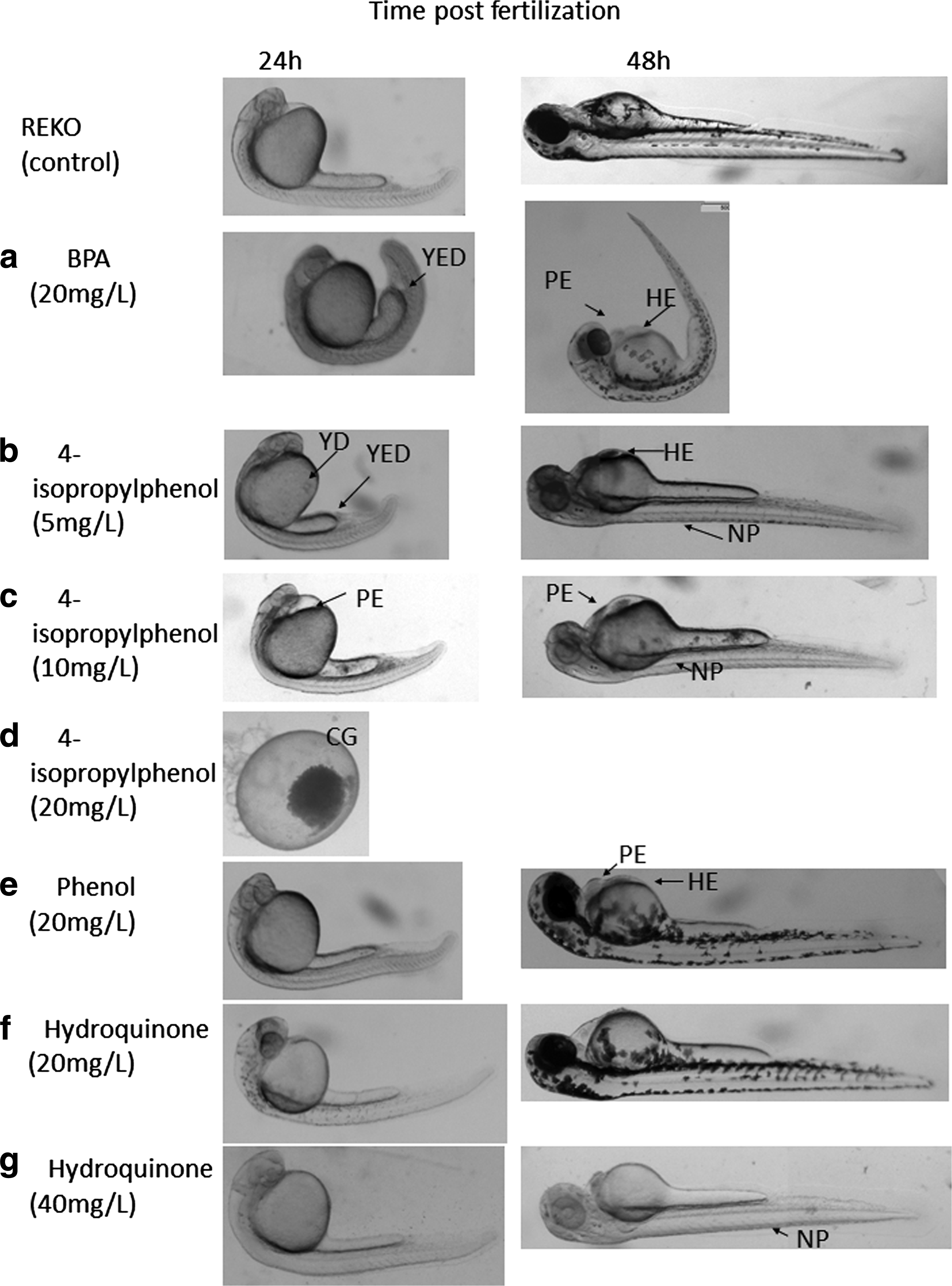
Toxicological effects on embryonic development of Danio rerio at 24 and 48 hpf. PE, pericardial edema, NP, no pigmentation, HE, hematoma, YD, yolk defects, YED, yolk extension defect, CG, coagulated. The chorions were removed from the embryos with the most representative features.
Significantly different values are highlighted in bold.
REKO was run with every set of samples, total 4 replicates, 20 eggs each.
All experiments were run in duplicates, 20 eggs each. Mortality percent is calculated based on the total amount of embryos. The rest of the parameters are calculated based on the specified number of embryos.
BPA, bisphenol A; hpf, hours postfertilization.
The same concentration of 4-isopropylphenol (20 mg/L) led to 100% mortality of the embryos at 24 hpf. The lowering of concentrations of 4-isopropylphenol to 10 mg/L decreased the lethality to 18% in 24 hpf. Yolk extension, otolith malformations, and pericardial edema decreased with a decrease of 4-isopropylphenol concentration. Thus, at 5 mg/L, only 27% of the embryos had yolk extension deformations at 24 and 48 hpf, whereas pericardial edema was observed in 16% of the embryos at 48 hpf. Yolk defects were observed in 54% of 24 hpf embryos exposed to 10 mg/L of 4-isopropylphenol (Fig. 3), and they were not observed later, at 48 hpf. Also, there were no significant changes in heartbeats in 10 mg/L 4-isopropylphenol-treated embryos compared with the control group, whereas 5 mg/L 4-isopropylphenol increased the embryos' heartbeats.
Phenol and hydroquinone were selected as the least toxic compounds based on molecular docking. Phenol (20 mg/L) caused retardation in development at 24 hpf and 13% pericardial edema and hematoma, as well as increased the heartbeat of the embryos at 48 hpf. Hydroquinone at 20 mg/L had no effect on the embryo development and a moderate effect on embryo pigmentation at 48 hpf. At 40 mg/L, hydroquinone caused retardation in the embryo development (60%), less intense pigmentation (50%), and slight decrease of the heartbeat.
Discussion
Molecular docking
Generally, the Surflex, Autodock, and Autodock Vina results are in agreement to the prediction of the most toxic and less toxic compounds. The differences could be explained by different algorithms implemented into the applied programs. Thus, Surflex splits a ligand into fragments, which are docked independently, and then the molecule structure is recreated, whereas AutoDock and AutoDock Vina use the Lamarckian version of genetic algorithm. It was shown that to obtain reliable results, it is important to use several programs. For example, in the test of 1300 molecules, AutoDock predicted better pose and score for small molecules, whereas Surflex was slightly more successful with large molecules. 25 Furthermore, since Surflex was considered to be a more accurate tool than Autodock when compared with in vivo studies, 25 below we discuss Surflex results.
BPA is known to activate human ERRγ, 26 whereas in zebrafish it activates ERRγ-B. 15 We used 4-OH-tamoxifen as a reference since it interacted with both ERRγ-A and ERRγ-B. Based on 4-OH-tamoxifen binding affinities, we could suggest that Autodock Vina program may not be accurate enough for the studies of BPA and its products of degradation since it predicted higher binding affinities for BPA than those of 4-OH-tamoxifen.
It was shown previously that only one 4-hydroxyphenyl group and one methyl group of BPA and its derivatives were required for efficient binding to human ERR-γ. 26 Moreover, this phenol moiety of BPA was found to bind to Glu275 and Arg316, whereas the methyl group interacted with aromatic amino acids Phe435 and Phe450. Our molecular docking calculations predicted similar pose and interactions for BPA (Fig. 2), 4-OH tamoxifen, and 4-isopropylphenol to those described by Okada. 26 In the case of zebrafish ERRγB, we show that two methyl groups of BPA and 4-isopropylphenol interacted with Phe288 and Phe397, and hydrogen bonds with Ala393. Moreover, our results showed that BPA and its metabolites also formed hydrogen bonds with Asn346 in human ERRγ and with Leu230 in zebrafish ERRγB. Based on those results, we could suggest that Asn346 and Leu230 might be the other important amino acids involved in the interaction between BPA-like compounds and ERRγ.
To conclude, a relatively high score of 4-isopropylphenol for ERRγ-B and similar to BPA interactions with Phe288 and Phe397, Ala393 and Leu230 could suggest similar effects on the zebrafish embryos as those of BPA.
Even though it was shown that the estrogen receptor was not a main target for BPA, still binding affinities to ERα, ERβ, and zebrafish ERα of BPA, as well as of BPA degradation products could play an important role in explaining their estrogenic effects. For example, it was shown that at 10–5 M concentration, 17β-estradiol could provoke the formation of a smaller head and larger abdomen and suppressed organogenesis in Xenopus laevis embryos. 20 It was suggested that ER might be involved in the induction of those developmental defects in X. laevis embryos. Similar effects were observed for BPA, which could suggest a similar mechanism of toxicity through ERs. 27 Our molecular docking results revealed high binding affinity of 4-isopropylphenol to ERs, especially for ERβ. We could expect that it would have a similar mechanism of action to that of BPA and provoke similar defects in zebrafish embryos.
The high correlations of ranks of the studied compounds for human and zebrafish receptors suggest that toxic effects whose mechanism involves ERRγ and ERα could be approximated from zebrafish studies to higher vertebrates.
Zebrafish screening
Based on zebrafish studies, we could align the toxicity of the studied compounds at the same concentration as 4-isopropylphenol> BPA> phenol> hydroquinone.
Our studies of zebrafish embryo exposure to BPA at 20 mg/L are in agreement with earlier reports, where BPA provoked tail deformation, pericardial edema, 9 otolith malformation, 12 and a decrease in heartbeat. 28 Previous BPA studies indicate that there are several paths/mechanisms leading to such toxic effects. It was shown that BPA is an agonist of ERγ. 15 In zebrafish, binding of BPA to ERγ-B provokes otolith abnormalities. 15 The BPA affects the cardiovascular system by induction of reactive oxygen species production, a decrease in nitric oxide concentration, ion channel inhibition/activation, disruption of Ca2+ handling, and lowering the activity of acetylcholinesterase. 29 This could lead to vasoconstriction and decreased blood supply to the heart, as well as to reductions in heartbeats. In zebrafish exposed to BPA, the decreased heartbeats were reported. 28
In our studies, 4-isopropylphenol appeared to be more toxic than BPA at the same concentration of 20 mg/L. At the same time, it could have a partly similar mechanism of toxicity to those of BPA since otolith malformation, yolk extension deformation, and pericardial edema were observed. Even though 4-isopropylphenol induced the formation of a third otolith, while BPA led to aggregation of otoliths, the underlying mechanism of otolith malformations is similar. It was shown that otic vesicle abnormalities were strongly correlated with the binding affinity of BPA and BPA-like compounds to zebrafish ERRγ. 15
However, 4-isopropylphenol did not significantly reduce the heartbeats. The 5 mg/L of 4-isopropylphenol led to an increase in heartbeat rate as it was in the case of 20 mg/L phenol and 20 mg/L hydroquinone. This suggests another mechanism of action compared with BPA.
Two percent hydroquinone (20 g/L) was used as a safe depigmenting agent for the last 50 years. 30 Previous zebrafish studies of hydroquinone report that it suppresses embryo pigmentation at 11 mg/L concentration 13 with no lethal endpoints. In our studies, we also did not observe toxic effects on the embryo development in 20 and 40 mg/L hydroquinone. 30 However, highly toxic properties of hydroquinone at 6 mg/L were already presented by Lahnsteiner, 31 which could be due to exposure of the embryos to hydroquinone at an earlier stage (16-cell stage, 1.5 hpf) than we used in our studies (5 hpf).
Based on our results, the degradation of BPA under photo-Fenton reaction could lead to the formation of a more toxic compound, that is, 4-isopropylphenol. The results of our studies on molecular docking and octanol–water partitioning coefficient calculations suggest high toxicity of 4-isopropylphenol, which is confirmed by zebrafish embryo test results. The zebrafish studies showed that 4-isopropylphenol could have some toxic effect similar to BPA mechanisms, but further investigations are required to confirm this supposition. Also, based on molecular docking and octanol–water coefficients, other candidates for zebrafish screening should be 3-methylbenzo[b]tetrahydrofuran and 2-methylbenzo[b]tetrahydrofuran. Our results obtained from zebrafish embryo test could be extrapolated to human as there is good correlation between docking results for human ER and zebrafish ER. However, the differences in binding affinities of the compounds, as well as their pose and formation of hydrogen bonds in the case of human and zebrafish receptors, should be taken into account when approximating the toxicity results from zebrafish to human.
The combination of molecular docking, octanol–water partitioning coefficient calculations, and zebrafish embryo development test could be applied to other xenoestrogens and their products of degradation studies as a less time-consuming and less labor-intensive alternative to animal testing.
Footnotes
Acknowledgments
The research was supported by National Science Centre (NCN grant 2013/11/D/NZ7/02346). Molecular docking calculations were performed using Sybyl in the Interdisciplinary Center for Mathematical and Computational Modeling (ICM) at Warsaw University under the computational Grant G14-6. We acknowledge the use of the Zebrafish Core Facility at the International Institute of Molecular and Cell Biology in Warsaw, Poland, supported by FishMed project within the EU Seventh Framework Programme (no. 316125) and grants from Polish Ministry of Science and Higher education. We wish to thank Małgorzata Wiweger (PhD) of Zebrafish Core Facility, International Institute of Molecular and Cell Biology in Warsaw, for her valuable advice and assistance during the zebrafish experiments.
Disclosure Statement
No competing financial interests exist.