
Abstract
Abstract
It has been demonstrated that glyphosate-based herbicides are toxic to animals. In the present study, reactive oxygen species (ROS) generation, antioxidant capacity against peroxyl radicals (ACAP), and lipid peroxidation (LPO), as well as the activity and expression of the acetylcholinesterase (AChE) enzyme, were evaluated in Danio rerio males exposed to 5 or 10 mg/L of glyphosate for 24 and 96 h. An increase in ACAP in gills after 24 h was observed in the animals exposed to 5 mg/L of glyphosate. A decrease in LPO was observed in brain tissue of animals exposed to 10 mg/L after 24 h, while an increase was observed in muscle after 96 h. No significant alterations were observed in ROS generation. AChE activity was not altered in muscles or brains of animals exposed to either glyphosate concentration for 24 or 96 h. However, gene expression of this enzyme in the brain was reduced after 24 h and was enhanced in both brain and muscle tissues after 96 h. Thus, contrary to previous findings that had attributed the imbalance in the oxidative state of animals exposed to glyphosate-based herbicides to surfactants and other inert compounds, the present study demonstrated that glyphosate per se promotes this same effect in zebrafish males. Although glyphosate concentrations did not alter AChE activity, this study demonstrated for the first time that this molecule affects ache expression in male zebrafish D. rerio.
Introduction
G
Herbicides containing glyphosate are used in large scales as several commercial formulations (Roundup Original®, Roundup Transorb®), which may include substances identified as inert surfactants. 4 Such formulations have presented higher toxicity compared with glyphosate in isolation.5,6 Aquatic animals are susceptible to these effects, while fish have been the most studied and characterized,7–22 and the toxicity mechanisms were those linked with oxidative stress and cholinesterase (ChE) interference.
A state of oxidative stress in an organism is a common response to exposure to pesticides. Interference with bioenergetics and detoxification pathways may be induced by pesticide exposure, which in turn leads to higher generation of reactive oxygen species (ROS) and inhibition of antioxidant capacity. 23 Specifically, it has been demonstrated that glyphosate exposure could increase ROS generation.6,7,24 Furthermore, commercial formulations of glyphosate can cause alterations in the activity of enzymes related to the antioxidant defense system,7–10 as well as lipid peroxidation (LPO) in fish.11–13 Such alterations in antioxidant status would be associated with mitochondrial damage.15,26 Roundup®, a glyphosate-based herbicide, has recently been demonstrated to induce alterations in the redox balance of zebrafish tissues, including the expression of genes related to the antioxidant defense system. 15 Such effects have been attributed to the interaction of glyphosate with the respiratory electron chain, and the inhibition of mitochondrial complexes. The reduced energetic efficiency of mitochondria may account for some toxic effects of glyphosate-based herbicides, resulting in an impairment of cellular energetic pathways and an increase in the generation of ROS. 26
Another toxicity mechanism of glyphosate-based herbicides in animals is the alteration of ChE activity. Acetylcholinesterase (AChE) is a member of the enzyme family known as ChE and is responsible for degrading the neurotransmitter acetylcholine in cholinergic synapses. AChE is usually attached to the extracellular side of the plasma membrane of vertebrate and invertebrate cells, where it controls ionic currents in excitable membranes and plays a pivotal role in the nerve conduction process and at the neuromuscular junctions. 17 It can also occur in a soluble monomeric form in the cytoplasm. The accumulation of acetylcholine caused by AChE inhibition results in a cholinergic hyperactivity. 17 Some studies have demonstrated that exposure to commercial formulations containing glyphosate decreases AChE activity,10,11,18,19 although glyphosate is not considered to be a classical AChE inhibitor. 27 However, recent works have demonstrated that glyphosate inhibits ChE activity in various tissues of fish; specifically Danio rerio and Jenynsia multidentata, and mussels, Perna perna, exposed in vitro to concentrations in the millimolar range. 20 Glyphosate effects on the expression of the AChE gene were not investigated.
With respect to aforementioned results and the lack of literature regarding specific effects of glyphosate, the objective of the present study was to evaluate the effects of exposure to glyphosate for 24 and 96 h on oxidative stress parameters in the fish D. rerio, as well as the activity and expression of AChE.
Materials and Methods
Animals and treatment
This study was approved by the Ethics Committee for Animal Use from the Federal University of Rio Grande-FURG (CEUA-FURG). All handling and maintenance of the zebrafish was in compliance with the Westerfield. 28 Adult zebrafish (Danio rerio) were obtained from commercial distributors (Redfish, Porto Alegre – RS, Brazil) and were maintained according to the protocols for the species. In this sense, only male animals were employed in the present study to avoid biased data due to sex differences between samples.
The experimental setup employed in the present study were previously described in the study of Lopes et al. 14 Briefly, the animals were maintained in tanks with dechlorinated and aerated tap water, at 28°C ± 2°C, pH 7.0 ± 1, with a photoperiod of 12 hours light and 12 hours dark. The nitrogen compounds levels were monitored during the experimental period with commercial kits (Labcon) and maintained at levels considered safe. The fish were fed ad libitum daily with commercial fish food. After acclimation period, the animals were divided into three experimental groups. The control group was maintained in dechlorinated water and the exposed groups were treated with pure glyphosate (Analytical Standard - SIGMA). Nominal concentrations of 5.0 (low) and 10.0 mg/L (high) were used based on previous studies that exposed fish species to the commercial formulation of this herbicide7–11 and or pure substance. 14 The animals were exposed for up to 96 hours, with water replacement after 48 hours in order to maintain quality. The experiments were performed in 2 L aquariums, each containing three fish with a mean animal length and weight of 37.9 ± 0.6 mm and 0.541 ± 0.2 g, respectively. The water conditions (temperature, pH, nitrogen compounds) during the experimental period were monitored. Water samples were collected after 48h for glyphosate measurements. Animals were randomly sampled after 24 and 96 h in each group and consequently anesthetized and sacrificed. Brains, gills and muscles were immediately excised and frozen at −80°C for subsequent analyses. These procedures comply with the protocols defined by the AVMA Guidelines. 29
Glyphosate determination
Glyphosate concentration in water samples was determined in accordance with Harayashiki et al., 21 using an ion chromatograph (IC Compact 881; Metrohm, Herisau, Switzerland) with conductometric detector, using an ion exchange column (Metrosep A Supp 5 150/4.0) and a chemical suppressor. Data collection and treatment were performed using the software MagIC Net 2.3 (Metrohm).
Generation of ROS
ROS generation was analyzed using the dye dichlorofluorescein diacetate (H2DCF-DA) protocol. 30 Brain, gill, and muscle tissues from pools of three animals (n = 4) were homogenized (1:9 w/v) in cold buffer (100 mM Tris-HCl, 2 mM EDTA and 5 mM MgCl2·6H2O, pH = 7.75). The homogenates were centrifuged at 20,000 g for 20 min at 4°C. Total protein content in the obtained supernatant was measured with a commercial kit (Doles Reagentes Ltd., Goiânia, Brazil) based on the Biuret protein assay. Samples were diluted with the homogenization buffer to 2.2 mg/mL of protein to standardize the protein content. Samples were then loaded (10 μL) into a plate with the reaction buffer (127.5 μL of 30 mM HEPES, 200 mM KCl and 1 mM MgCl2, pH = 7.2). After determining the background fluorescence, H2DCF-DA was added. Fluorescence intensity was monitored for 60 min at 28°C using a fluorometer (VICTOR2; Perkin Elmer) with excitation and emission wavelengths of 485 and 520 nm, respectively. Fluorescence intensity curve areas were integrated, and the total area was used for comparisons. The results are expressed as percentage fluorescence area × min (FA × min) compared to the control group.
Antioxidant capacity against peroxyl radicals
The same samples used in ROS determination were also used for ACAP analysis. ACAP was evaluated using the measurement of ROS with or without addition of the peroxyl radical generator 2,2-azobis 2 methylpropionamidine dihydrochloride (ABAP; 4 mM; Aldrich). Fluorescence intensity was monitored for 60 min at 37°C, using a fluorometer (VICTOR2; Perkin Elmer), with excitation and emission wavelengths of 485 and 520 nm, respectively. The difference between the FAs with and without ABAP was used as the measure of antioxidant capacity. The data were expressed as 1/percent of the relative area of fluorescence of the experimental samples compared to the control group, where higher values of ACAP indicate greater antioxidant capacity. 31
Lipid peroxidation
LPO was determined using the ferrous ion oxidation xylenol orange (FOX) method. 32 Pools of the brains, gills, or muscles from two animals for each organ (n = 6) were homogenized in methanol at a ratio of 1:9 (w/v) and centrifuged at 1000 g for 5 min at 4°C. The lipid hydroperoxides were determined using FeSO4 (1 mM), prepared immediately before use, along with H2SO4 (0.25 mM) and xylenol orange (1 mM). Sample absorbance was measured in a microplate reader (580 nm) after incubation at room temperature for 4 h for brain and muscle and 6 h for gills. Cumene hydroperoxide (CHP) was used as a standard. The results are expressed as nmoles CHP/g wet weight.
AChE activity
The AChE activity was measured according to the protocol of Ellman et al. 33 and adapted by Rao et al. 34 Samples of brain and muscle tissue from pools of three animals (n = 4) were homogenized in 50 mM phosphate buffer containing 20% glycerol (pH = 7.4) and centrifuged at 9000 g for 20 min at 4°C. The resulting supernatants were designated the S9 fraction (cytosolic). The pellets were suspended in the same buffer containing Triton X-100 (0.5%) and stirred for 30 min at room temperature. Resuspended pellet samples were then centrifuged at 9000 g for 30 min at 4°C. The obtained supernatants were designated the TX S9 fraction (membrane-bound). Total protein content in each of these fractions was measured as described above. Acetylthiocholine iodide (1 mM) and 5,5-dithio-bis 2-nitrobenzoic acid (DTNB) were used as the substrates, and the change in absorbance at 412 nm was monitored for 2 min at 25°C and pH 7.2. The results are expressed as nmoles of acetylcholine iodide hydrolyzed per minute per mg of protein.
Gene expression
The AChE gene (ache) expression was analyzed using the methodology described by Rosa et al. 35 Total RNA was extracted from pooled samples of brain and muscle from three animals (n = 4) using the TRIzol reagent. RNA concentration was determined spectrophotometrically (260/280 nm), and its integrity was verified using agarose gel (1%) electrophoresis. RNA was treated with DNAse I (Applied Biosystems) following the manufacturer's instructions. Complementary DNA (cDNA) was prepared from the total RNA using the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems). The obtained cDNA was used as a template for the amplification of the acetylcholinesterase gene using specific primers designed on the basis of the gene sequences available at GenBank (www.ncbi.nlm.nih.gov). The primers are described in Table 1. Quantitative polymerase chain reaction (PCR) was performed with an ABI Prism 7300 Sequence Detection System (Applied Biosystems) using SYBR-Green PCR Master Mix (Applied Biosystems). The β-actin and β-2-microglobulin (b2m) gene expression was used to normalize the ache expression in brain tissue. Elongation factor 1 alpha (ef1α) and b2m were used to normalize ache expression in muscle tissue.
AChE, acetylcholinesterase.
Statistical analysis
The data from the biochemical analyses were compared using analysis of variance, with a significance level of 5%, following confirmation that the prerequisites for homoscedasticity and homogeneity were met. When the differences were significant, the means of groups were compared by the a posteriori Newman–Keuls test with a significance level of 5%. The data for the gene expression analyses were compared using REST software. 36
Results
Glyphosate measurement
Glyphosate concentrations measured in experimental media were 4.5 ± 0.6 mg/L for the lower glyphosate concentration (nominal concentration = 5 mg/L) group and 8.6 ± 0.1 mg/L for the higher glyphosate concentration group (nominal concentration = 10 mg/L).
Generation of ROS
No significant differences were observed for the generation of ROS in the brains after 24 and 96 h of exposure to glyphosate compared to the control group (Fig. 1A). The same was observed for the gills (Fig. 1B) and muscle (Fig. 1C) at both experimental times.

Concentrations of the ROS in brain
Antioxidant capacity against peroxyl radicals
Glyphosate exposure resulted in no significant effects on the ACAP in the brains (Fig. 2A) or muscle (Fig. 2C) after 24 and 96 h compared to the control group. No significant differences were observed after 96 h of exposure in gill tissues for the same analysis. However, at the 24-h time point, there was a significant twofold increase in the antioxidant capacity in the group exposed to the lowest glyphosate concentration compared to the control group and to the animals exposed to the highest concentration (Fig. 2B).

The antioxidant capacity (ACAP) in the brains
Lipid peroxidation
There was a reduction of 45% ± 1% in LPO in the brains at the highest concentration after 24 h of exposure, compared to control group animals and the animals exposed to the lowest glyphosate concentration, but this difference was absent after 96 h (Fig. 3A). No significant differences were observed in the gills at either concentration or exposure time (Fig. 3B). After 24 h, no differences in LPO were observed in muscle tissue. However, at the 96-h time point, there was a significant increase of 30% ± 2% in the muscles of the animals exposed to the higher concentration compared to the other experimental groups.

The lipid peroxidation in the brains
AChE activity and gene expression
Exposure to glyphosate did not affect the AChE activity in either cellular fractions of the muscle or brain at either experimental time (Fig. 4). A significant decrease in AChE expression was observed in brain tissue after 24 h of exposure. Specifically, the expression relative to the control was reduced by factor of 0.8 at lower concentration and 0.6 at higher glyphosate concentration (Fig. 5A). However, after 96 h of exposure to the higher concentration, the expression of ache in this tissue increased 15-fold (Fig. 5A). In muscle tissue, the expression of the ache gene was not altered after 24 h of exposure (Fig. 5B). However, there was a 3.7-fold increase for the animals exposed to the highest glyphosate concentration after 96 h (Fig. 5B).

The activity of acetylcholinesterase in the brain fraction S9, brain fraction TXS9, muscle fraction S9, and muscle fraction TXS9 of D. rerio exposed to glyphosate (0, 5, and 10 mg/L) for 24 h
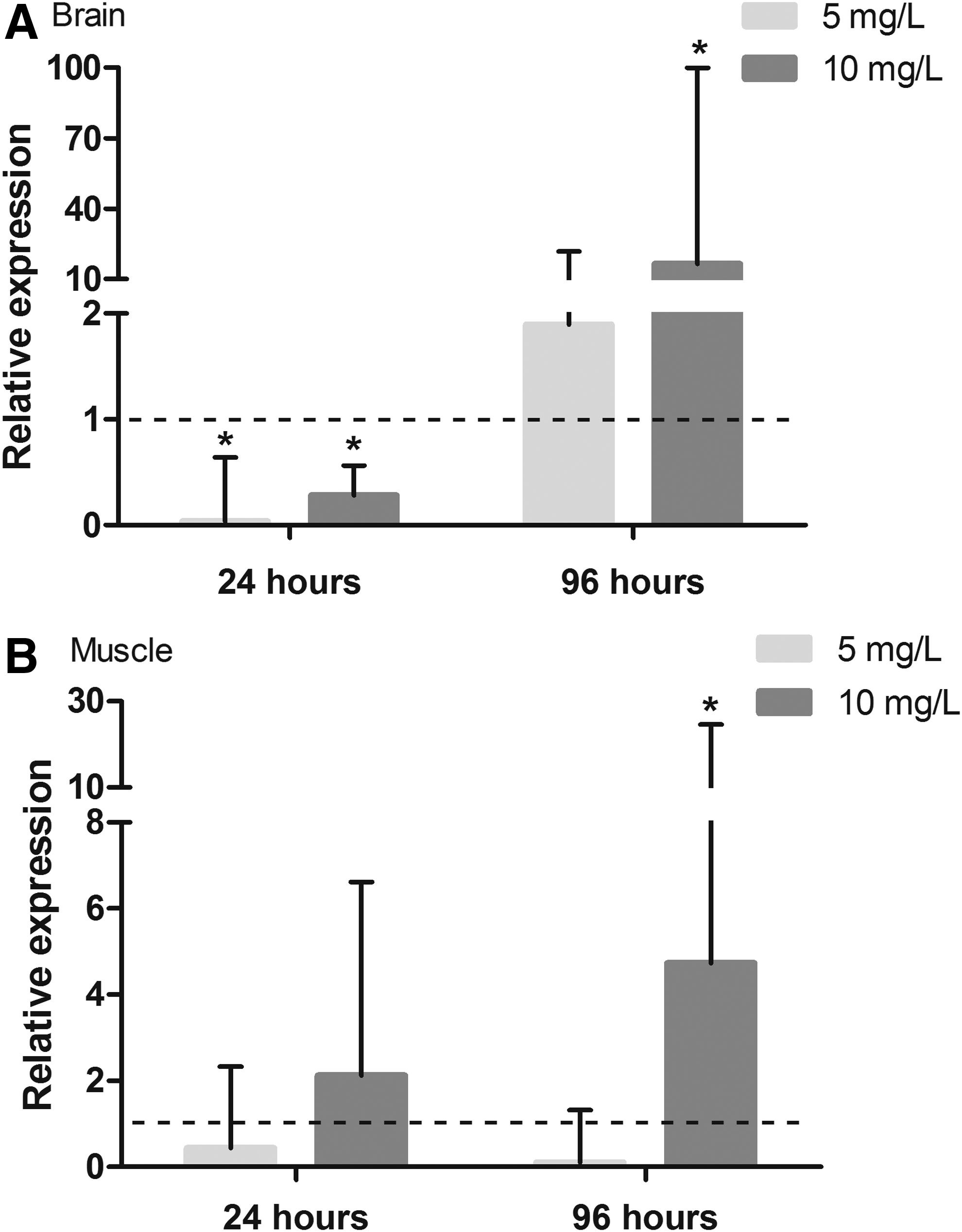
The relative acetylcholinesterase expression in the brains
Discussion
Pesticide exposure, in general, can generate oxidative stress in animals. 23 Oxidative stress is characterized by an imbalance between prooxidant and antioxidant processes, in favor of the prooxidant, resulting in damage to biomolecules (DNA, proteins, and lipids) and disruption of redox signaling.37,38 To determine whether glyphosate causes oxidative stress in D. rerio, the effects of its exposure were evaluated using a triad of oxidative stress indicators: ROS generation, antioxidant capacity, and oxidative damage to lipids.
In this study, no significant differences in the generation of ROS from any analyzed tissue were observed at either of the times examined. Roundup, at concentrations between 0.084 and 0.84 mg/L, has been observed to inhibit some enzymes of the electron transport chain complex in mitochondria isolated from rat kidneys. 26 The same study determined that glyphosate alone, even at a higher concentration (2.47 g/L), did not cause this effect. This author speculated that other substances present in the commercial formulation could have promoted such effects, or the synergistic effect of glyphosate and those constituent substances. However, in another study, an increase in ROS generation was observed in human keratinocytes exposed in vitro to glyphosate (from 1.65 to 16.5 mg/L) for 24 h. 6 These concentrations are in the same range as those in the present study. It must be noted that glyphosate was administered in the water in the present study and not directly into the cell culture as in the aforementioned study. Exposure of aquatic organisms through water involves the processes of absorption and body distribution, among others. Consistent with this idea, Poecilia vivipara exposed in vivo to Roundup did not experience effects regarding ROS generation. 21 These data corroborate evidence that glyphosate and its commercial formulations cause no alterations in oxidative stress through increases in the generation of ROS.
Although no glyphosate effects regarding ROS generation were shown, an alteration in the oxidative state of gills was observed through alterations to ACAP. The present study used an approach that considers the total antioxidant capacity of the tissue against peroxyl radicals. This method is advantageous because it considers the contribution of all antioxidants present in the tissue, rather than the contribution of isolated enzymes or low molecular antioxidant scavengers. This method therefore provides a broader view of the antioxidant status in the tissue. 39 Previous reports investigating the effects of glyphosate and glyphosate-based herbicides have evaluated selected enzymes and demonstrated several alterations in their respective activities, some of which were reduced, while others were enhanced. In the present study, an increase in the antioxidant capacity of the gills was observed in animals exposed to the lower concentration after 24 h. The gills of fish may be a target organ for pollutants because they are the first site of external contact for the organism and, thus, represent an important exposure route. These results are in agreement with those of other authors who observed an induction in the antioxidant capacity of animals, 24 as well as an increase in antioxidant enzymatic activity in fish exposed to Roundup. 22 However, in the present study no alterations in ACAP were observed in other tissues and experimental times. These findings corroborate those of Harayashiki et al., 21 who observed no effect from exposure to 0.1 and 0.7 mg/L of Roundup (glyphosate equivalent) on antioxidant capacity in gill, liver, and muscle tissues of P. vivipara after 96 h. Alterations in the initial exposure period would mitigate damage to the tissue in terms of lipid hydroperoxides and would permit the trigger of other mechanisms to deal with such stress. In this sense, biotransformation or cell extrusion mechanisms would be activated and this would be responsible for ACAP values being maintained at control levels. However, both studies evaluated only the ACAP. It is therefore possible that other antioxidants not measured by these methods could be affected in response to glyphosate exposure.
To evaluate oxidative damage to biomolecules, LPO was analyzed. LPO is a type of damage that can be caused by many factors, including ROS generation or an alteration in antioxidant capacity. 40 A decrease in this parameter in brain tissue was observed at the 24-h time point for the higher glyphosate concentration tested, while the same concentration induced an increase in the muscles after 96 h of exposure. Interestingly, Glusczak et al. 11 also observed a decrease in LPO in brain tissue and an increase in this parameter in the muscles of Rhamdia quelen after 96 h of exposure to Roundup at concentrations of 0.2 and 0.4 mg/L. These differences in LPO responses were attributed to tissue-specific characteristics, including variations in the mechanisms of antioxidants and the fact that different fish tissues present different levels of peroxide production.40,41 It is important to note that, according to the present results, the ACAP is higher in the brain than in muscle. This feature could explain the differences in LPO between these tissues (data not shown). This finding is an indication of the selective protection of the brain against an increase in the generation of ROS. However, after 96 h, LPO levels returned to control group levels in the brains. This return is likely a consequence of other mechanisms of defense that would be triggered to protect tissue from glyphosate toxicity. The observed increase in AChE gene expression in this tissue at this same period is an example of this type of triggered defense.
According to Oruç and Usta, 17 LPO may be associated with decreases in AChE activity. The accumulation of acetylcholine caused by AChE inhibition results in a cholinergic hyperactivity that can initiate an accumulation of ROS and lead to LPO. 17 Therefore, the effects observed in the previous studies concerning lipid oxidation in brain and muscle could be an indication of an imbalance in cholinergic signaling, leading to prooxidant effects. Although an alteration in the lipid oxidation profile was observed in the present study, no decrease in AChE activity was observed in either muscle or brain.
Even though no alteration in AChE activity after glyphosate exposure was observed in the present study, there were changes in ache gene expression in the muscles and brains of zebrafish exposed to glyphosate. Glyphosate is not considered a classical AChE inhibitor. However, Sandrini et al. 20 showed that glyphosate was capable of inhibiting AChE activity in the brain and muscle of two fish species (D. rerio and J. multidentata), as well as in invertebrate tissues (P. perna), in vitro. It should be noted that the IC50 observed for both tissues in D. rerio is higher (1100 mg/L) in brain S9 fraction than in the TXS9 fraction (400 mg/L), although similar IC50 values were described for the muscle S9 and TXS9 fractions (850 and 800 mg/L, respectively). Although differences in the sensitivity were observed in vitro in the present study, no significant differences were observed in AChE activity in brain or muscle tissue after 24 or 96 h of exposure to both glyphosate concentrations in any fraction. In addition, the glyphosate concentrations used in the present work were 41 times lower than those used in the aforementioned study. In contrast, studies with Roundup showed a decrease in AChE activity in the brain and muscle of fish after exposures for 24, 96, and 120 h.9–11,18 The effects of commercial herbicides may be attributed to an increase in the absorption of the pesticide facilitated by the surfactant present in its formulation or as a result of the surfactant itself.
Although AChE activity was not affected by glyphosate exposure, its genetic expression was altered in both of the tissues analyzed. It has been previously demonstrated that exposure to organophosphate pesticides promotes an alteration in ache expression; 26 however, this has not been investigated specifically for exposure to glyphosate or its commercial formulations. In the present study, ache expression decreased in the brain after 24 h for both concentrations and increased in both the brain and muscle after exposure to 10 mg/L of glyphosate for 96 h.
Alterations in cellular signaling and gene expression are essential processes following exposure to a stressor. Exposure to sublethal concentrations of pesticides or other stressor agents induces short- or long-term responses that involve cholinergic systems. 42 According to Soreq et al., 43 several endogenous and exogenous stimuli can induce rapid and long-lasting upregulation of ache expression. These stimuli include psychological or physical stress, which in turn may induce cholinergic excitation through the release of acetylcholine. Thus, elevated cortisol levels and consequent overexpression of AChE are expected. Furthermore, the stimulation of AChE synthesis and its activity would protect animals from chemical stressors such as anti-AChE molecules. 43 In this manner, stress would indirectly affect ache expression and activity. In addition, anti-AChE agents would directly alter ache expression. 44 It has been demonstrated that the ache promoter region contains multiple response elements, including sites for egr-1 protein binding. Egr-1 protein is a factor that enhances transcription of the ache gene. This protein becomes concentrated in the nucleus as a result of a signaling cascade activated by the muscarinic acetylcholine receptors following the accumulation of ACh in the synaptic cleft. Thus, inhibition of AChE may result in accumulation of the neurotransmitter ACh and, subsequently, the induction of the expression of ache messenger RNA (mRNA).
A possible explanation for the results shown in the present study is that an alteration in AChE activity might have occurred at a different time than those selected for analysis. The alteration in the level of ache mRNA expression suggests that the amount of the enzyme AChE present was not enough to degrade the acetylcholine. Therefore, the enzyme activity may have appeared to remain unchanged because an increased rate of synthesis of the enzyme was able to compensate for the inhibition process. An unexpected result observed in the present study was the repression in ache expression in brain tissue in the first 24 h of glyphosate exposure. Similar results were observed in the common carp exposed to chlorpyrifos, an organophosphate pesticide that is a classical AChE inhibitor, 45 but the mechanism for this effect is not well understood.
Conclusion
Exposure to glyphosate per se promotes a physiological response in male D. rerio tissues (brain, muscle, and gills). Glyphosate exposure caused an imbalance in the oxidative status and altered the cholinergic system in a tissue-dependent manner. These results are consistent with the toxicity mechanisms previously described for the commercial formulation.
Footnotes
Acknowledgments
F.M.L. is a post-graduate student financed by CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—process no. 42004012008P9). The authors thank the National Institute of Science and Technology—Aquatic Toxicology (INCT-TA) from Conselho Nacional de Desenvolvimento Científico e Tecnológico—process no. 573949/2008-5 and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq—process no. 480919/2013-5). E.G.P. was a research fellow from the Brazilian CNPq.
Disclosure Statement
No competing financial interests exist.