
Abstract
Abstract
Gymnotus coatesi is a small and rare species of banded knife fish that was originally described by LaMonte in 1935, found along the main stretch of the Amazon River. There is no described cytogenetic data on this species. We analyzed the karyotype of five specimens of G. coatesi collected from Cururutuia Stream in Bragança, Pará, Brazil. The obtained diploid number is 50 and the karyotypic formula is 24 m/sm +26 st/a. The constitutive heterochromatin is DAPI positive and distributed mainly in the centromeric and pericentromeric regions of the chromosomes. Ag-nucleolus organizer regions staining showed nine active sites. The 5S rDNA probe hybridized chromosome pair 17 in the interstitial part of the long arm. Fluorescence in situ hybridization (FISH) with telomeric probes revealed signals only at terminal regions of the chromosomes. The 18S rDNA probe hybridized to 21 sites, and these signals colocalized with the telomeric sequences. This relatively high number of 18S rDNA sites may reflect gene duplication mediated by transposable elements. These results indicate that although the diploid number of G. coatesi is within the range previously observed for other members of the genus, various karyotypic characteristics distinguish G. coatesi from the other species of the genus and members of the Gymnotiform order.
Background
G
Materials and Methods
Five specimens of G. coatesi (three male and two female) were collected in Igarapé Cururutuia, Bragança, Pará, Brazil, S01°04′42.0″ W046°44′18.6″ (Fig. 1). Chromosomes were prepared from kidney tissues as described, 13 with some modifications. The prepared chromosomes were analyzed by Giemsa staining, C-banding, 14 and AgNO3 staining. 15 Fluorescence in situ hybridization (FISH) was performed following the protocol of Ref., 16 using telomeric sequence, TTTAGG (All Telomere probes, Oncor) probes, 18S rDNA probe, 17 and 5S rDNA probe obtained from G. carapo. The probes were labeled with biotin or digoxigenin by nick-translation, and the signals were detected with avidin (Cy3 or FITC) or anti-digoxigenin-FITC. The karyotypes were classified as described by Ref. 18

Gymnotus coatesi specimen (above) and place the specimens were collected in Bragança, Para (below).
Results
The analyzed specimens of G. coatesi showed 2n = 50 and a karyotypic formula (KF) = 24 m/sm +26st/a, with heteromorphism observed in the sizes of chromosome pairs 1, 3, and 5, and the short arms of pairs 8, 9, and 15 (Fig. 2a, b). There was no evidence of morphologically differentiated sex chromosomes in our meiotic analyses (Supplementary Fig. S1; Supplementary Data are available online at www-liebertpub-com.web.bisu.edu.cn/zeb).
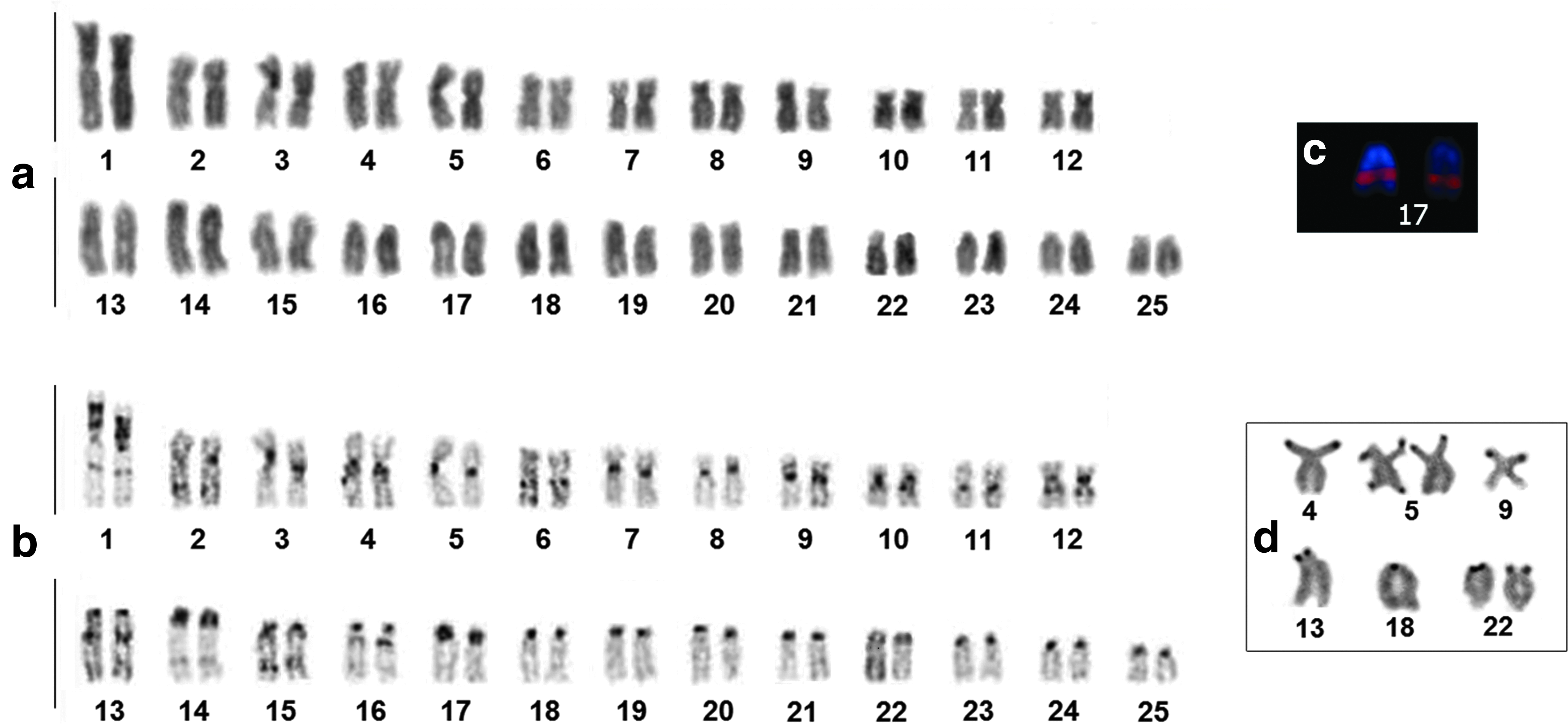
G. coatesi karyotype.
Constitutive heterochromatin was found in the centromeric/pericentromeric regions of all chromosomes: pair 1 has a large heterochromatic block along almost all of the short arm and a small band in the interstitial region of the long arm; pairs 6, 10, 12, 13, and 16 have heterochromatic bands in the proximal regions of their long arms; pair 9 has heterochromatic bands in the proximal regions of their short arms; and pairs 2, 14, and 15 have heterochromatic bands in the interstitial regions of their long arms (Fig. 2b).
The 5S rDNA probe hybridized to the interstitial region of the long arm of pair 17 (Fig. 2c). G. coatesi was found to have multiple NOR sites, including nine active sites on eight chromosomes, with pair 5 showing sites in the distal regions of both arms (Fig. 2d). The 18S rDNA probe hybridized to 21 sites on 19 chromosomes (Fig. 3a). The telomeric probe hybridized specifically to the distal regions of all chromosomes (Fig. 3b). All of the 18S rDNA signals colocalized with the telomeric signals (Fig. 3c).
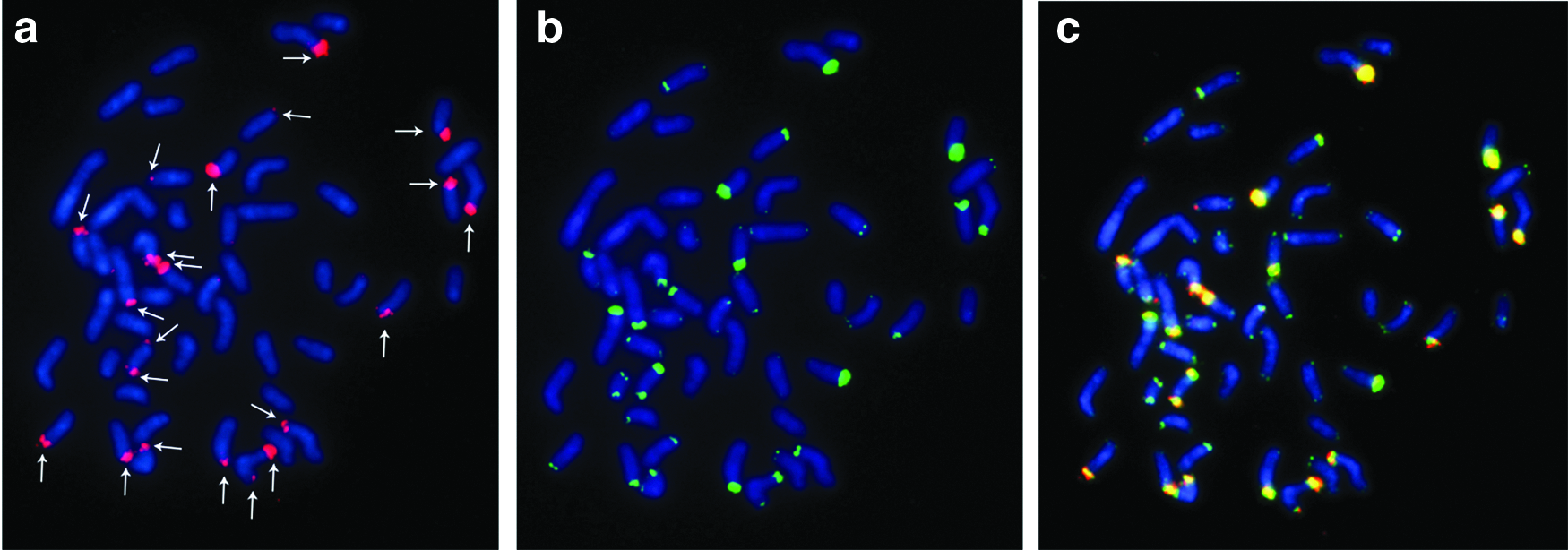
FISH in G. coatesi:
Discussion
The diploid number of G. coatesi (2n = 50, KF = 24 m/sm +26st/a) is shared with other species of this genus, such as Gymnotus sp. (2n = 50, KF = 26 m/sm +24 st/a 19 ;), Gymnotus sp. ‘‘Negro’’ (2n = 50, KF = 22 m/sm +28 st/a in females, and 21 m/sm +29 st/a in males 20 ;), and two other species of the G. coatesi clade, G. javari (2n = 50, KF = 20 m/sm +30 st/a 21 ;), and G. coropinae (2n = 50/49, KF = 28 m/sm +22 st/a in females and 26 m/sm +23 st/a in males 20 ). The interspecific differences in KFs likely reflect rearrangements that did not modify the diploid number, such as inversions and translocations.
In recent studies2,4,5 G. jonasi is placed as the most basal specie within the G. coatesi clade, followed by G. stenoleucus. 5 Here, we plotted the 2n of the species in the phylogeny adapted from the data 5 (Fig. 4), and confirmed G. jonasi basal position, with diploid number 2n = 52 (KF = 12 m/sm +40 st/a 22 ;). This is because it shares this trait with G. pantherinus 23 of G. pantherinus clade, which is a sister clade of G. coatesi, and with Electrophorus electricus, which is a sister taxa of genus Gymnotus.24,25 G. stenoleucus has 2n = 48 (KF = 20 m/sm +28 st/a, 26 ). We propose that the evolutionary chromosomal events comprised two fusions in G. stenoleucus, which changed this karyotype. G. coropinae, G. javari, and G. coatesi appear to have diverged from the basal diploid number by a fusion event.

Representation of the diploid number of the species in the clade G. coatesi, clade Gymnotus pantherinus, and Electrophorus electricus plotted according to recent phylogeny in the genus. 5
Despite sharing its diploid number with G. coropinae and G. javari, G. coatesi has a distinct KF, suggesting that rearrangements such as pericentric inversions and translocations may have occurred in the karyotypes of these species. G. coatesi shares its pattern of centromeric constitutive heterochromatin with the other species of its clade, but G. coatesi also has noncentromeric heterochromatic blocks on several chromosomes (Figs. 2b and 5), including a very obvious example on the short arm of pair 1 (Figs. 2b and 5). Similar to G. jonasi, G. coatesi lacks the morphologically differentiated sex chromosomes found in G. coropinae 20 .

Constitutive heterochromatin distribution, Ag-NOR, 5S rDNA e 18S rDNA in the karyotype of G. coatesi.
G. coatesi has 5S rDNA sites on a single pair of chromosomes (Figs. 2c and 5). In this, it is similar to Gymnotus sylvius and G. coropinae,20,27 but differs from G. paraguensis, which has these sequences on 19 chromosome pairs. 16
G. coatesi also differs from the other species of the genus in that many of its chromosomes carry 18S rDNA sequences (Figs. 2d, 3a, and 5). Within the genus, only two other species have been described as having multiple NOR: Gymnotus pantanal with two pairs 28 and G. jonasi with three pairs. 22 The presence of a high number of 18S rDNA sites, several of which are silenced, and their colocalization with telomeric sequences make G. coatesi unique among the described members of its genus. Recent studies suggest that rDNA sequences may be linked to transposable elements, and that the insertion of these elements into ribosomal sequences could mediate the relocation, dispersion, and/or activation of these sites.20,29–31 Thus, the numerous copies described herein may reflect gene duplication through retrotransposition or transposition.32,33 The colocalization of telomeric and 18S rDNA sequences may be the result of inter-spacing of those sequences, as proposed in atlantic eels 34 (Anguilla anguilla and Anguilla rostrara) and in the rainbow trout 35 (Oncorhynchus mykiss). It is not known exactly what this colocalization means to the karyotype. However, further investigations may help clarify the evolutionary chromosomal processes that may have occurred in this genus.
Conclusions
G. coatesi has a diploid number in the range observed for its genus, but it has additional traits that distinguish it from the other species of order Gymnotiformes. These include numerous 18S rDNA sequences and their co-localization with telomeric sequences. This large number of 18S rDNA sequences might reflect the amplification and dispersion of these sequences mediated by the association of mobile elements (transposons and/or retrotransposons), which makes those sequences a target for future studies.
Footnotes
Acknowledgments
This study is part of Master dissertation of MAM who was a recipient of a CAPES Scholarship in Genetics and Molecular Biology, UFPA. The authors are grateful to members of the team of cytogenetics laboratory UFPA in the field work and chromosomal preparations and to MSc. Jorge Rissino, MSc. Shirley Nascimento, and Maria da Conceição for assistance in laboratory work. Authors thank Dr. Jonathan Read for allowing the colection of samples in his grange. This research was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) through the Edital Universal (Proc. 475013/2012-3) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) through the Edital 047/2012 PRÓ-AMAZÔNIA: Biodiversidade e Sustentabilidade on a project coordinated by CYN; by FAPESPA (Pará Foundation for Supporting Science) through the National Excellence on Research Program (PRONEX, TO 011/2008) and Banco Nacional de Desenvolvimento Econômico e Social–BNDES (Operação 2.318.698.0001) on a project coordinated by JCP. CYN (308428/2013-7) and JCP (308401/2013-1) are grateful to CNPq for Productivity Grants. We also thank the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio) for the collection permit 020/2005 (Registration: 207419).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.