
Abstract
Abstract
This work constitutes the first cytogenetic characterization of a trans-Andean species of Heptapteridae. The catfish Pimelodella cf. chagresi from the Upper Rio Magdalena was studied, applying standard cytogenetic techniques (Giemsa, C-banding, and argyrophilic nucleolar organizer region [Ag-NOR]) and fluorescence in situ hybridization techniques using repetitive DNA probes: microsatellites (CA15 and GA15) and ribosomal RNA (rRNA) multigene families (18S and 5S recombinant DNA [rDNA] probes). The species showed a unique diploid chromosome number 2n = 50 (32m [metacentrics] +14sm [submetacentrics] +4st [subtelocentrics]) and a XX/XY sex chromosomal system, where the heteromorphic Y-chromosome revealed a conspicuous accumulation of all the assayed domains of repetitive DNA. P. cf. chagresi karyotype shares common features with other Heptapteridae, such as the predominance of metacentric and submetacentric chromosomes, and one pair of subtelomeric nucleolar organizer regions (NORs). These results reflect an independent karyological identity of a trans-Andean species and the relevance of repetitive DNA sequences in the process of sex chromosome differentiation in fish; it is the first case of syntenic accumulation of rRNA multigene families (18S and 5S rDNA) and microsatellite sequences (CA15 and GA15) in a differentiated sex chromosome in Neotropical fish.
Introduction
T
Although the vast majority of Neotropical fish do not have differentiated sex chromosomes, 16 several sex chromosomal systems have been identified, including both simple (ZZ/ZW, XX/XY, XX/X0) or multiple systems (X1X1X2X2/X1X2Y, XX/XY1Y2, ZZ/ZW1W2, Z1Z1Z2Z2/Z1Z2W1W2).17–19 In Pimelodella, a XX/XY sex chromosome system has been described in Pimelodella boschmai and Pimelodella sp., both from the Mogi-Guaçu River Basin, Brazil.7,20 Although the evolutionary mechanisms involved in the formation of heteromorphic sex chromosomes are not fully understood, the interruption of recombination in certain chromosome regions has been proposed as the most relevant aspect of the differentiation process of these elements.21,22
Recombination disruption has been defined as a previous stage of heterochromatinization or accumulation process of repetitive DNAs in sex chromosomes.21,23 Repetitive DNA sequences constitute a large part of eukaryotic genomes, and these sequences play an important role in the structure, function, and genome evolution in fish. 24 Repetitive DNA is composed of dispersed repeats such as transposable elements (TEs) or tandem sequences, which include satellites, minisatellites, microsatellites, and multigene families such as ribosomal RNAs (rRNAs) and histone genes. 25 In fish, sex chromosome differentiation has been associated with microsatellite accumulation,26–28 satellite DNAs,29,30 TEs,31,32 as well as 5S 33 or 18S recombinant DNAs (rDNAs).34–37 However, the molecular forces that propagate and maintain different domains of repetitive DNAs in the genome remain unknown.21,38 Therefore, mapping of repetitive sequences in fish is a useful tool to infer evolutionary patterns of sex chromosome differentiation in different taxa.27,28,35 Although the study of association dynamics between repetitive DNAs and sex chromosome differentiation in fish has increased in recent decades, the available information is still limited, considering the number of species described, which constitute slightly more than one-half of the total approximate number of recognized living vertebrate species, with some 31,800 species distributed in 536 families. 39
Descriptions and karyotypic comparisons among species have been shown to be useful for the detection of cryptic species, 13 and provide useful information for evolutionary studies and understanding of speciation mechanisms. 40 This work constitutes the first cytogenetic characterization in trans-Andean Heptapteridae species. We aim to characterize the karyotype of Pimelodella cf. chagresi using standard cytogenetic techniques and mapping the chromosomal distribution of different domains of repetitive DNA, including microsatellites and rRNA multigene families, to obtain novel information about chromosomal evolutionary trends and karyotypic features within this genus.
Materials and Methods
Specimen collection
Ten specimens of P. cf. chagresi (two males and eight females) were collected from the Alvarado River, Upper Magdalena River Basin, Colombia (04°31′23.6"N 074°58′"09.2"W). Voucher specimens were deposited in the zoological collection of the Universidad del Tolima, Tolima, Colombia (collection No. CZUT-IC 12802). The collected specimens were anesthetized and euthanized with clove oil (300 mg/L), 41 as approved by the Universidade Federal de Viçosa Ethics Committee (permit 52/2015).
Karyological analyses
Mitotic simulation in vivo was induced by intramuscular application of yeast. 42 After a 12-h stimulation period, colchicine (0.0125%) was applied intraperitoneally and mitotic chromosomes were obtained from kidney cell suspensions, following Bertollo et al. 43
Slides containing mitotic chromosomes were prepared with conventional staining with Giemsa (10%), the regions of constitutive heterochromatin were evidenced using C-banding procedure, 44 and nucleolar organizer regions (NORs) were identified with silver nitrate precipitation. 45 Distribution patterns of multigene families of ribosomal genes (rDNA) and microsatellite DNA were mapped using fluorescence in situ hybridization (FISH) following Pinkel et al. 46 with modifications. Ribosomal probes 18S and 5S were labeled using polymerase chain reaction with digoxigenin-11-dUTP or biotin-16-dUTP (Roche Applied Sciences) and the signal was detected with anti-digoxigenin-rhodamine or FITC-avidin (Roche Applied Sciences). Repetitive DNA domains were characterized using microsatellite (GA)15 and (CA)15 probes labeled with Cy3 fluorochrome at the 5′ end (Sigma).
Digital images were captured in BX53F Olympus microscopes with DP73 and MX10 Olympus cameras for colored and fluorescent techniques, respectively, using cellSens imaging software (Olympus Corp.). Chromosomes were classified according to their arm ratio in metacentrics (m), submetacentrics (sm), and subtelocentrics (st). 47
Results
The analysis of mitotic metaphase cells of P. cf. chagresi in the Alvarado River, Upper Magdalena River Basin, revealed a diploid number of 2n = 50 chromosomes with a fundamental number (FN) = 100, composed of 32m, 14sm, and 4st (Fig. 1A, B). An XX/XY sex chromosome system was identified, where the X chromosome is a small submetacentric and its Y homologue is the largest of the submetacentric chromosomes (Fig. 1B).
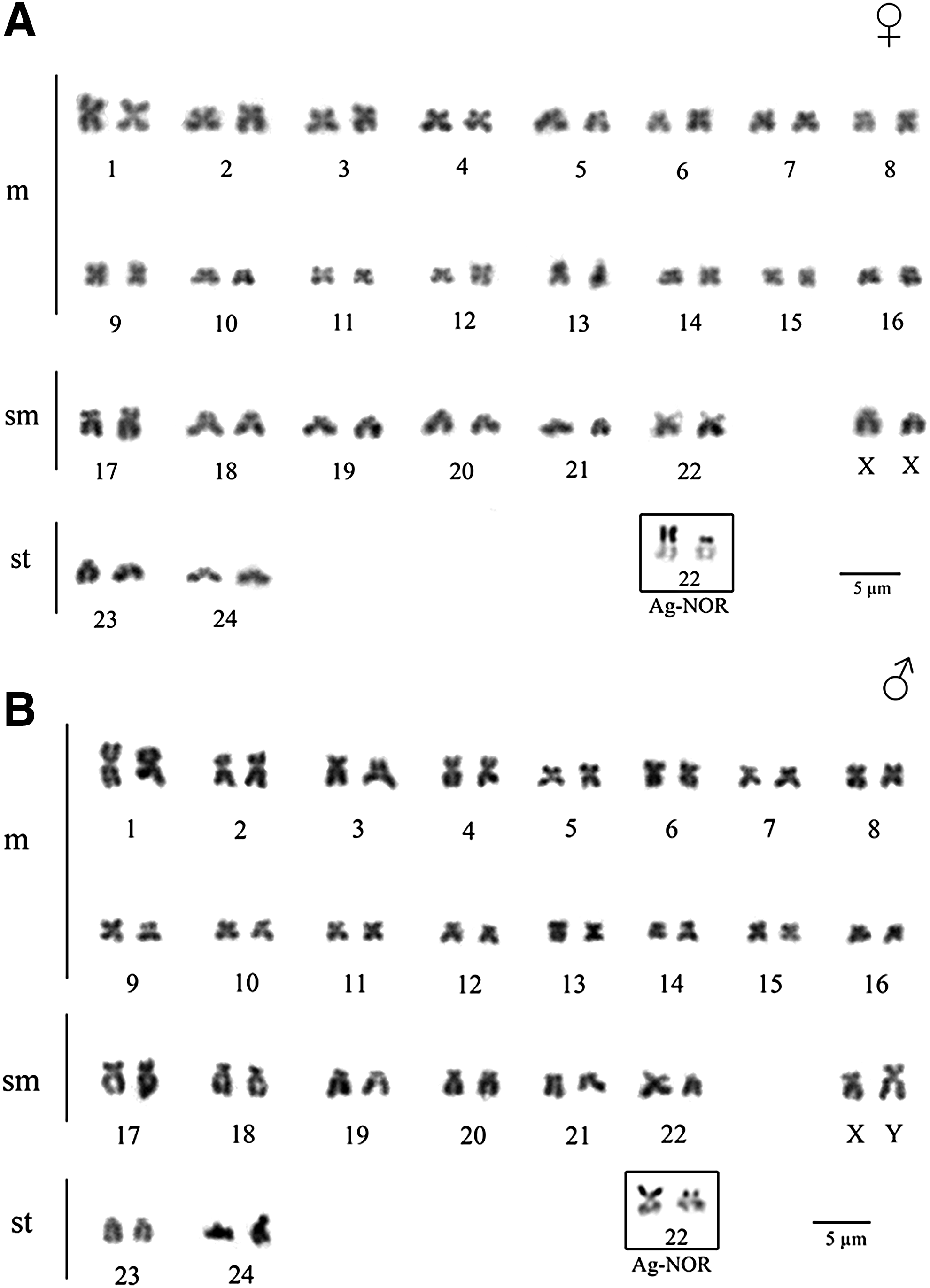
Giemsa-stained karyotype of Pimelodella cf. chagresi in females
The heterochromatic blocks were preferentially distributed at pericentromeric regions of most chromosomes; however, a metacentric pair 13 revealed a conspicuous C-band at the terminal region on long arms (Fig. 2A, B). Additionally, a conspicuous C-band was observed at the terminal region on the long arm of the Y chromosome (Fig. 2B).

Heterochromatic patterns of P. cf. chagresi in females
FISH with 18S rDNA probe (Fig. 3A, B) and argyrophilic nucleolar organizer region (Ag-NOR) sites (Fig. 1A, B) coincided, indicating that the active genes of the major ribosomal clusters (45S rDNA) occur on the terminal region of the short arms of the autosomal submetacentric pair number 22. The 5S rDNA probe revealed fluorescent signals on short arms of another autosomal metacentric pair, in a pericentromeric position (Fig. 3C, D). Additionally, a third site showed synteny for both 18S and 5S rDNA probes on the long arm of the Y chromosome (Fig. 3B, D).
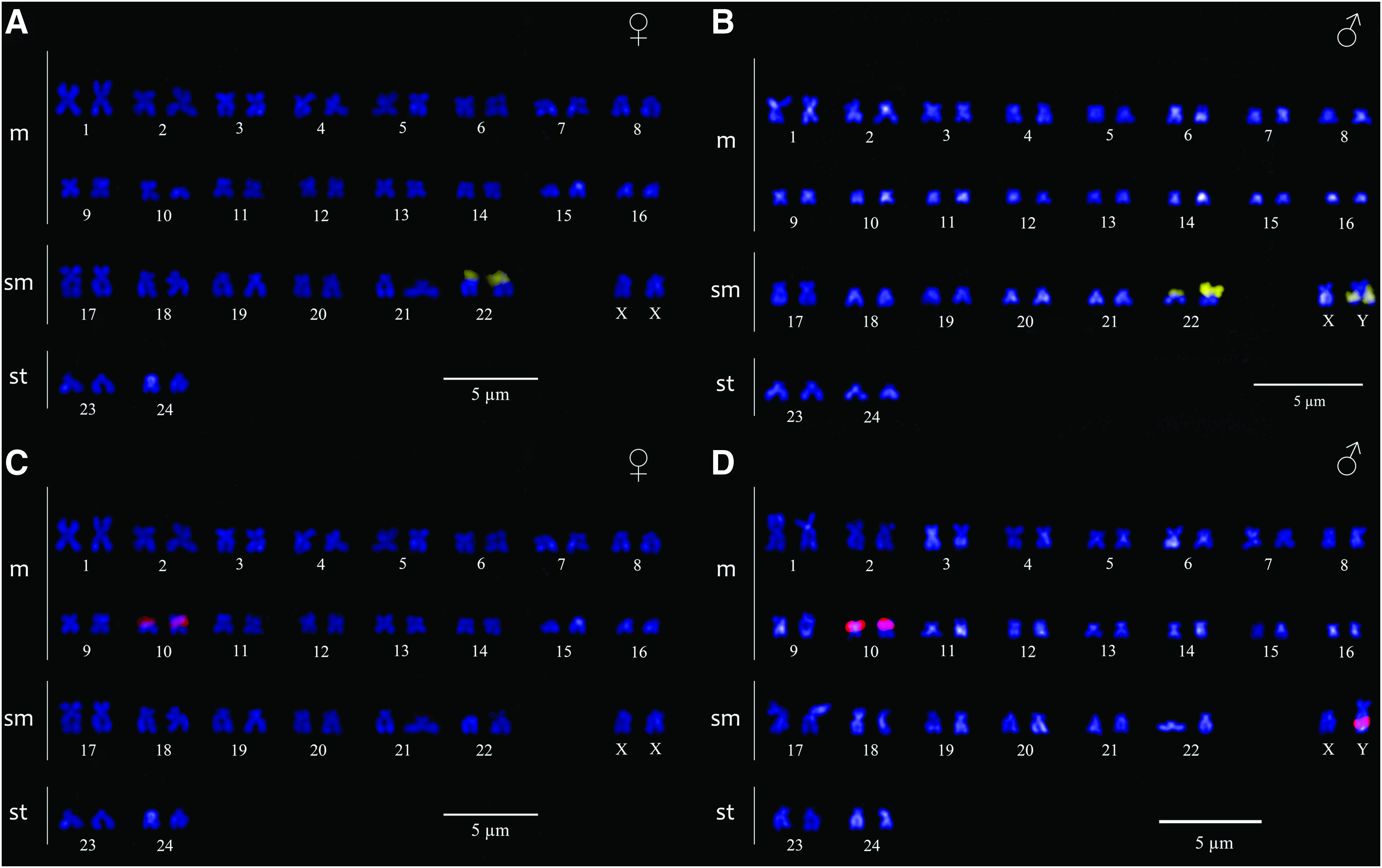
FISH pattern of P. cf. chagresi with 18S rDNA
The Y chromosome also revealed the largest accumulation of regions complementary to the microsatellite (GA)15 and (CA)15 probes on its long arm, contrasting with the patterns observed in the autosomes, where these regions were preferentially localized in the subtelomeric regions (Fig. 4).
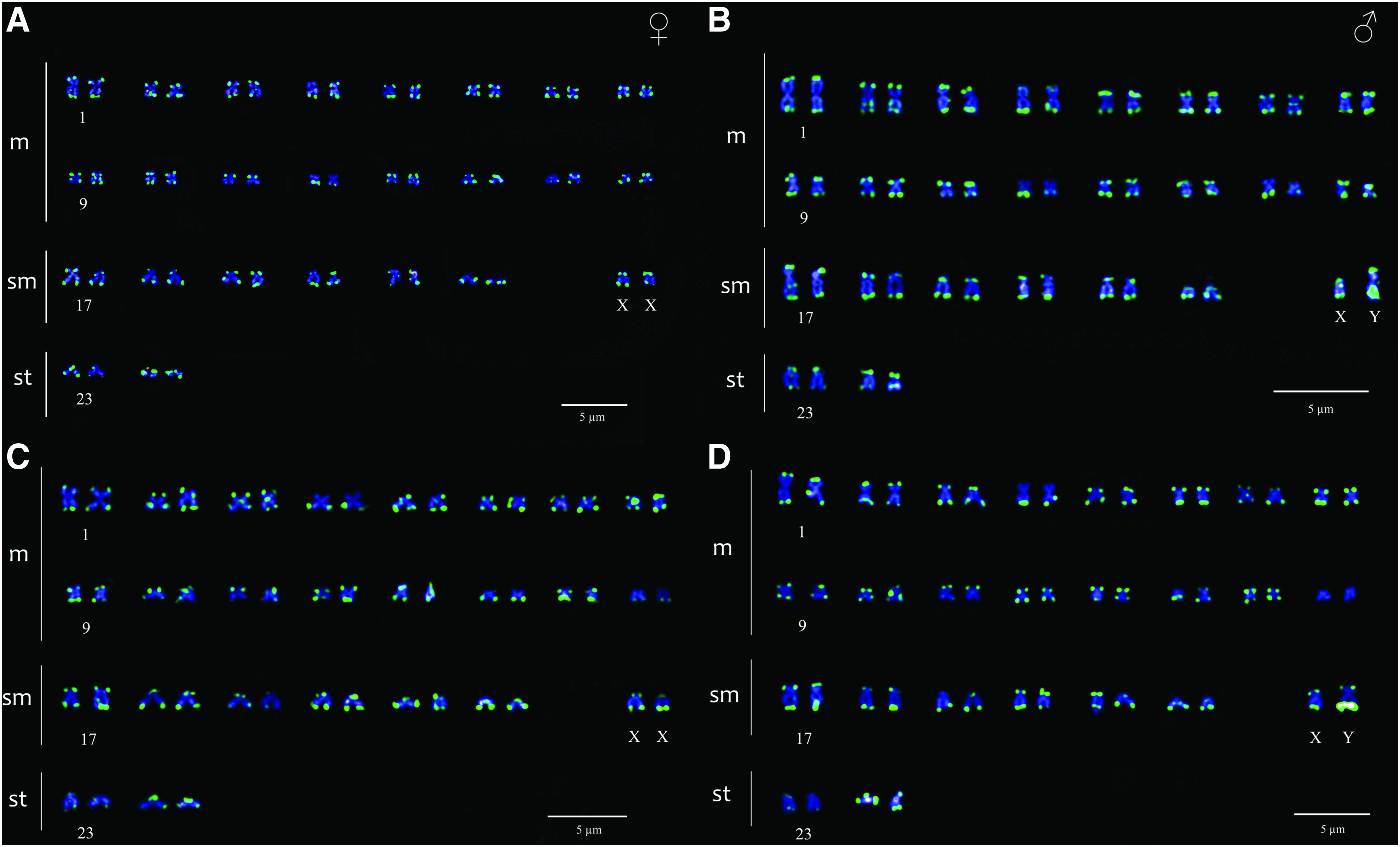
FISH pattern of P. cf. chagresi obtained with microsatellites GA(15)
Discussion
This work represents the first cytogenetic study of the Pimelodella genus in the trans-Andean region. Most studies on this genus have been focused in the cis-Andean region,7,10–15 where the diploid numbers are more frequently 2n = 46 (14 populations), whereas 2n = 52 and 2n = 58 are less common (reported in 3 and 4 populations, respectively)6–13,48–50 (Table 1). The diploid 2n = 58 number has been proposed as the plesiomorphic character state of Heptapteridae; subsequent chromosome number reduction would be the result of chromosome fusion 8 ; however, the hypothesis that the diploid number within Siluriformes could be a result of parallelism and convergence remains unresolved. 51 Although 2n = 50 in P. cf. chagresi is a novelty, this species shares other common features of Pimelodella species and other Heptapteridae, such as the predominance of metacentric and submetacentric chromosomes, and C-band blocks preferentially distributed at pericentromeric regions and on the active nucleolar organizer regions (NORs) 8 (Table 1).
2n, Diploid number; a, acrocentric; FN, fundamental number; m, metacentric, NOR, nucleolar organizer regions; rDNA, recombinant DNA; sm, submetacentric; st, subtelocentric.
Although differentiated sex chromosomes into Siluriformes are rare, a XX/XY sex chromosome system has been previously reported in Pimelodella sp. from the Upper Paraná Basin, where the heteromorphic sexual pair was described as a medium-sized submetacentric X chromosome and a small metacentric Y chromosome. 20 In P. boschmai, the sex chromosome pair is represented by a pair of metacentrics, where the Y chromosome is larger than its homologue. 7 However, in P. cf. chagresi, the sex chromosomes are submetacentric elements, and the large Y chromosome is associated with the accumulation of rRNA multigene families (18S and 5S rDNA) and microsatellite sequences (CA15 and GA15).
In the XX/XY and ZZ/ZW systems, the heteromorphic chromosome (Y or W) is generally smaller than its homologue, and is often degenerate or entirely heterochromatic. 24 Our results revealed that Y chromosome differentiation in P. cf. chagresi also involved heterochromatinization, but resulted instead in a large Y chromosome. Heterochromatin accumulation has also been reported in heteromorphic W sex chromosomes in other otophysan fish genera such as Leporinus and Triportheus,27,28,52,53 indicating that heterochromatinization has also been involved in the evolution of some ZZ/ZW sex chromosome systems. Although only 17 Pimelodella species out of 71 have been karyotyped,6,7,9–13 our results suggest that diploid numbers and sex chromosome systems may be good characters to indicate phylogenetic relatedness of species groups within this genus.
Autosomal banding patterns of (CA)15 and (GA)15 microsatellites DNA in P. cf. chagresi follow the general pattern of microsatellites in other fish, with predominant accumulation in subtelomeric and centromeric regions,27,28,54 However, it has been reported that accumulation in sex chromosomes show high levels of intrageneric variation, suggesting different evolutionary dynamics for these chromosomes among species. 28 Microsatellite DNA represents a highly dynamic component and is distributed ubiquitously throughout the genome, 55 suggesting these elements may be early sex chromosome colonizers, contributing significantly to the processes of differentiation and heterochromatinization. 27 The accumulation of repetitive sequences may be the result of different transposition and amplification events in the vicinity of sex-determining region 33 ; in this condition, microsatellites play an important role in the sex chromosomes' evolution as a consequence of their high ability to adopt unusual and varied DNA conformations. 24
The syntenic condition of 5S and 18S rDNAs on the Y chromosome in P. cf chagresi, represents an unusual pattern not previously reported in sex chromosomes of fish. rRNA multigene families are generally located in distinct genome regions 56 and in teleosts, and the general pattern presented by the vast majority of species studied is a single rDNA site of each multigene family.57,58 So far, the syntenic condition in the mapping of these genes has been reported only in autosomal chromosomes,36,59–61 being hypothesized as a possible ancestral condition in fish.62,63
The presence of rRNA genes in sex chromosomes of fish has also been reported in other species. In Oncorhynchus mykiss, 5S rRNA genes are localized in the partially undifferentiated X chromosome. 33 In Erythrinidae, the presence of these repetitive sequences in sex chromosomes shows variable patterns; in Hoplias malabaricus the presence of 18S rRNA genes coincides with an active NOR that was detected on the X chromosome; 34 whereas in Erythrinus erythrinus, Y chromosome differentiation evolved associated with 5S rRNA genes and TEs. 31 In other groups, the presence of 18S rRNA genes on the W chromosome is shared among species, such as Triportheus.35,36
It has been argued that the evolutionary process and organization of rDNA repeated units are subject to homogenization processes by factors, such as duplication and deletion, unequal crossover, or gene conversion.64–66 However, these processes may be counterbalanced by other mechanisms, such as the creation of new genes by duplication.65,67 The insertion of a genomic segment derived from a duplication event can also initiate the evolutionary process of sex chromosome differentiation. 68 Thus, the considerable accumulation of different domains of repetitive DNAs on the Y chromosome in P. cf. chagresi, could indicate different process of translocation or transposition in the differentiation of this element, which have been retained by the suppression of recombination in this area, 23 thus influencing the evolutionary transition from homologous to heteromorphic sex chromosomes.
In our results, we observed that the presence of 18S rDNA signal on the Y chromosome does not represent an active NOR region, which could suggest the presence of pseudogenes.60,69 Additionally, the presence of a third signal of 5S rRNA genes on this chromosome, in which suppression of recombination can cause genetic degeneration or accumulate loss-of-function mutations without reduced fitness, 21 suggests that phylogenetic perspectives should be carefully addressed because the clustering of rRNA genes seems to reflect their particular evolutionary pathway and not the evolutionary history of taxa, as has been proposed by Nakajima et al. 70 If this is true, rRNA genes would evolve faster than species cladogenesis; this interesting hypothesis may be tested when species phylogenetically close to P. cf. chagresi are karyotyped.
This study has underlined the potential role of repetitive DNA sequences in sex chromosome differentiation. However, in addition to contributing additional information to the evolutionary framework of sex chromosomes in fish, it could also address future studies that include cytogenetic tools in population analysis of the genus Pimelodella in the trans- and in the cis-Andean regions, a group with a complex evolutionary history, 71 where the taxonomy is poorly understood and the identity of species has not been resolved. Yet, P. cf. chagresi, together with other trans-Andean taxa, may in fact represent an outstandingly unique phylogenetic unit within the Neotropical region, as has been reported for many other species in the continent. 72
Footnotes
Acknowledgments
This study was supported by “Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES),” Brazil. The authors thank Marina Cunha for contributions in cytogenetic techniques.
Disclosure Statement
No competing financial interests exist.