
Abstract
Abstract
Androgenesis is useful for induction of doubled haploids from male genetic resources and contributes to the restoration of individuals from cryopreserved sperm. Here, we determined the suitable conditions for egg in vitro preservation and the suitable dose of UV irradiation for genetic inactivation of the egg nucleus, and established an improved procedure for induction of androgenetic-doubled haploids in zebrafish. The suitable solution for egg preservation was evaluated by the fertilization rate using different types of solutions or conditions. Hank's solution with 0.5% bovine serum albumin (pH8.0) was suitable for the preservation of zebrafish eggs. In addition, we discovered an improvement of fertilization rates by temporal preservation of ovulated eggs in the suitable solution. UV irradiation of eggs at 50–75 mJ/cm2 induced haploid embryos. Microsatellite genotyping using eight loci revealed the paternity and homozygosity of the putative androgenetic doubled haploids. The yield rate of androgenetic doubled haploids, which were induced by UV irradiation and heat shock, ranged from 0.4% to 10.7%.
Introduction
A
The induction of androgenesis can be accomplished by two methods: one uses irradiation and the other, thermal shock. Androgenesis using egg irradiation is the conventional method, and requires the genetic inactivation of the egg nucleus. Conversely, androgenesis using only thermal shock is a new method. Androgenetic doubled haploids were induced by cold shock of just-fertilized eggs, which were then heat shocked to double the chromosome set. 3
In the conventional method, androgenetic doubled haploids can be induced by three processes: (1) short-term storage of ovulated eggs in vitro, (2) inactivation of the egg nucleus, and (3) recovery of diploidy.
Ovulated eggs must be preserved in the proper solution during inactivation of the maternal genome, because the eggs must be prevented from drying out and being inactivated until fertilization. When ovulated eggs are released into the water, the eggs are immediately activated, form the perivitelline space, and lose fertilizing capacity. Therefore, we must use suitable solution for the species when eggs are preserved before fertilization. In previous studies, ovulated eggs of fish were stored in coelomic fluid, ovarian fluid, and artificial solution. In zebrafish (Danio rerio), ovarian fluid of rainbow trout (Oncorhynchus mykiss), 4 coho salmon (O. kisutch), 5 and Hank's solution 6 were used for temporary preservation of eggs. However, it is still unknown which solution is the best medium to keep ovulated eggs inactivated.
In zebrafish, androgenetic haploids have been induced through irradiation with γ-rays, 7 X-rays, 5 and UV light. 1 In particular, androgenesis using UV irradiation is convenient because special facilities and large-scale equipment are not required. However, suitable doses of UV irradiation have not been demonstrated in previous studies. Moreover, the best procedure to induce androgenesis in zebrafish has not been determined.
It is important to evaluate the suitable dose of UV irradiation to inactivate the egg nucleus completely, and to reduce the side effects of UV irradiation on embryogenesis. Residual maternal genomic substances caused by insufficient UV irradiation might result in the expression of unexpected maternal traits and aneuploidy. In previous studies, aneuploid larvae and chromosome fragments were detected when androgenesis was induced by lower than the suitable UV dose.8–10 UV irradiation of ovulated eggs influences not only the egg nucleus, but also substance in the egg cytoplasm and the yolk. Larvae with a cell mass without a body axis appeared at the higher UV doses in zebrafish 1 and muskellunge (Esox masquinongy). 11
The individuals induced by androgenesis using haploid sperm exhibit abnormalities characterized as “haploid syndrome,” resulting in inviability. However, androgenetic haploid can recover diploidy and viability by suppression of the first cleavage.
In this study, we report the successful induction of androgenetic doubled haploids by UV irradiation and heat shock treatment. First, we determined the suitable conditions for preservation to keep eggs inactivated and to achieve higher fertilization rates. In addition, we developed a method to improve the fertilization rate in artificial fertilization of zebrafish. Next, we evaluated suitable doses of UV irradiation for genetic inactivation of the egg nucleus based on the fertilization rate, survival rate, external appearance, and ploidy status of the larvae. Finally, we established the improved procedure for induction of androgenetic doubled haploids in zebrafish.
Materials and Methods
Egg and sperm collection
Wild-type strain and golden strain zebrafish were reared in our laboratory, maintained under a 14-h light and 10-h dark photoperiod at 28.5°C. Eggs were collected by squeezing and they were placed in a preservation solution. Sperm were collected by micropipette and diluted with Kurokura solution. 11
Evaluation of the type of preservation solution
Hank's solution with 0.5% bovine serum albumin (BSA) (pH 8.0), rainbow trout ovarian fluid (RTOF; pH 8.0), and fetal bovine serum (FBS; pH 7.2) were tested for the preservation of ovulated eggs. The eggs were placed into each solution and preserved for 10 or 30 min at 23°C. All experiments were performed in triplicate using different females.
Evaluation of the pH of the preservation solution
Various pH levels (pH 4.0, 7.0, 8.0, 9.0, and 12.0) of Hank's solution with 0.5% BSA were prepared to determine the suitable pH for ovulated egg preservation. Eggs were placed directly into Hank's solution at various pH levels and preserved for 30 min at 23°C. All experiments were performed five times using different females.
Evaluation of the condition of the temporary preservation
The effect of agitation in the preservation solution on the fertilization capacity of ovulated eggs was evaluated because we intended to perform genetic inactivation of the egg nucleus with agitation to disperse eggs uniformly. Eggs were directly placed into Hank's solution with 0.5% BSA (pH 8.0) and preserved under either agitating or static conditions for 5, 10, and 30 min at 23°C. Under the agitating condition, the dishes containing the eggs were horizontally mixed at 60 rpm on a shaker (VS-2, SAKURA SEIKI Co. Tokyo, Japan) during preservation. All experiments were performed five times using different females.
Genetic inactivation of egg nucleus
Genetic inactivation of the egg nucleus was performed by UV irradiation of the ovulated eggs. The ovulated eggs were placed in the center of a chamber. The chamber was prepared as previously described. 10 Ovulated eggs were placed into Hank's solution with 0.5% BSA (pH 8.0). The chamber was filled with 2.4 mL Hank's solution with 0.5% BSA, and the eggs were uniformly dispersed to avoid overlap. UV irradiation was carried out with horizontal agitation at 60 rpm using the shaker.
As preliminary experiments, the eggs were UV irradiated for 0, 25, 75, 150, and 200 s at 1.0 mW/cm2, which correspond to 0, 25, 75, 150, and 200 mJ/cm2, respectively. In next evaluation of the suitable UV dose, the eggs were UV irradiated for 0, 50, 75, 100, 125, and 150 s at 1.0 mW/cm2, which correspond to 0, 50, 75, 100, 125, and 150 mJ/cm2, respectively. The UV irradiated eggs were transferred to a new dish filled with Hank's solution without BSA and washed. The eggs were moved to new dry dish. Sperm from the golden strain was added to the irradiated eggs, mixed well, and activated by adding 0.2 mL water at 28.5°C. Preliminary experiment tested five doses was performed five times using different females. Evaluation of the suitable UV dose tested six doses was performed six times using different females.
Induction of androgenetic doubled haploids by heat shock treatment
Androgenetic doubled haploids were induced by inhibition of the first mitotic division by heat shock treatment as previously described.5,12 Ovulated eggs from one female were divided into two pools. The eggs from first pool were fertilized with sperm from the golden strain without any treatment (control group). The eggs from second pool were UV irradiated for 75 s at 1.0 mW/cm2 and fertilized with sperm from the golden strain (UV irradiation group). The eggs were incubated at 28.5°C after fertilization. At 13 min after fertilization, the part of eggs from UV irradiation group were transferred to a heat shock container (the bottom of a 320 mL volume plastic box was removed and covered with mesh), which was placed in a water bath at 41.4°C, and incubated for 2 min. After the heat shock treatment, the container was moved back to a larger container filled with water at 28.5°C. Fifteen minutes after incubation in the larger container, the eggs were returned to a plastic dish using a pipette and incubated at 28.5°C. All experiments were performed in triplicate using different females.
Fertilization rate, survival rate, and external appearance
In evaluation of the suitable condition for egg preservation and the suitable UV dose, we measured fertilization rate and survival rate and observed the external appearance in the control and all the experimental groups. The fertilization rate was calculated as the number of cleaved eggs relative to the total number of eggs at the 8–128 cell stage. The survival rate was calculated by the number of surviving embryos relative to the number of cleaved eggs at 12, 24, and 48 hour postfertilization (hpf). External appearance was observed at 12 hpf (gastrulation), 24 hpf (somite stage), and 48 hpf (hatching stage) using a stereomicroscope (MZ16F-RCFL, LEICA). In induction of doubled haploids, the survival rate and external appearance were examined at 72 hpf. The developmental stage of zebrafish was determined according to a previous study. 13
All surviving embryos containing normal and abnormal appearance were counted when calculating the survival rate. “Abnormal” embryo that might not grow into adult was also classified to “surviving” embryo if it survived at each stage. Embryos and hatched larvae exhibited abnormalities like edema, ascites, dwarfing, or curved body axis were classified as “abnormal.” Embryos and hatched larvae without these abnormalities were classified as “normal.”
Ploidy and paternity analysis
The relative DNA content of hatched larvae and live embryos were measured by flow cytometry (CyFlow; Partec GmbH, Münster, Germany), according to a previously described procedure. 10 In evaluation of the suitable UV dose, each live embryo at 48 hpf was incubated in 100 μL of solution A (CyStain DNA 2step, Partec GmbH) that extract nuclei for 20 min. Then, 50 μL of the solution was mixed with 400 μL of solution B (CyStain DNA 2step; Partec GmbH) that contain 4′, 6-diamidino-2-phenylindole and stain extracted nuclei, and analyzed by flow cytometry.
In induction of doubled haploids, each hatched larva at 72 hpf was incubated by 50 μL of solution A for 20 min. Then, 10 μL of the solution was mixed with 300 μL of solution B and analyzed by flow cytometry. The remaining 40 μL of the solution was used for DNA extraction for microsatellite genotyping. Six diploids appeared in the control group, six haploids appeared in the androgenesis group, and 20 putative doubled haploids appeared in the heat shock group, and each set of parents were genetically analyzed using eight microsatellite loci 14 : Z8874, Z644, Z8495, Z7576, Z6622, Z992, Z6010, and Z5265 (batch A); Z8874, Z644, Z8495, Z10215, Z7576, Z9331, Z9708, and Z5265 (batch B); Z1781, Z8874, Z8495, Z7576, Z992, Z6010, Z9331, and Z9708 (batch C). DNA extraction and microsatellite genotyping were performed according to a previously described procedure. 3 Microsatellite genotyping was conducted using the sequencer ABI PRISM 3130xl (Applied Biosystems). The data for microsatellite genotyping were analyzed using Gene Mapper software v3.7 (Applied Biosystems).
Statistical analysis
The data for evaluation of the suitable condition for egg preservation and suitable UV dose are shown as mean ± standard error, and were analyzed with one-way analysis of variance, followed by Tukey's test (p < 0.05).
Results
Evaluation of the type of preservation solution
To determine the suitable solution for egg preservation, three types of solutions, Hank's solution with 0.5% BSA, RTOF, and FBS, were tested with two preservation periods (Table 1). The relative fertilization rate of eggs in Hank's solution with 0.5% BSA was higher than that of the other preservation groups, regardless of preservation time. Moreover, in Hank's solution with 0.5% BSA for 10 and 30 min, the relative fertilization rate was improved when compared with that of the control group (no egg preservation). The survival rate in all preservation groups at 48 hpf was not significantly different from that of the control group.
Each value is the mean ± SE of the triplicate.
Different alphabet superscripts in each column indicates significant differences as determined by one-way ANOVA and Tukey's test (p < 0.05).
ANOVA, analysis of variance; BSA, bovine serum albumin; hpf, hour post fertilization; SE, standard error.
Evaluation of the pH of the preservation solution
To choose the suitable pH for egg preservation, five levels of pH (pH 4.0, 7.0, 8.0, 9.0, and 12.0) of Hank's solution with 0.5% BSA were tested (Table 2). The relative fertilization rate of the pH 8.0 group was higher than that of the other preservation groups. There were no cleaved eggs in the pH 4.0 and pH 12.0 groups because all eggs were activated before artificial insemination. The relative fertilization rate in pH 4.0, 9.0, and 12.0 groups was significantly lower than that of the control group (no egg preservation). The survival rate in pH 7.0 and 8.0 groups at 48 hpf was higher than that of the pH 9.0 group.
Each value is the mean ± SE of five replicates.
Different alphabet superscripts in each column indicates significant differences as determined by one-way ANOVA and Tukey's test (p < 0.05).
Evaluation of the condition of the temporary preservation
To determine the effects of preservation conditions of ovulated eggs on fertilization capacity, agitation, and static conditions were tested in three preservation periods (Table 3). In each agitation preservation group, the relative fertilization rate was higher than that of the static preservation groups. Particularly, the relative fertilization rate of agitation preservation for 5 and 10 min groups was significantly improved when compared with that of the control group (no egg preservation). In all preservation groups, the survival rate at 48 hpf was higher than that of the control group. However, there was no statistically significant difference between the control and preservation groups in survival rate at 48 hpf.
Each value is the mean ± SE of five replicates.
Different alphabet superscripts in each column indicates significant differences as determined by one-way ANOVA and Tukey's test (p < 0.05).
Evaluation of the suitable UV dose
First, to conduct preliminary experiments of UV irradiation for genetic inactivation of the egg nucleus, five doses (0, 25, 75, 150, and 200 mJ/cm2) were tested. The fertilization rate and survival rate of the control and all UV irradiated groups were shown in Table 4. In the 25–200 mJ/cm2 groups, the fertilization rate gradually decreased with increasing UV dose. In the 150 and 200 mJ/cm2 groups, the fertilization rate was lower than that of the control group. Conversely, the fertilization rates in 25 and 75 mJ/cm2 groups were higher than that of the control group, despite UV irradiation treatment. In addition, the fertilization rate of the 25 mJ/cm2 group was significantly higher than that of the 200 mJ/cm2 group.
Each value is the mean ± SE of five replicates.
Different alphabet superscripts in each column indicates significant differences as determined by one-way ANOVA and Tukey's test (p < 0.05).
The survival rates of all UV irradiated groups were lower than that of the control group. In the 25–200 mJ/cm2 groups, the survival rate at 12, 24, and 48 hpf gradually decreased with increasing UV dose. The survival rate at 12 hpf in the 150 and 200 mJ/cm2 groups, at 24 hpf in the 75–200 mJ/cm2 groups, and at 48 hpf in the 25–200 mJ/cm2 groups were significantly lower than that of the control group at each time point. In particular, in the 200 mJ/cm2 group, only two individuals (1.3% ± 1.30%) survived at 48 hpf among all five trials.
The relative DNA contents of surviving embryos from all UV irradiated groups were measured by flow cytometry. Diploid embryos of the intact controls were used as the standard diploid (2C) DNA content. Haploidy was detected in all UV irradiated groups (Table 5). In the 75 and 200 mJ/cm2 groups, haploid embryos only were detected. Out of 384 surviving embryos, six normal diploid embryos appeared in the 25 mJ/cm2 group. The proportion of haploid embryos was 87.5% and 83.0% in the 25 and 150 mJ/cm2 groups, respectively.
Each value is the total of five replicates.
1.0C–1.1C.
Two 1.1C, two 1.2C, one 1.8C.
One 1.7C.
FCM, flow cytometry.
Next, to investigate the suitable UV dose for genetic inactivation of the egg nucleus, six doses (0, 50, 75, 100, 125, and 200 mJ/cm2) were tested. The fertilization rates of control group and all UV-irradiated group are shown in Table 6. In the 50–150 mJ/cm2 groups, the fertilization rate gradually decreased with increasing UV dose, except in the 125 mJ/cm2 group. In the 100–150 mJ/cm2 groups, the fertilization rate was lower than that of the control group. In contrast, the fertilization rates in 50 and 75 mJ/cm2 groups were higher than that of the control group, despite UV irradiation treatment. Moreover, the fertilization rate of the 50 mJ/cm2 was significantly higher than that of the 150 mJ/cm2 group.
“No. of eggs used,” “Fertilization,” and “Embryo survived” are the mean ± SE of six replicates.
“Haploid syndrome,” “Incomplete epiboly,” and “Cell mass” are the mean ± SE of the triplicate.
Different alphabet superscripts in each column indicates significant differences as determined by one-way ANOVA and Tukey's test (p < 0.05).
The survival rate of the control and all UV irradiated groups are also shown in Table 6. The survival rates of all UV irradiated groups were lower than that of each control group. In the UV irradiated groups, the survival rate at 12, 24, and 48 hpf gradually decreased with increasing UV dose. The survival rate at 12 hpf in the 125 and 150 mJ/cm2 groups, at 24 hpf in the 75–150 mJ/cm2 groups, and at 48 hpf in the 50–150 mJ/cm2 groups were significantly lower than that of the control group at each time point.
External appearance of the control group and all UV irradiated groups were observed at 12, 24, and 48 hpf. Representative embryos in each experimental group are shown in Figure 1. At 12 hpf, embryonic development in the UV irradiated groups did not complete epiboly although the control embryos reached the first somite stage (Fig. 1a–f). In 50 and 75 mJ/cm2 groups, embryos reached 80%–90% epiboly stage. However, in 100 mJ/cm2 group, embryos were at 60%–80% epiboly stage. In 125 and 150 groups, embryos only reached before 50% epiboly stage. The delay was progressively noticeable among the UV irradiated groups with increasing UV dose. At 24 hpf in the control group, the yolk extended toward the tail (Fig. 1g), but in the 150 mJ/cm2 group, the yolk did not extend in that direction (Fig. 1l). In the 50–125 mJ/cm2 groups, yolk extension was observed in some embryos (Fig. 1h–k).
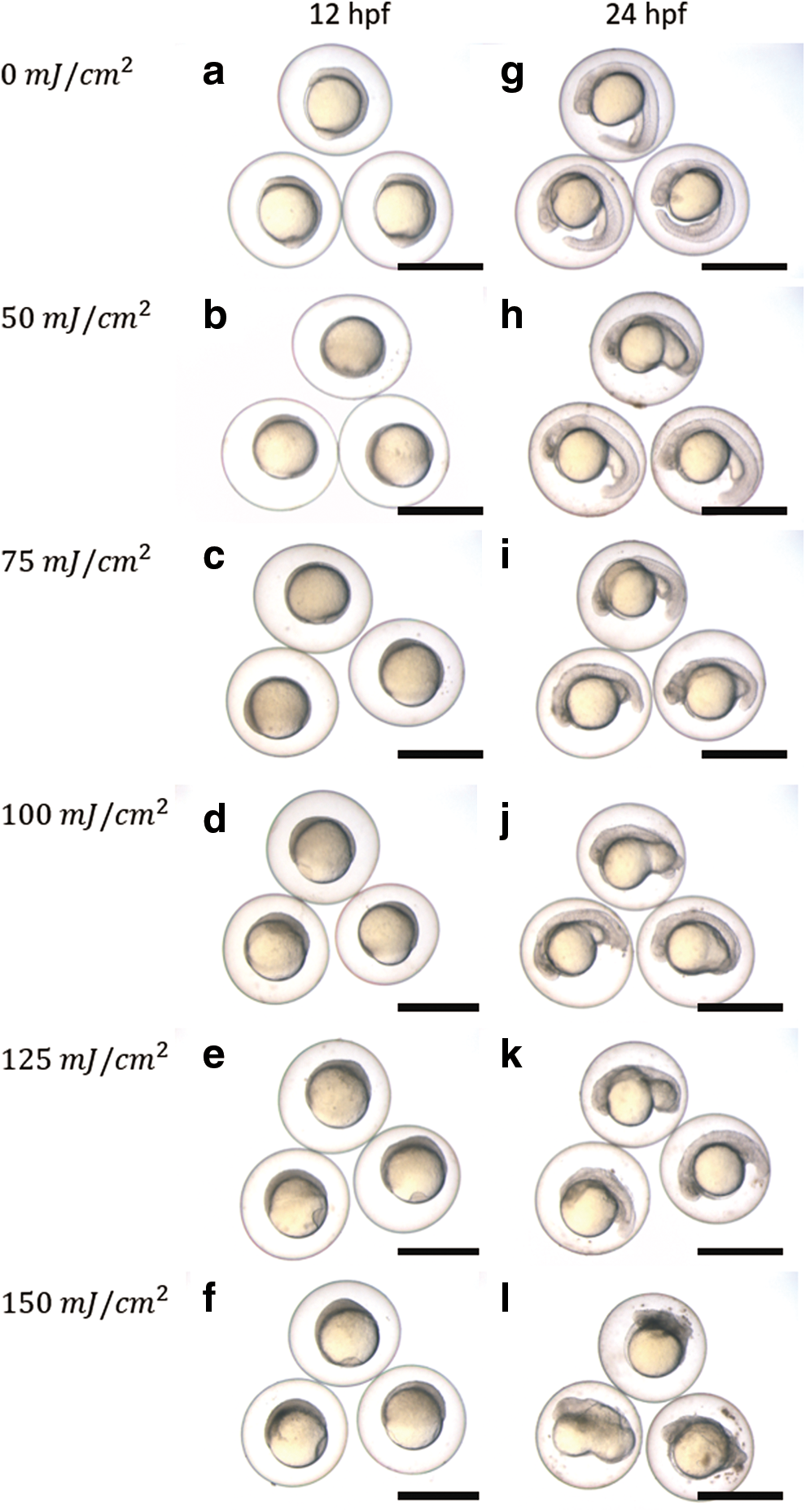
External appearance of 12
At 48 hpf in the control group, embryos had normal appearance (Fig. 2a). Conversely, abnormalities characterized as those of “haploid syndrome” (edema, ascites, dwarfing, microcephaly, and microphthalmia) were observed in embryos from the UV irradiation eggs (Fig. 2b). Most of the abnormal embryos in all UV-irradiated groups exhibited “haploid syndrome” (Table 6). However, in the 100–150 mJ/cm2 groups, abnormal embryos showed severe abnormalities such as “incomplete epiboly” and “cell mass” (Fig. 2c, d). In particular, abnormal embryos classified as “incomplete epiboly” were observed in the 100–150 mJ/cm2 groups (8.3% ± 8.33% to 21.8% ± 9.96%). In addition, the proportion of “cell mass” embryos in the 125 and 150 mJ/cm2 groups (13.0% ± 8.07% and 12.2% ± 6.19%, respectively) was higher than in the other groups (4.2% ± 4.17% to 7.3% ± 6.35%). The proportion of “haploid syndrome,” “incomplete epiboly,” and “cell mass” embryos were calculated using data from three trials.

External appearance of embryos at 48 hpf in evaluation of the suitable UV dose.
The relative DNA content of surviving embryos from all UV irradiated groups was measured by flow cytometry. Diploid embryos of intact controls were used as the standard diploid (2C) relative DNA content. Haploidy was detected from all UV irradiated groups (Table 7). The proportion of haploid embryos was more than 90% in all UV irradiated groups. Out of 134 surviving embryos, 1 normal diploid embryo appeared in the 75 mJ/cm2 group. In the UV irradiated groups, a few aneuploids and diploids were also detected.
Each value is the total of six replicates.
Four 1.1C, one 1.8C.
One 1.1C.
Three 1.1C.
Two 1.1C.
One 2.1C.
Induction of doubled haploids
We attempted to induce androgenetic doubled haploids by inhibition of the first mitotic division by heat shock treatment. The three batches (A–C) of developmental status and ploidy status are shown in Table 8. The fertilization rate in the UV irradiation and heat shock groups ranged from 32.7% to 95.4%. The survival rate at 72 hpf of the UV irradiation and heat shock group, which ranged from 0.4% to 19.3%, was lower than that of the control groups (74.0% to 90.9%).
External appearance of hatched larvae at 72 hpf for all experimental groups is shown in Figure 3. In the control group, embryos had a normal appearance (Fig. 3a). Conversely, most hatched larvae from the UV-irradiation group exhibited abnormalities characteristic of haploid syndrome (Fig. 3b). In the UV irradiation and heat shock group, most larvae had an abnormal appearance (Fig. 3d). There were six larvae with normal appearance only in batch B (Fig. 3c, Table 8), similar to that of control larvae. Larva in Figure 3c exhibited the golden phenotype with lighter pigmentation pattern around yolk when compared to larva from control group in Figure 3a. These six normal larvae (classified at 72 hpf) did not show swimming up and feeding, and resulted in inviability until 11 dpf presumably due to the lack of swim bladder inflation. Diploid individuals were detected in each batch (Table 8).
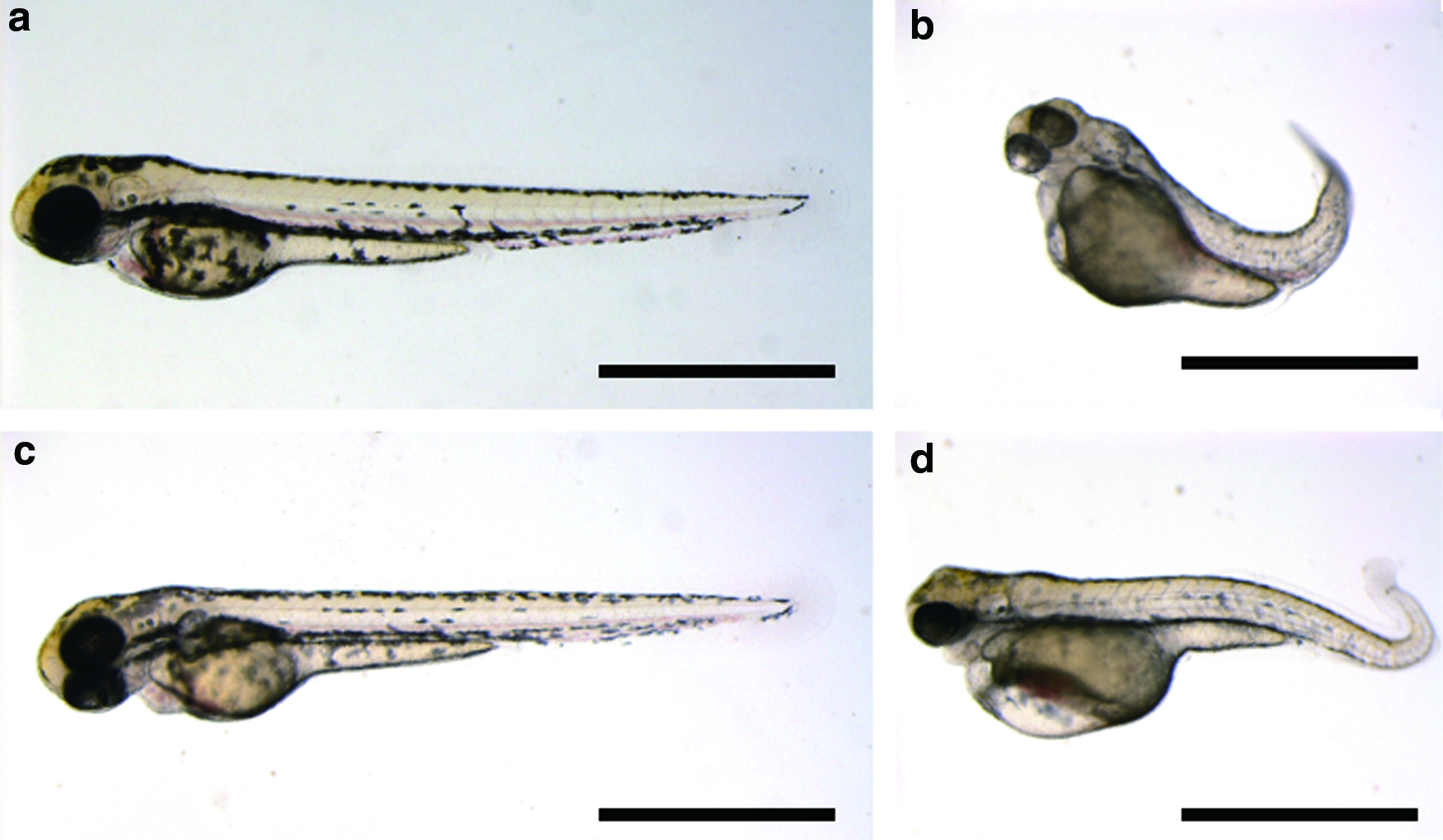
External appearance of hatched larvae at 72 hpf in induction of doubled haploids.
The paternity and homozygosity of 20 putative androgenetic doubled haploids that appeared from the three batches were determined by genetic analysis using eight microsatellite loci of zebrafish. 15 For all of the eight loci tested, only paternal alleles were transmitted to the androgenetic haploid and androgenetic doubled haploid progeny, and no maternally derived alleles were observed (Tables 9–11). For example, at locus Z9331 from linkage group 19 of Batch B, the male genotype was 291/293, and the female was 277/280. Ten out of 16 diploid progeny had homozygous 291/291 genotypes, whereas 6 had homozygous 293/293 genotypes. The 20 diploid progeny that appeared in the heat shock group were homozygous in the 8 loci tested.
Asterisk indicates inconclusive loci because the male and female shared the same allele size.
Asterisk indicates inconclusive loci because the male and female shared the same allele size.
Asterisk indicates inconclusive loci because the male and female shared the same allele size.
Discussion
Temporal preservation of zebrafish ovulated eggs
Preservation in Hank's solution with 0.5% BSA at pH 8.0 resulted in the highest fertilization rate of the three solutions when compared three types of preservation solution. In a previous study, Hank's solution 6 and salmonid ovarian fluid4,5 were primarily used for temporary preservation of zebrafish eggs. In this study, it was suggested that Hank's solution with 0.5% BSA is better than salmonid ovarian fluid to preserve zebrafish eggs. In addition, although FBS was not previously used for egg preservation, the fertilization rate of the FBS preservation group was not significantly different from that of the RTOF group. Thus, it might be possible for zebrafish ovulated eggs to be kept in FBS before artificial insemination.
When we evaluated the suitable pH for egg preservation, preservation in the pH 8.0 group resulted in a higher fertilization rate than did the other pH groups. There were no cleaved eggs in the pH 4.0 and 12.0 groups because all eggs were already activated before artificial insemination. These results indicated that the pH of the preservation solution has an important role in maintaining egg fertility. We therefore concluded that Hank's solution should be prepared at pH 8.0 to preserve zebrafish ovulated eggs.
Considering the fertilizing environment, pH and protein concentration of the ovarian fluid were correlated with the fertilization rate in common carp Cyprinus carpio. 16 The pH of ovarian fluid of high-quality egg batches with more than 70% fertilized eggs ranged from pH 8.35 to 8.85. This suggests that ovarian fluid around the eggs influences fertilization. Moreover, it was demonstrated that the pH of ovarian fluid also influences sperm motility and duration in rainbow trout. 15 Several sperm samples of rainbow trout were not activated after dilution with 400 times amount of ovarian fluid that ranged from pH 7.29 to 7.67. In rainbow trout, along with an increase in pH, increased duration and motility of sperm were observed. From the results of this study, it is speculated that the pH of the preservation solution could also improve sperm motility in the fertilizing environment.
Next, we investigated the effect of agitation during preservation on fertilization rate. In each agitation group, the relative fertilization rate was higher than that of the static group. This result indicated that there was no physical damage to eggs resulting from agitation. In addition, it was demonstrated that the agitation treatment during egg preservation increased the fertilization rate. In a previous study, mechanical agitation of eggs in ovarian fluid did not affect hatching and normal embryo rate in common carp. 17 These results indicated that washing the surface of the egg was effective in maintaining and improving egg fertility. However, egg preservation tests were not conducted more than 30 min in this study. It might be important to elucidate the maximum preservation period of ovulated eggs in Hank's solution.
Through this series of experiments, the fertilization rate increased compared with that of the control group by temporary preservation in Hank's solution with 0.5% BSA before fertilization. The phenomenon similar to this result was observed in the preservation of rainbow trout eggs.18,19 It was determined that short-term storage and/or washing in the artificial medium, “Cortland medium,” improved fertility and survival rate. These results suggest that storage of ovulated eggs in the suitable solution increases egg fertility.
In addition, it has been shown that there are sperm attractant glycoproteins that are present around the micropyle, and sperm activating proteins that are released from the unfertilized egg in the herring Clupea pallasii and black flounder Pleuronectes obscurus.20,21 It was also stated that the chorion around the micropyle of the bitterling Rhodeus ocellatus, which is in the same family of Cyprinidae as the zebrafish, has a low-molecular weight substance that activates and aggregates sperm. 22 According to the present study, temporary preservation in Hank's solution makes good effect on the ability of substances, such as glycoproteins related to sperm attraction, although it has not known whether that substance exists in the zebrafish eggs. In the other words, improvement of the fertilization rate in the Hank's solution preservation group was caused by an increase of sperm attraction activity following preservation in Hank's solution with 0.5% BSA at pH 8.0. It could be hypothesized that removing the ovarian fluid residue from zebrafish eggs and preservation in Hank's solution improved the fertility because of a desirable egg surface for artificial fertilization.
Suitable dose of UV irradiation
Through evaluation of the suitable UV dose, the fertilization rate dramatically decreased in the 150 and 200 mJ/cm2 group as compared with that of the control group, but there was no significantly difference among the 0–125 mJ/cm2 groups. Therefore, it is suggested that UV irradiation over 150 mJ/cm2 resulted in a reduction in fertilization rate. Conversely, in the 25–75 mJ/cm2 groups, the fertilization rate exceeded that of the control group (0 mJ/cm2). This improvement might be an influence of preservation in Hank's solution with 0.5% BSA during the UV irradiation treatment.
The flow cytometry analysis showed that haploid embryos appeared in 25–200 mJ/cm2 UV dose groups. Therefore, doses of more than 25 mJ/cm2 of UV could inactivate the egg nucleus in zebrafish eggs. However, diploid normal (biparental) embryos and aneuploid embryos appeared in the 25 mJ/cm2 group. Furthermore, there were few surviving embryos in the 200 mJ/cm2 group. Thus, we concluded that less than 25 mJ/cm2 and more than 200 mJ/cm2 of UV were not the suitable dose for induction of androgenetic zebrafish. Out of 229 survival embryos, two haploid embryos appeared in control group might be caused by spontaneous gynogenesis or other types of parthenogenesis. However, because we do not have any evidence, we cannot demonstrate how these embryos were produced.
Next, to determine the suitable dose of UV, external appearance was observed in test with six doses (0–150 mJ/cm2). Most of the embryos resulting from UV irradiated groups exhibited an appearance characterized as haploid syndrome. 5 However, a few embryos from putative haploids exhibited additional abnormalities known as “incomplete epiboly” or “cell mass.”
Embryos characterized as “incomplete epiboly” were possibly induced by slow gastrulation. The developmental delay in the gastrulation was noticeably observed among the UV irradiated groups. “Incomplete epiboly” was observed only in higher UV doses (100–150 mJ/cm2) groups because speed of gastrulation was negatively correlated with UV dose. It was reported that the dose-dependent retardation of epiboly was observed in biparental zebrafish embryos with UV-irradiation at 30 mpf. 23 In these UV-treated embryos, abnormal length and array microtubules were observed in the yolk cytoplasmic layer. During epiboly, epibolic movements are driven by organized microtubules in the yolk cytoplasmic layer. 24
On the other hand, “cell mass” is a severe abnormality caused by aberrant embryonic axis formation. It is known that formation of a single body axis results from normal migration of the dorsal determinants that change the distribution from the vegetal pole to a marginal part of the blastoderm. 25 This transportation of substance is related to the organization of microtubules in the yolk. 26 It was also stated that UV irradiation of the embryo at early stages disrupted the formation of the vegetal microtubule array and caused the loss of the axis. In addition, embryos without apparent axis formation were observed in the induction of zebrafish using UV irradiation. 1 It was also reported that muskellunge eggs developed as a cell mass following twice the UV irradiation than is the suitable UV doses. 9
These results indicated that UV irradiation effects not only inactivation of the egg nucleus, but also microtubule organization in the yolk. Therefore, the abnormalities of “incomplete epiboly” and “cell mass” appeared in >100 mJ/cm2 groups because of excessive UV doses. We therefore concluded that doses of more than 100 mJ/cm2 of UV were excessive to inactivate the maternal genome and induce androgenetic zebrafish. In this study, suitable dose of UV irradiation was not evaluated by pigmentation phenotype because developmental delay occurred after gastrulation made it difficult to observe pigmentation patterns at regular developmental stage. Thus, we employed the fertilization rate, survival rate, ploidy analysis, and external appearance for suitable UV dose evaluation. In conclusion, the suitable UV dose for genetic inactivation of the maternal genome and induction of androgenetic zebrafish was 50–75 mJ/cm2.
Induction of doubled haploids
Diploid progeny that appeared from the UV irradiation and heat shock treatment group possessed only paternal alleles for all eight loci tested. This result indicated that it was possible to induce androgenetic doubled haploids by treatment with UV irradiation and first cleavage suppression. Thus, the diploid larvae appeared in each UV irradiation and heat shock group were considered androgenetic doubled haploids. The yield rate of androgenetic doubled haploids relative to the total number of eggs used ranged from 0.4% to 10.7%. Moreover, we succeeded in inducing six androgenetic diploids (0.5%) from 1267 irradiated eggs. In a previous study, it was reported that the yield rate of androgenetic doubled haploids by X-ray irradiation and heat shock treatment ranged from 1.3% to 2.1%. 5 In zebrafish, the generation of androgenetic doubled haploids was performed using only thermal shock. 3 The yield rate of normal androgenetic doubled haploids was 1.10% ± 0.19% in that study. 3 Although androgenetic doubled haploids have been induced in common carp, 17 loach, 27 and amago salmon (Oncorhynchus masou ishikawae), 28 the yield rates were quite low in these studies.
Androgenetic doubled haploids are valuable in the establishment clonal lines because it is expected that androgenetic doubled haploid fish produce genetically identical gametes. Therefore, clonal lines can be established in only two cycles of androgenesis. In the second cycle, it could be possible to produce genetically identical progenies by using the sperm of first-generation of androgenetic doubled haploids.
In conclusion, we established an improved method to generate androgenetic doubled haploids in zebrafish using UV irradiation and heat shock.
Footnotes
Acknowledgments
This work was financially supported in part by the Japan Society for the Promotion of Science (JSPS), KAKENHI grant no. 17J02645. We would like to acknowledge Dr. Miloš Havelka for his suggestion about this study and publication.
Disclosure Statement
No competing financial interests exist.