
Abstract
Abstract
The current analysis investigates whether the uplift of the Serra da Esperança and the Ponta Grossa Arch in the Serra Geral resulted in ichthyofaunistic changes in adjacent basins. For this, we describe the phylogeographic structure among populations of Trichomycterus collected in hydrographic basins in southern Brazil by using partial nucleotide sequences of the mitochondrial gene Cytochrome C Oxidase subunit I. Analyses revealed that the nomenclature Trichomycterus davisi fails to contain the whole genetic diversity range found in the collected specimens and indicates at least six genetic lineages in Trichomycterus. Diagnostic morphological characteristics not associated to T. davisi could be identified in some specimens from the Iguaçu Piquiri haplogroup, indicating the occurrence of species Trichomycterus stawiarski. The lack of morphological differences among the other clades clearly suggests a cryptic species case. Molecular analyses revealed at least five new species besides T. davisi in the hydrographic basins and support the interpretation that genetic structure in T. davisi species complex is explained by tectonic events intrinsic to the areas of influence of Serra da Esperança and the Ponta Grossa Arch which occurred around 1.7 My.
Introduction
T
These organisms have limited ability to disperse overland and need connections between river basins for dispersion; therefore, a strong relationship is expected between the history of the basins and the fish fauna.4,6 Watershed divides, such as high mountains, are often regarded as effective dispersal barriers for fish, and fish distributions are often used to assign hydrographic basin limits. 6 Smaller isolated basins are separated from large inland drainages by the scarped, mountainous landscapes on the eastern margin of the Brazilian mountain ranges that extend from southern to southeastern Brazil. 7
The Serra Geral (also called Serra da Esperança or Serra da Boa Esperança in Paraná State) is a Lower Cretaceous formation and extends over more than 1 million km2 in the Paraná subbasin of central and southern Brazil, Paraguay, and northeast Argentina. 8 It is included in the Atlantic Forest biome, one of the most threatened hotspots in the world. 9 Strugale et al. 10 analyzed the structure of the São Bento Group (Pirambóia, Botucatu and Serra Geral formations) using aerial photographs, satellite images, aeromagnetometric data, and digital terrain models to establish the structural framework and paleostress trends related to the evolution of the Ponta Grossa Arch. 11 This arch comprises an uplift of the crystalline basement along the southeastern portion of the Paraná Basin. 11 These two geological formations have been linked to both the diversification and mixing of the ichthyofauna between the adjacent river basins, suggesting vicariant processes and/or fauna interchange.7,12
Trichomycterus species occur in the main drainage basins of southern Brazil. Trichomycterus davisi and other species have a restricted distribution, 13 specifically in the Iguaçu River and Ribeira de Iguape River basins. 14 The type-locality of T. davisi is Serrinha, Iguaçu River basin, Balsa Nova, Paraná State.15,16 However, specimens of T. davisi were found in the Paranapanema River basin (Upper Paraná River basin), 17 while recent samples reported specimens with the morphological characteristics of T. davisi at various sites in the Ivaí, Piquiri, and Tibagi River basins (Upper Paraná River Basin).18–20
Although T. davisi has been identified by morphological and meristic characteristics, it presents a wide variation in color patterns throughout its ontogenetic development. Phenotypic variation impairs the identification of different sized specimens. According to de Pinna, 21 there are three main factors that cause taxonomic problems in Trichomycterus, namely, a long taxonomic history, paraphyletic groups, and incomplete knowledge of the true diversity of the species. Furthermore, the species' wide geographic distribution, mainly inhabiting stream sources and running water courses, makes studies on the genus highly difficult. Many species feature a highly endemic degree in restricted areas.22–24 Few studies have investigated the species' genetic composition. Cytogenetic studies have revealed great diversity in the amount of intra- and interindividual chromosomes in T. davisi of the Iguaçu River basin.25,26
Molecular markers of mitochondrial DNA have been used to identify the species boundaries, with special reference to gene Cytochrome C Oxidase subunit I (COI), the main marker used worldwide for animals in the Consortium for the Barcodes of Life. 27 Low rates of intraspecies genetic distances for COI are generally reported when compared to estimates of interspecific genetic distance.27,28 The tool is currently used in population studies and species identification,29–34 including cryptic species. 35 Recently, tools for the delimitation of possible species have been suggested and even improved, mainly within the context of the coalescence theory36,37 that takes into account standards and processes related to recent speciation events. 38 General Mixed Yule Coalescence (GMYC), one of the most common approaches to define species on the basis of data within a single locus, is widely used to evaluate biodiversity and phylogenetic ecological communities.39,40
The headwaters of the Ivaí River and the tributaries of the upper reaches of the Piquiri, Tibagi, Iguaçu, and Ribeira de Iguape River basins are located on lands between the Serra da Esperança and the Ponta Grossa Arch. Therefore, the phylogeographical study of fishes related to biogeographic barriers, such as the geological formations mentioned in this study, should contribute to the understanding of the evolution and spatial distribution of these organisms and may be important areas for testing related biogeographic hypotheses or focusing on the assumptions of phylogeography. To test whether the uplift of the Serra da Esperança and the Ponta Grossa Arch in the Serra Geral resulted in ichthyofaunistic changes in adjacent basins, we describe the phylogeographic structure among populations of Trichomycterus.
Materials and Methods
Sampling sites and DNA extraction
Specimens of the Trichomycterus were collected from five hydrographical basins in southern Brazil: Iguaçu River basin (28 collection sites); Ivaí River basin (10 collection sites); Piquiri River basin (13 collection sites); Tibagi River basin (16 collection sites); and Ribeira de Iguape River basin (11 collection sites) (Fig. 1). Samplings were approved (002/2012) by the Committee of Ethics for the Use of Animals in Experimentation, State University of Maringá (UEM), with permission for the collection and transport of zoological material SISBIO (ICMBio), process 14028-1 of 10/12/2008 to W.J.G.
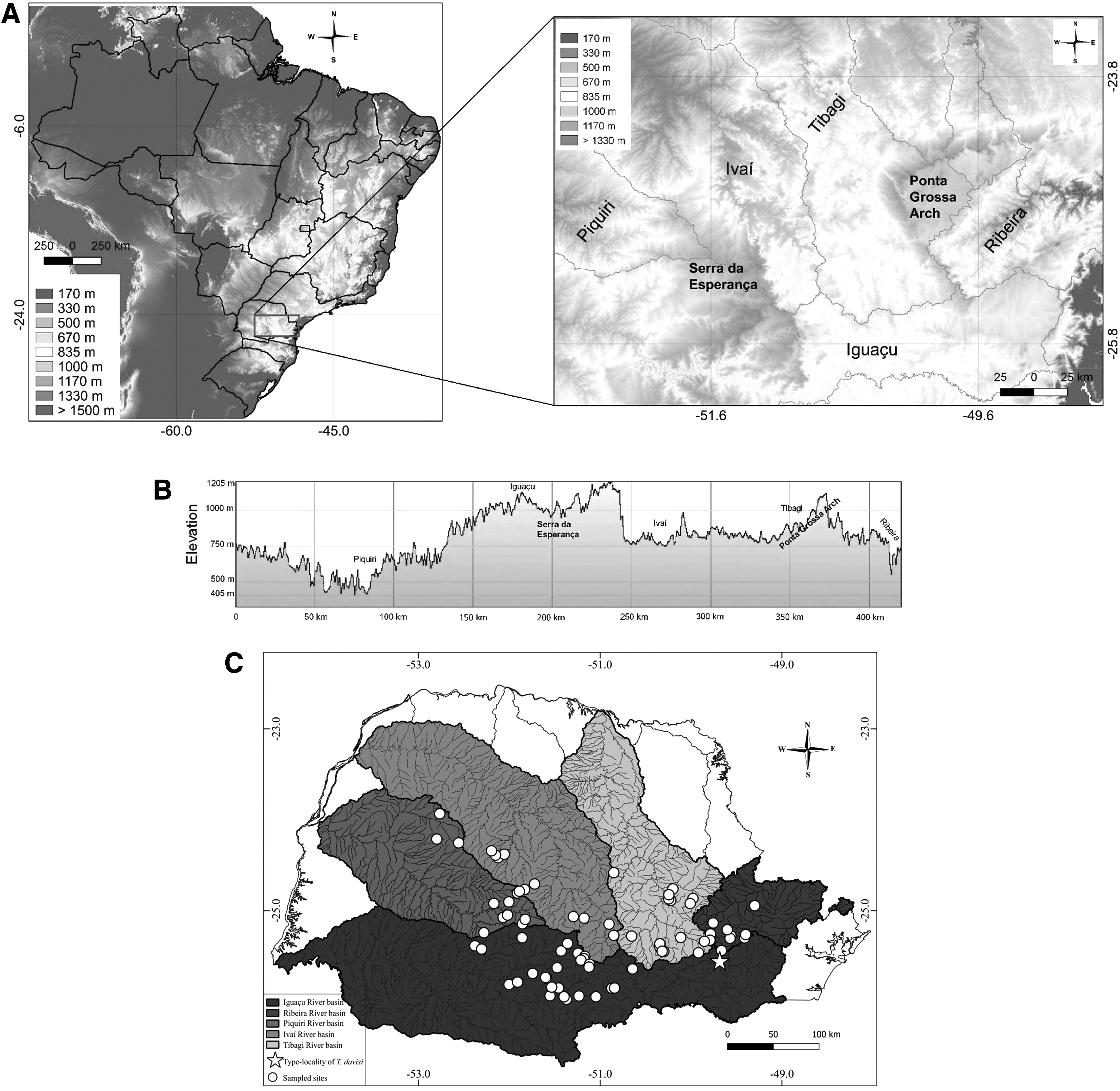
Highlights of the study area.
After collection, the specimens were anesthetized with benzocaine, as per the Resolution 1000/2012 of the Federal Council of Veterinary Medicine, Brazil. Fish muscle samples were removed, fixed in commercial ethanol, and stored at −20°C. Specimens were fixed in 70% alcohol and deposited in the ichthyologic collection of the Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (Nupélia) of the Universidade Estadual de Maringá, Maringá, Paraná State, Brazil.
DNA was extracted using commercial Invitek and Invitrogen kits, as per the manufacturers' instructions, and quantified with 1% agarose gels and Low DNA Mass Ladder (Invitrogen) using known concentration fragments for comparison.
PCR and sequencing
COI gene was partially amplified from 76 samples of Trichomycterus using primer pairs prescribed by Ivanova et al. 41 for fish: H7152 (5′ CAC CTCA GGG TGT CCG AAR AAY CAR AA 3′) and L6448-F1 (5′ TCA ACC AAC CAC AAA GAC ATT CGG CAC 3′). Amplification reactions were in a volume of 25 μL, with 10 ng template DNA, Tris-KCl (20 mM Tris-HCl pH 8.4 and 50 mM KCl), MgCl2 (1.5 mM), primers (2.5 mM each), dNTPs (0.1 mM each), and Taq DNA-polymerase (1 U). Reaction conditions comprised initial denaturation at 94°C for 2 min; 94°C for 30 s; hybridation at 52°C for 40 s; and elongation at 72°C for 1 min, followed by 35 cycles at 94°C for 30 s, 52°C for 40 s, and 72°C for 1 min. 41 Amplicons were quantified in 1.0% agarose gels, purified in polyethyleneglycol–NaCl following Rosenthal et al., 42 and then sequenced in both directions with a BigDye Terminator Kit. Nucleotide sequencing was determined using automatic sequencer ABI 3500.
Data analysis
Nucleotide sequences were visualized and manually edited by Chromas Lite 2.01, and their identity was verified by GenBank (www.ncbi.nlm.nih.gov) using BLASTN. Sequences were then aligned by algorithm Clustal W 1.6 43 implemented in MEGA 6. 44 The exclusion of identical sequences and identification of haplotypes were performed by DnaSP v5. 45
An ultrametric tree was built with BEAST 2. 46 A strict molecular clock was used and calibrated according to Bermingham et al., 47 with 0.006 per million years and a Coalescent (Constant Population Size) tree prior specified. Chain convergence was analyzed by Tracer 1.6 with an effective sample size >200, and 10% of the chain was discarded as burn-in. Later, the resulting tree was edited using the program FigTree. Subpartitions and models were estimated by PartitionFinder, 48 which indicated three subpartitions (first, second, and third codon positions) and the models TrNef+I, F81, and TrN+Γ for each codon position, respectively. A phylogenetic tree was built using maximum likelihood method with raxmlGUI, 49 considering the same subpartitions estimated by PartitionFinder and 1000 bootstrap resampling with rapid bootstrap algorithm. Parastegophilus paulensis was used as outgroup for both trees (GenBank access: GU701960).
The program SplitsTree 50 was used to construct a neighbor-net, or rather, the distance scale, in which the length of each line represents the divergence between each sample. 51 GMYC, 52 Poisson Tree Processes (PTP), 53 and Automatic Barcode Gap Discovery (ABGD) 54 were used for Operational Taxonomic Units delimitation test. GMYC was performed on platform R with the splits package, using the single threshold method. PTP and ABGD were performed on http://species.h-its.org/ptp and wwwabi.snv.jussieu.fr/public/abgd/abgdweb.html, respectively.
Results
Seventy-six DNA sequences were obtained from the partial fragment of the COI gene (603 bp) amplified from Trichomycterus samples, and 51 single haplotypes were used for analysis. Nucleotide composition of sequences was 29.2% (T), 27.3% (C), 24.9% (A), and 18.7% (G). All sequences were deposited in GenBank (KY231966–KY232041). The greatest number of sequences with the greatest number of haplotypes was obtained from the Iguaçu River basin, followed by the Piquiri, Tibagi, Ivaí, and Ribeira de Iguape River basins (Table 1).
Phylogenetic analyses
An ultrametric tree reconstructed in BEAST 2 software is shown in Figure 2. Most samples from the Iguaçu and Piquiri River basins were mixed. However, a group with three haplotypes of the Iguaçu River proved to be different from the other samples, with good support (Iguaçu II), similar to samples from the Tibagi River basin. All samples from the Ribeira site were grouped with high support precisely where the haplotype of the region of the type-site of T. davisi (Amola Faca Stream of the Iguaçu River basin, next to Serrinha) was included. Although samples from the Ivaí River also formed a haplogroup with good support, two haplotypes of this basin (Hap_38 and Hap_41) were grouped with samples from the Iguaçu and Piquiri River basins.

Ultrametric cladogram by Bayesian inference constructed from COI sequences of Trichomycterus populations in the Iguaçu, Ivaí, Piquiri, Tibagi, and Ribeira de Iguape River basins. The dashed line shows the threshold time according to the GMYC delimitation. COI, Cytochrome C Oxidase subunit I.
Samples were grouped to determine genetic distances between groups (Fig. 2), and correction was performed using Kimura-2-parameters (Table 2). Iguaçu, Piquiri, and the two haplotypes (Hap_38 and Hap_41) of the Ivaí River basin were considered a single group (Iguaçu + Piquiri). Intraspecies variation reached 1.34% for Iguaçu + Piquiri; 1.35% for Iguaçu II; 0.91% for Tibagi; 1.1% for Ivaí; and 0.36% for Ribeira + Amola Faca.
Neighbor-net also revealed the distinction between the sampled river basins (Fig. 3) and evidenced the high variability of sequences sampled in the Iguaçu and Piquiri River basins, which may have contributed toward the low support rates for most of the branches at the sites in the two analyses.
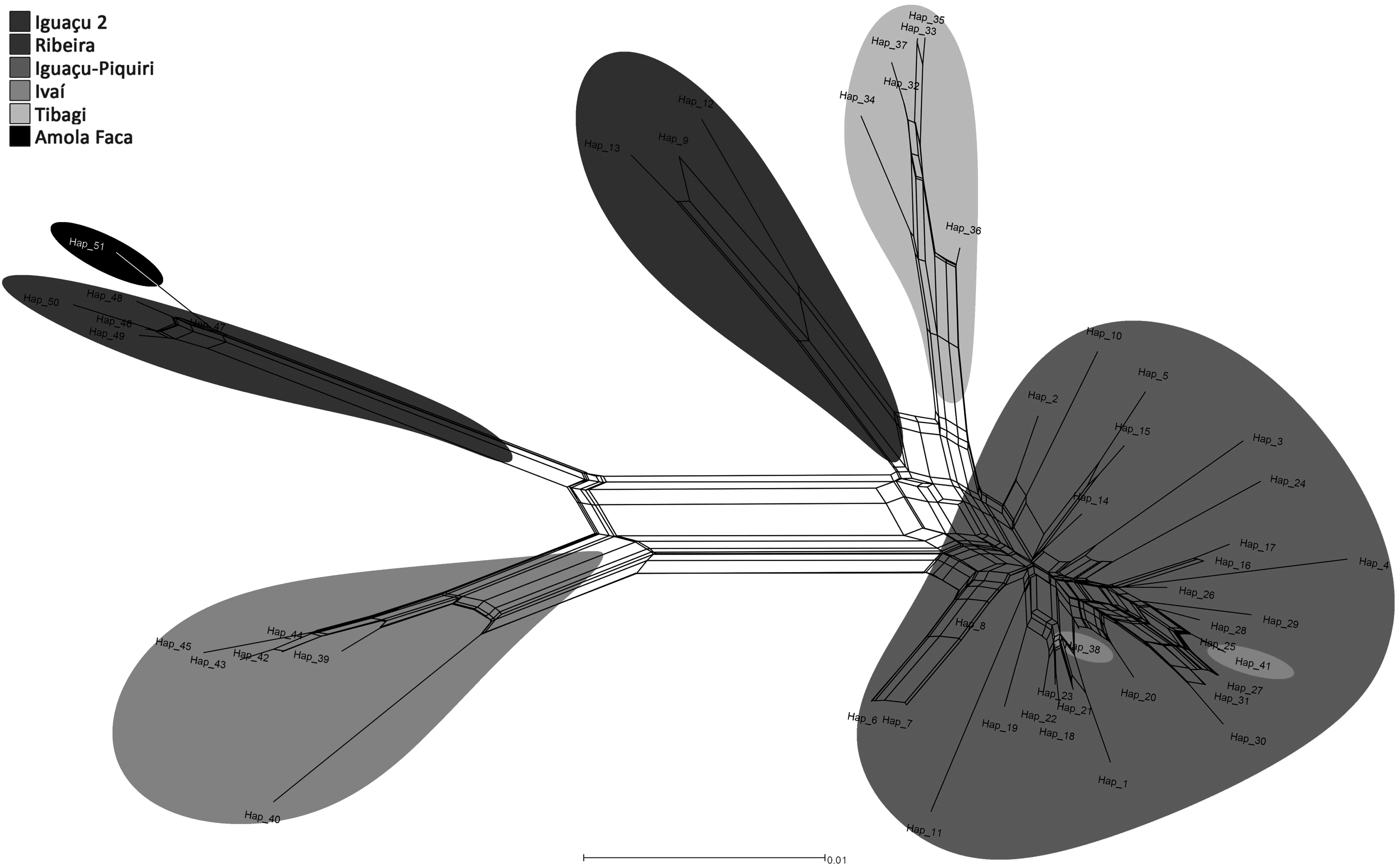
Network of Trichomycterus haplotypes sampled from the Iguaçu, Ivaí, Piquiri, Tibagi, and Ribeira de Iguape River basins on the basis of sequences of mitochondrial COI.
Characteristics diagnosed in the species Trichomycterus stawiarski could be identified in some specimens from the Iguaçu + Piquiri haplogroup. The other haplogroups featured morphological characteristics of T. davisi and thus confirmed previous analyses.
Species boundaries
The GMYC, PTP, and ABGD delimitation tests were performed to delimit possible cryptic species from the sampled specimens. The ultrametric calibrated tree was used in GMYC (null model = 100.9015, GMYC model = 102.6728; p = 0.17; threshold time = −1.7429, dashed red line in Fig. 2), and six possible Operational Taxonomic Units (OTUs) were delimited. Although the Iguaçu + Piquiri group was divided into two possible OTUs, samples from the two basins remained mixed. Samples from groups Tibagi, Iguaçu 2, Ivaí, and Ribeira + Amola Faca were delimited as separate OTUs. Estimated time of diversification by the delimitation test was 1.74 My. The time estimated for the cladogenesis between specimens of Ribeira de Iguape River, type-locality, and Ivaí basins and Tibagi, Iguaçu 2, and Iguaçu + Piquiri was 4.23 My (3.81–6.65 My, 95% high probability density interval).
The phylogenetic tree was used in PTP, and seven possible OTUs were delimited (Fig. 2 and Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/zeb). Instead, the division of Iguaçu + Piquiri, as observed in GMYC results, in PTP Iguaçu + Piquiri was grouped in a single OTU, and Iguaçu 2 was divided in two OTUs, while GMYC delimited only a single OTU. Also, Ivaí was divided in two OTUs, which was delimited as a single OTU in GMYC test. The phylogenetic tree and the results of the delimitation carried in PTP web server are presented in Supplementary Figure S1.
A distance matrix was used in ABGD, and five possible OTUs were delimited (Fig. 2). In this test, Iguaçu + Piquiri was grouped with Tibagi in a single OTU, and Ivaí was divided in two OTUs as in PTP delimitation. Nevertheless, all the delimitation testes used in this study delimited T. davisi as a single OTU, separated from all the other sampled specimens, and GMYC test was most congruent with previous morphological identification (Fig. 2).
Discussion
Molecular analyses show that, beside T. davisi sensu stricto, which only occurs next to the type-locality (in Amola Faca Stream of Iguaçu River basin, next to Serrinha region, Balsa Nova, PR, Brazil) and in the Ribeira do Iguape basin, at least five to six new species occur in the hydrographic basins analyzed, considering species concept as evolution of metapopulation lineages, 55 and depending on the delimitation test considered. The results actually increase the number of Trichomycterus species already described for the regions under analysis.56,57 Furthermore, they also show the occurrence of T. stawiarski in the Piquiri and Ivaí River basins and thus widen the distribution of a species that was considered endemic to the Iguaçu River basin. 57
All analyses have demonstrated that the fish population from the Ribeira de Iguape basin forms one monophyletic group, supported by a high posterior probability and bootstrap values in the tree reconstructions. Neighbor-net depicts the isolated condition of the Ribeira do Iguape haplogroup when compared to other populations. Among the contrasts investigated with the phylogroups revealed by the delimitation tests, the greatest distance belongs to the Ribeira de Iguape population, whose specimens remained grouped with those of the Amola Faca Stream of the Iguaçu River basin (next to Serrinha and the type-locality). As reported above, specimens of the Amola Faca Stream of Iguaçu River and Ribeira de Iguape basins may be attributed to T. davisi sensu stricto.
The threshold time estimated according to GMYC results was 1.7 My, which agrees with tectonic active areas in a period more recent than 1.6 My. 58 These events may have caused a sharing of ichthyologic fauna of the central and southern basins with coastal rivers, with special focus on areas that include the sources of the Ribeira de Iguape and Iguaçu Rivers. The aforementioned event may have caused the presence of the species T. davisi, whose type-locality lies in the Iguaçu River basin (in the Serrinha region, Balsa Nova, PR, Brazil), within the Ribeira do Iguape basin. A relevant aspect that should be emphasized is the Ponta Grossa Arch, which would have triggered the acceleration of river dynamics and enhanced ichthyofauna interchange between the adjacent drainages. This would explain ichthyologic exchanges that occurred between the coastal Ribeira de Iguape River and the Iguaçu River. 7
Two scenarios may explain the origin of T. davisi. First, the species originated in the Iguaçu River basin. The drainage of the Ribeira de Iguape River is at a lower topographic level than the headwaters of the Iguaçu River. Erosive processes have caused the capture of stretches of these headwaters, expanding the drainage area of the Ribeira de Iguape River along the first Plateau of Paraná. 59 Second, its origin occurred in the Ribeira de Iguape River basin, the more basal clade between all the other basins, according to our molecular data (Fig. 2). Significant tectonic reactivations, such as the Ponta Grossa Arch, may have been able to raise the terrain and interconnect coastal drainages (Ribeira de Iguape River basin) with drainages that flow into the interior of the continental plate (Iguaçu River basin). 7 This scenario is plausible by recent studies demonstrating recent movement along northwest to southeast fault zones, from the Miocene and Neogene to the present day. 60
Being a distinct OTU, the fish population of the Ivaí basin is closer to that of the Ribeira de Iguape than to other populations. It is important to underscore that haplotypes Hap_38 and Hap_41, which do not belong to the OTU identified in the current assay but to T. stawiarski, occur in the Ivaí basin. Data foreground the first inference on the populations analyzed, where in Ribeira de Iguape and Ivaí form a distinct clade from that of the other populations but within the T. davisi complex, where the emergence of the two clades anteceded all other cladogeneses.
T. davisi is widely distributed throughout the drainage of the Iguaçu River,12,16,57,61 which brings us to category V of the phylogeographic patterns 62 to explain the greater relationship between Ivaí and Ribeira de Iguape. In this case, ancestral genotypic distribution over a wide area is presumed so that newly emerged mutations are not yet spread throughout the range of a species. 62 The headwaters are located at the highest topographic levels of the Serra da Esperança between the divisions of the river basins of the Ivaí and Iguaçu rivers. This may explain the greater isolation of this area, even though it is more distant from the drainage basin of the Ribeira de Iguape River than the Tibagi River basin.
The population of the Tibagi basin also demonstrated monophyly in PTP and GMYC tests, and its grouping is sustained by a high posterior probability and bootstrap values. It substantially diverged from all the others. However, the Tibagi haplogroup lies nearer the haplogroup of the Iguaçu and Piquiri basins than those of Ribeira de Iguape and Ivaí. Delimitation tests of GMYC and PTP identified the population of the Tibagi River as a distinct OTU, as expected from pronounced nucleotide divergence. By the criterion of a 2% cut, usually used by researchers, 63 the distances (coupled to GMYC and PTP results) suggest that the Tibagi population should not be identified as T. davisi, and it may be considered a new Trichomycterus species, distinct from T. davisi sensu stricto.
Further analyses with other molecular markers are needed to test the status of a single species (T. stawiarski) or the possibility of a species complex within the phylogroup. Although most specimens from the Iguaçu River basin are grouped in the Iguaçu-Piquiri phylogroup, the GMYC, PTP, and ABGD tests revealed at least two other OTUs in the basin. The Iguaçu + Piquiri group and a second OTU would comprise the haplotypes Hap_13, Hap_9, and Hap_12. In the analysis, these haplotypes form a concise grouping with high posterior probability and bootstrap support values. Consequently, results show that two species should be in the Iguaçu River basin, or rather, T. stawiarski and a second OTU belonging to the T. davisi complex. The latter is more closely related to the T. davisi population of the Tibagi River basin, a fact that corroborates the share of some species, such as T. castroi, Hypostomus ancistroides, and Rhamdiopsis moreirai, 19 previously thought to be restricted to the upper Ribeira de Iguape and Iguaçu basins.
Most specimens from the Iguaçu + Piquiri phylogroup belong to the ichthyofauna of the Iguaçu River basin, with their special evolution history featuring high rates of endemism.57,64 The lack of morphological differences among the other clades clearly reveals a cryptic species case. The geomorphologic formation process of the basin hails from the Tertiary period and is associated with vertical movements causing the rise of the Serra do Mar and the origin of the three plateaus of the state of Paraná, Brazil. 65 The southwestern edges of the Brazilian Shield represent a major source of biogeographic differentiation between the costal Atlantic lowlands (such as the Ribeira de Iguape River basin) and the elevated inland river basins (such as the Iguaçu River basin).7,66 The Quaternary tectonic activity causes recent drainage rearrangements, and it is involved in the sharing of several other species among the basins sampled in this study, such as Hyphessobrycon griemi, Hyphessobrycon bifasciatus and Hyphessobrycon reticulatus, 16 Characidium travassosi, 67 and Mimagoniates microlepis. 7 The identification of an area of endemism comprising the limits between the sampled basins would allow a better understanding of the evolution of this regional biota 68 and should be prioritized for local biodiversity conservation. 69
Although the Iguaçu River is a geologically ancient river, it has been under renewal by several features, such as rapids and waterfalls, alternated by meanders with wide curves and extensive meadows. 70 These habitat characteristics may enhance the isolation of fish populations and permit significant evolution processes in the communities, eventually leading toward speciation. Isolated populations are more liable to evolutionary changes and acceleration depends on the different environments into which the populations are inserted. In fact, small populations undergo faster evolutionary process due to selective pressures in divergent environments, 71 as well as stochastic effects. 72 A very close example was found for Hypoptopomatinae, which Neoplecostomini lineages exhibited increased phenotype diversification rates characterizing river rapids as incubators of biological diversification and specialization. 71
In the current analysis, we found six to seven different species, depending on the delimitation test considered. Most of specimens presented morphological characters of the T. davisi. However, some specimens from the Iguaçu, Piquiri, and Ivaí basins presented morphological characters of T. stawiarski. Analysis with a species delimitation tests revealed that T. davisi does not contain the genetic diversity scale of the specimens collected in the southern Brazilian hydrographic basins and demonstrates at least six genetic lineages in Trichomycterus. Despite the low posterior probability for OTUs in the species delimitation tests, interspecies distance rates and high support values for groups formed in the ultrametric tree support such results.
The data suggest the importance of molecular analyses in assessing diversity and in establishing distinct genetic groups in Trichomycterus populations in adjoining basins. Several research projects have successfully used barcode DNA to detect new species of Neotropical fish and identify cryptic species.35,73 The current investigation demonstrated that rates of interpopulation genetic distances indicated by COI comply with the 2% threshold proposed for the delimitation of species, with 95% of the 1088 barcode sequences of fish revealing intraspecies rates with less than 2% distance. 63 Hebert and Gregory 29 suggested that new species can be proposed when a threshold 10 times higher than the mean intraspecies divergence is reached. In the current analysis, the threshold was reached when the group formed by Ribeira do Iguape and Amola Faca was compared with the other clades. In the other groups, intergroup distance lies at a threshold approximately three to five times higher than intragroup rates. However, the comprehensive concept by Hebert and Gregory 29 was criticized by other authors74,75 and may not be valid for all fish groups, including the current case.
It should be understood that high mean rates for intragroup distance in certain clades are because of the great variety of haplotypes. The phylogroup Iguaçu + Piquiri should be given special attention because its haplotypes may significantly differ and reach a 2.2% rate (Hap_30 para Hap_6). In this specific case, large diversity in the phylogroup may be related to the sampling range. Specimens were collected at different sites of the basin, sometimes 290 km apart, frequently separated by geographic barriers that favor isolation and establishment of genetic differences among the specimens.
In conclusion, our data support cases of cryptic diversity and undescribed species in the T. davisi species complex. A taxonomic study of the distinct units herein founded is recommended. The interpretation that genetic structure in T. davisi is explained by tectonic events intrinsic to the areas of influence of Serra da Esperança and the Ponta Grossa Arch that occurred around 1.7 My, which resulted in a river dynamism between the various drainages studied here; or by isolating populations and promoting the formation of new species; accelerating the exchange of the ichthyofauna by capture of headwaters. In addition, other species occurring in the studied basins must be analyzed to understand what events have contributed to the present diversity of the ichthyofauna of the rivers that occur in the region of the Ponta Grossa Arch and the Serra da Esperança.
Footnotes
Acknowledgments
The authors are grateful to Fundação Araucária (Seti-PR) for the financial support, to Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the project funding and research scholarship, and to the Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (Nupélia) for the infrastructure made available for this research.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.