
Abstract
Abstract
Astyanax is a genus with a wide distribution ranging from the south United States to north of Patagonia (Argentina). The available cytogenetic data on Astyanax indicate a high karyotypic diversity, with diploid number of 36–52 chromosomes, presence of B chromosomes, heterochromatin polymorphism, and variations with respect to the number and localization of nucleolar organizer regions (NORs) and 18S and 5S ribosomal DNA sites. In the present study, we estimated the evolutionary history and times of divergence for 10 nominal Astyanax species from the South and Central/North American (Cna) continents, which present distinct chromosomal characteristics, based on molecular clocks inferred from mitochondrial DNA sequence. The molecular clock results indicate the origin of three distinct clades (Humeral dark spot [Hds]; Diffuse humeral spot [Dhs]; Cna group) during the late Miocene about 11.2 million years ago (Mya). Thus, Astyanax mexicanus (Cna) represent a species that diverged a long time ago (∼8.6 Mya) from the Hds group, and Astyanax schubarti is the oldest species (∼6.5 Mya) among the Dhs species.
Introduction
Astyanax is the most species-rich genus of the family Characidae with 146 validated species, 1 distributed from Texas and New Mexico in the United States 2 to the north of Patagonia in Argentina. 3 These species may occur broadly in some hydrographic basins (e.g., Astyanax altiparanae, Astyanax fasciatus) or have restricted distribution to a few tributaries or brooks (e.g., Astyanax paranae).4,5
Recently, using Cytochrome C Oxidase subunit I (COI) sequences, Rossini et al. 6 found 5 clades out of 64 nominal species plus 12 provisionally identified taxa. The authors make the separation in Clade 1, which mainly includes species belonging to the A. fasciatus and Astyanax scabripinnis species complexes, the Clade 2 encompasses the Central American species, the Clade 3 is primarily formed by species of the Astyanax bimaculatus complex, and the Clades 4 and 5 correspond to the remaining species.
The available cytogenetic data on Astyanax indicate wide karyotypic diversity with 2n = 36 chromosomes in Astyanax schubarti7–11 and Astyanax correntinus 12 ; 2n = 46 and 2n = 48 in some populations of A. fasciatus8,9,13–15; 2n = 50 for the majority of Astyanax species, as A. altiparanae, 16 A. paranae, 17 Astyanax abramis18,19; and 2n = 52 chromosomes in Astyanax sp. 20 Additionally, variations with respect to the number and localization of nucleolar organizer regions (NORs) and 18S and 5S ribosomal DNA (rDNA) sites,21–25 presence of B chromosomes, 22 and heterochromatin polymorphism, 26 which are suitable markers for the study of karyotypic evolution of the genus, occur in this group.
Cytogenetics has played an important role in recent decades regarding the study of the evolution of chromosomes in different plant and animal groups.27–29 From this, the use of chromosomal data combined with molecular analysis can contribute to the understanding of the evolutionary dynamics of chromosomes in the Astyanax group.
The molecular clock hypothesis states that DNA sequences evolve at a rate that is relatively constant over time and among different organisms. Thus, the genetic difference between any two species is proportional to the time, since these species last shared a common ancestor. Therefore, if the molecular clock hypothesis holds true, this hypothesis serves as an extremely useful method for estimating evolutionary timescales. 30 In this context, the aim of the present article was to estimate the evolutionary history and times of divergence of different Astyanax species with different diploid numbers based on molecular clocks inferred from nucleotide sequences of the COI.
Materials and Methods
Statement of ethics
All the institutional guidelines for the care and use of laboratory animals were followed. Animals were captured with the permission of the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio; number 43497-1) and used for laboratory experiments with approval from the Animal Experimentation Ethics Committee (CEUA; protocol 2335) of Universidade Estadual Paulista “Júlio de Mesquita Filho”.
Sampling and classical cytogenetics
We analyzed five individuals of A. altiparanae from the Piracicaba River in São Paulo State (SP), Brazil; six individuals of Astyanax marionae from the Rio Claro Stream in Mato Grosso State (MT), Brazil; three individuals of A. fasciatus from the Ribeirão Claro River (SP), Brazil; one individual of A. schubarti from the Piracicaba River (SP), Brazil. Voucher specimens were deposited in the fish collection of the Laboratório de Citogenética (LC), Universidade Estadual Paulista “Júlio de Mesquita Filho,” Rio Claro, SP, Brazil, as A. altiparanae (LC 1822), A. abramis (LC 100), Astyanax asuncionensis (LC 5851), Astyanax bockmanni (LC 1528), Astyanax eigenmanniorum (LC 1107), A. marionae (LC 7539), A. fasciatus (LC 1784), and A. schubarti (LC 1826). Mitotic metaphase chromosomes were obtained according to the technique proposed by Foresti et al. 31 The NORs were identified after impregnation by silver ion using the technique of Howell and Black. 32 To identify guanine and cytosine (GC)/adenine and thymine (AT)-rich regions, double staining with chromomycin A3/4′,6-diamidino-2-phenylindole (CMA3/DAPI) was performed as described by Piscor et al. 18 Chromosomes were classified as metacentric (m), submetacentric (sm), subtelocentric (st), and acrocentric (a) according to the ratio between long arm (q) and short arm (p). 33 For the determination of the fundamental number (FN), that is, number of chromosome arms, the m, sm, and st chromosomes were calculated as bearing two arms; and the a chromosomes, only one arm.
DNA extraction, probe synthesis, and fragment amplification
The genomic DNA was extracted from fins of the samples of Astyanax species as described by Sambrook and Russell. 34 The 18S rDNA probe was amplified by polymerase chain reaction (PCR) using specific primers as described by White et al. 35
The PCR analyses were performed using the primers described by Ivanova et al. 36 for amplification of the mitochondrial gene COI. The PCR product was visualized on 1% agarose gel; purification and sequencing was performed by a third-party company (Macrogen, Korea).
Fluorescence in situ hybridization
The fluorescence in situ hybridization (FISH) experiments were performed according to Pinkel et al., 37 with modifications described by Cabral-de-Mello et al. 38 The 18S rDNA probe was labeled through nick translation, with digoxigenin-11-dUTP (Roche, Mannheim, Germany), and signals were detected using anti-digoxigenin-rhodamine (Roche, Mannheim, Germany). Slides were mounted with Vectashield Mounting Medium (Vector, Burlingame, CA) containing DAPI (4′,6-diamidino-2-phenylindole). Chromosomes and fluorescent signals were visualized with an Olympus BX51 microscope coupled to a digital camera (Olympus model D71). Images were captured using a DP controller software.
Phylogenetic analysis and cladogenesis dating
The COI gene was amplified, sequenced, and deposited in GenBank (www.ncbi.nlm.nih.gov/genbank/) for A. schubarti, A. abramis, A. asuncionensis, A. bockmanni, A. eigenmanniorum, A. marionae, A. altiparanae, and A. fasciatus (accession numbers MH718733, MH718734, MH718735, MH718736, MH718737, MH718738, MH718739, MH718740, respectively). One COI sequence of Astyanax nasutus and two of Astyanax mexicanus were obtained from the GenBank database (accession numbers FJ439390.1, GU702225.1, and KM043816.1, respectively). Other two non-Astyanax sequences were obtained from the GenBank as well—Serrapinnus notomelas (Characidae, Cheirodontinae) from a related genus and the outgroup Rhamdia quelen belonging to the Siluriformes (accession numbers HM126642.1 and MG449795.1, respectively); those sequences were used to root the phylogenetic trees and provide good calibration points for the molecular clock.
Sequences were aligned on Dambe 5.0.8 software 39 using the ClustalW algorithm 40 under default parameters and eye-checked later for any obvious misalignments.
To estimate phylogenetic trees and date cladogenetic events, BEAUti v1.7.2 and BEAST v1.7.2 softwares 41 were selected. The “Lognormal relaxed clock (uncorrelated)” model was chosen for the molecular clock and the “Yule” process of speciation for the “Tree Prior” parameter. Three calibration points were included in the analyses; the colonization of Central America by the genus (which enabled the origin of Astyanax species in Central America) was dated by Ornelas-García et al. 42 (about 7.8–8.1 million years ago [Mya]); thus, the first calibration point was implemented using a log-normal prior offset to 6 Mya with a standard deviation of 0.7 for the origin of the clade that contains all Central American Astyanax species present in the analyses; the second calibration point regards the separation of A. mexicanus from the remaining Central American Astyanax species by the uplift and final closure of the Trans-Mexican Volcanic Belt about 3–6 Mya; thus, the second calibration point was implemented using a log-normal prior offset to 4.3 Mya with a standard deviation at 1.0 for the origin of the clade with all A. mexicanus specimens; the last calibration point was based on a fossil record of Colossoma macropomum, dated to at least 15 Mya 43 ; it was used to constrain the time of the most recent common ancestor of all Characiformes in the analyses; thus, the third calibration point was implemented using a log-normal prior offset to 15 Mya with a standard deviation at 0.5 for the origin of the clade with all Characiformes specimens analyzed here.
The analysis was performed in two independent runs with 10,000,000 generations each and parameters sampled every 1000 steps. The 10,000 initial samples were discarded (burn-in of 10%), the others were summarized, and the consensus tree with the divergence times was obtained using the TreeAnnotator v1.7.2 software 40 and visualized in the FigTree v1.4.2 software. 44 To check if the convergence parameters were satisfactory, the Tracer 1.6 software was used. 45
Results
Cytogenetic data in the Astyanax
The diploid number for A. altiparanae was 2n = 50 chromosomes—karyotype formula of 6 m + 20 sm +8st +16a, and FN of 84. A. marionae showed 2n = 48 chromosomes—karyotype formula of 8 m + 24 sm +10st +6a, and FN = 90. A. fasciatus presented 2n = 46 chromosomes—karyotype formula of 8 m + 18 sm +16st +4a, and FN = 88; and A. schubarti showed 2n = 36 chromosomes—karyotype formula of 12 m + 16 sm +4st +4a, and FN = 68 (Fig. 1a–d).
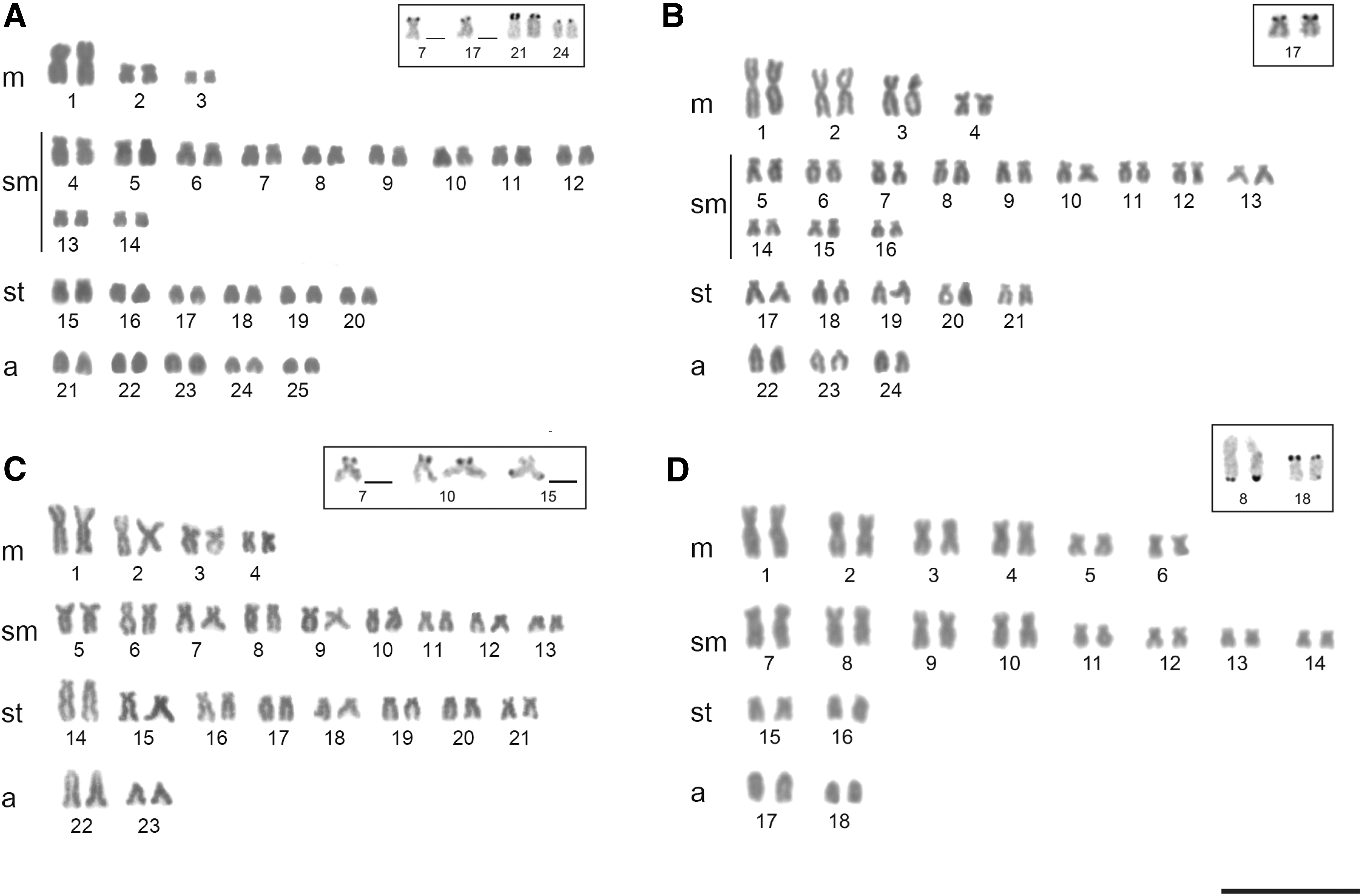
Karyotypes:
The Ag-NOR sites were identified on pairs 21 and 24, and in one of the homologs of pairs 7 and 17 in A. altiparanae, on pair 17 in A. marionae, on pair 10 and in one of the homologs of pairs 7 and 15 in A. fasciatus, and on pairs 8 and 18 in A. schubarti (Fig. 1, box). The majority CMA3 fluorochrome sites corresponded to the 18S rDNA sites, while DAPI staining was negative in these regions (Fig. 2).
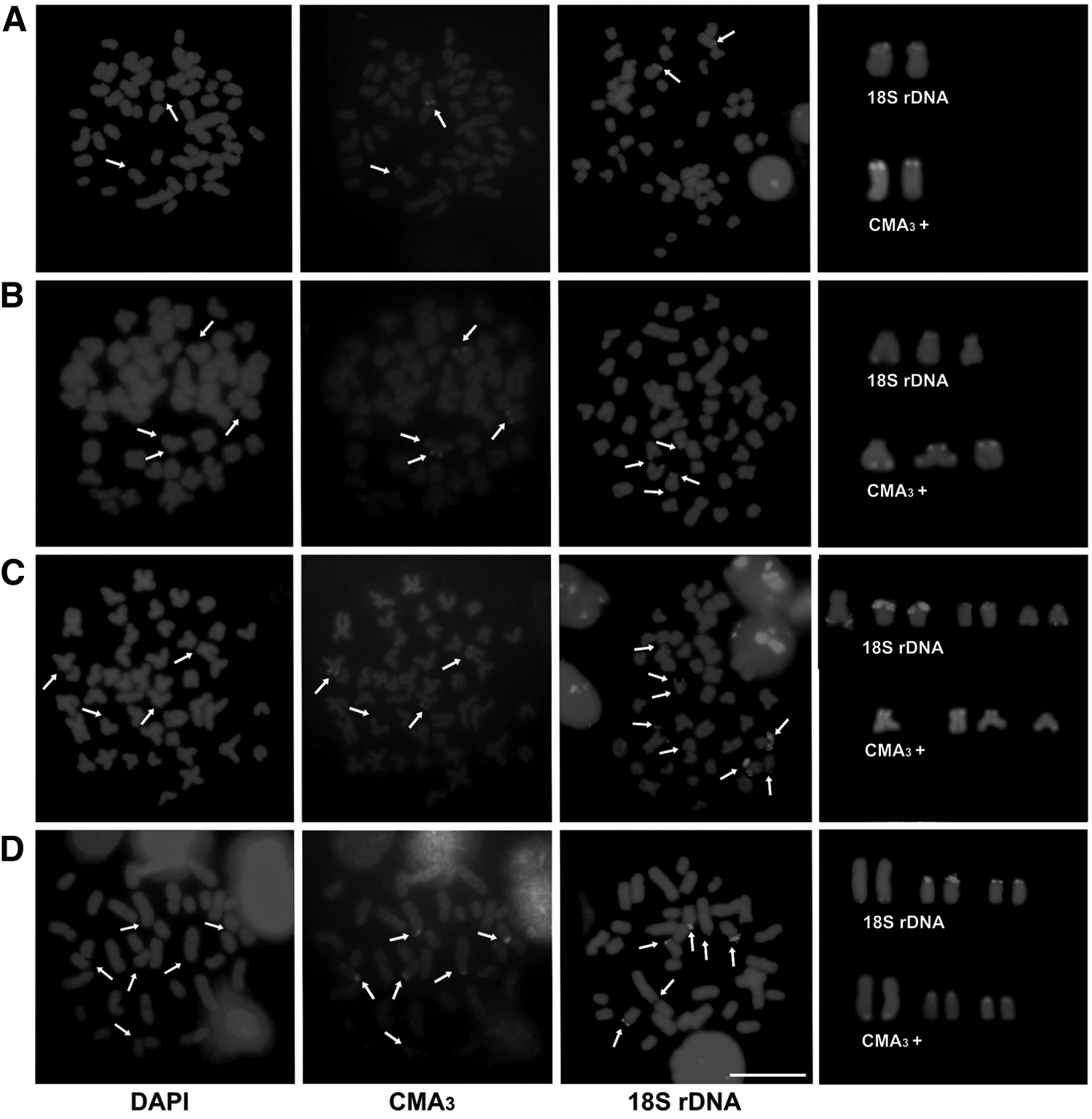
Chromosomes with guanine and cytosine-rich regions and fluorescent signals of repetitive sequences.
Fluorescent signals of 18S rDNA were observed on one pair (terminal region) in A. altiparanae, on three chromosomes all with terminal sites (one homologous with bi-arm marking) in A. marionae, on eight sites (terminal region) in A. fasciatus, and on six sites (terminal region) in A. schubarti (Fig. 2).
Literature analysis and molecular clock
A total of 10 partial mitochondrial DNA sequences were obtained from individuals with 4 different diploid chromosome number (2n = 36, 46, 48, and 50), corresponding to 10 nominal species of Astyanax from the South and Central/North American (Cna) continents. The COI sequence size was 678 bp for all species.
The molecular clock (present article) and cytogenetic data were plotted on the branch 4 in Figure 4 shown by Coutinho-Sanches and Dergam 46 (Fig. 3). The branch was separated into three clades according to the occurrence of these species: 1, Humeral dark spot (Hds) group; 2, Diffuse humeral spot (Dhs) group; and 3, Cna group (note that two South American groups were differentiates in humeral dark and diffuse humeral; Fig. 3).
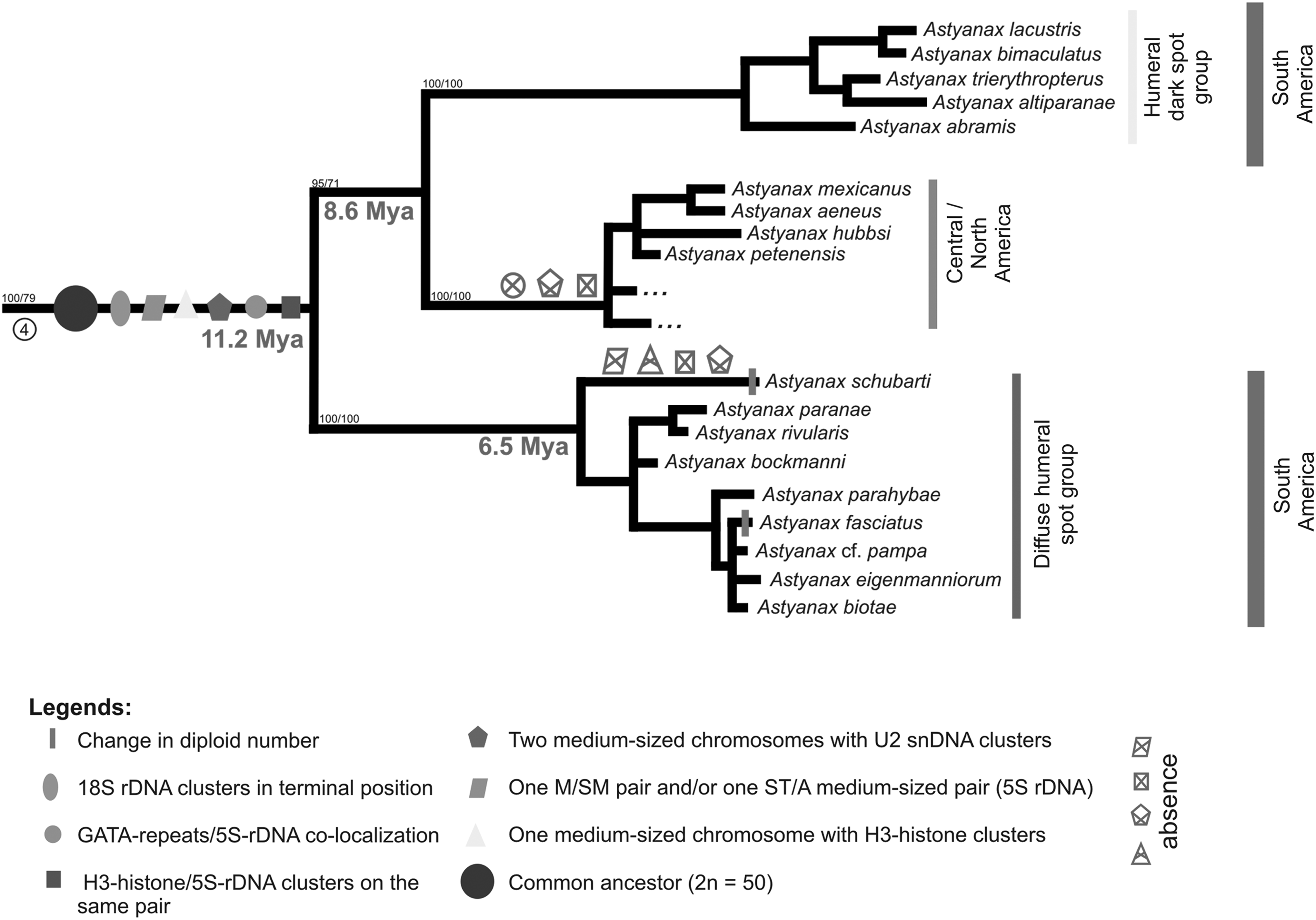
Cladogram adapted from Coutinho-Sanches and Dergam. 46 The gray numbers indicate the separation in million years ago. The X inside the geometric drawing indicates the absence of respectively cytogenetic characteristics in A. schubarti and Astyanax Central/North American branches.
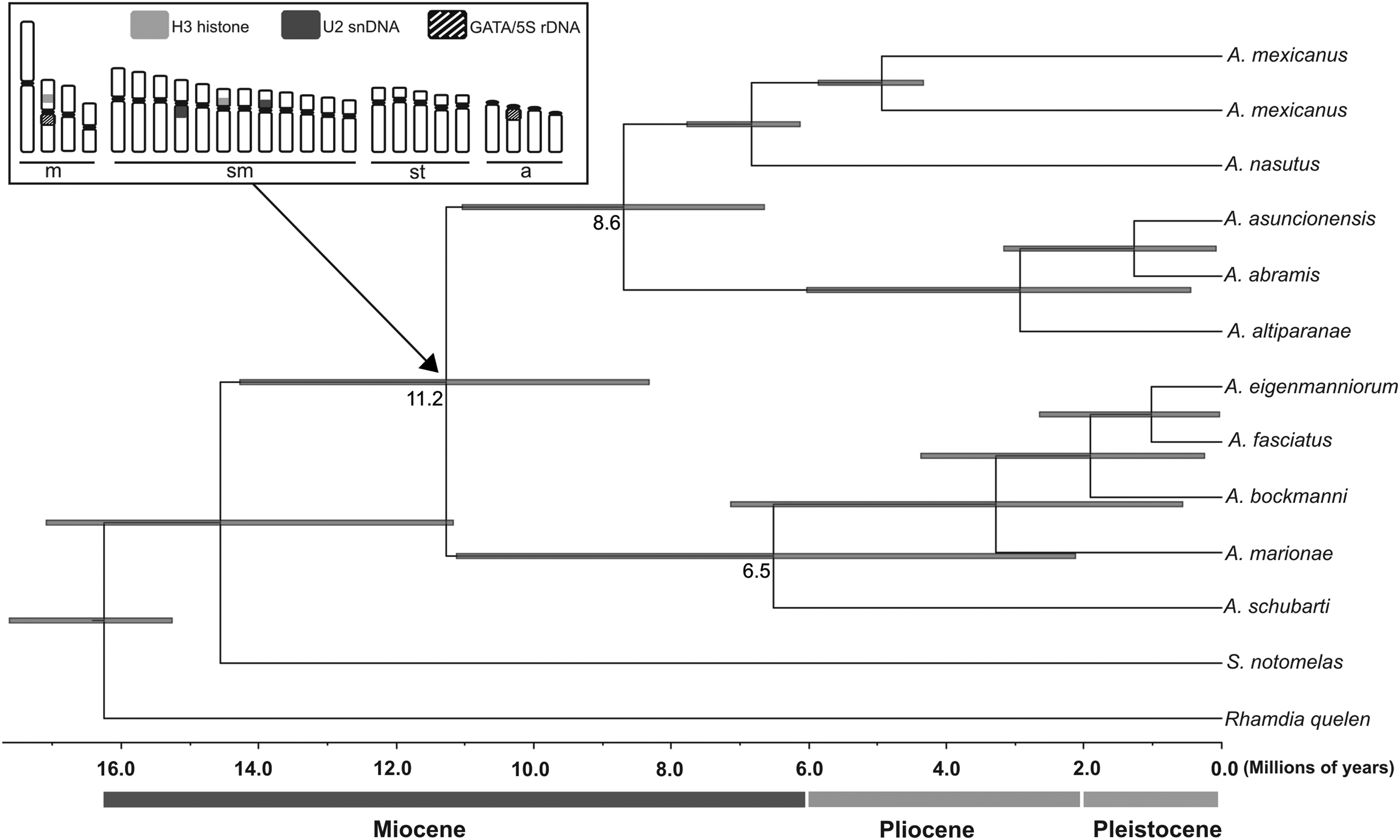
Calibrated Bayesian tree, based on the COI gene, showing the relationship within the Astyanax genus. Time, in millions of years, is shown on the x-axis. Numbers at branches are the dating of cladogenetic events, and gray bars (on the branches) represent 95% highest posterior density interval of divergence time estimates. In the top left square is the illustration of the ancestor karyotype with an arrow showing its probable origin. COI, Cytochrome C Oxidase subunit I.
The origin of cytogenetic characteristics was hypothesized from their presence and absence in the species analyzed in this article (Fig. 3). The branch 4 from Figure 4 shown by Coutinho-Sanches and Dergam 46 also was supported by phylogenetic analysis in the present article (Fig. 4). According to the molecular clock, we observed that the Hds and Cna groups diverged about 8.6 Mya, and the groups Hds/Cna and Dhs diverged about 11.2 Mya (note that A. schubarti was the species with ancient origin within the Dhs group—6.5 Mya; Fig. 4). The use of chromosomal data combined with molecular analysis allowed us to propose an ancestral karyotype for Astyanax, with 2n = 50 chromosomes, 18S rDNA in terminal position on the short arm, GATA-repeats/5S rDNA co-localization, H3-histone/5S rDNA on the medium-sized pair, two medium-sized chromosomes with U2 snDNA clusters, 5S rDNA clusters in one m/sm pair, and/or one st/a medium-sized pair and one medium-sized chromosome with H3 histone clusters (Figs. 3 and 4).
Discussion
The present study revealed four different diploid chromosome number, 2n = 36 for A. schubarti, 2n = 46 for A. fasciatus, 2n = 48 for A. marionae, and 2n = 50 for A. altiparanae, with karyotypes dominated by bi-armed chromosomes. Similar results were also found in other species of Astyanax: 2n = 36 chromosomes in A. correntinus, 12 2n = 46 and 2n = 48 in A. scabripinnis, 22 and 2n = 50 for almost all other species (e.g., A. abramis, A. bockmanni, A. asuncionensis, Astyanax jacuhiensis).12,18,19,47
The NOR phenotypes (Ag-NOR and FISH-18S), observed in karyotypes, differed among these four Astyanax species analyzed, with presence of two (A. altiparanae) to eight (A. fasciatus) sites located in terminal position (on both p- and q-arms). According to Nakajima et al., 48 the terminal regions of chromosomes can promote transposition events, leading to the dispersal of DNA segments, which could be responsible for variations in NOR in Astyanax species. Despite the numerical variation of the 18S rDNA sites, the terminal chromosomal position did not change among the different species, suggesting the ancestral state that diverged about 11.2 Mya. Other repetitive sequences are less dynamic than the 18S rDNA, as the 5S rDNA that is located in one medium-sized chromosome pair in Astyanax species.10,15,18,23,49–51
Our study also revealed that NORs were CMA3-positive and DAPI-negative, demonstrating that the 5, 8S, 18S, and 28S rDNA sequences are interspaced by GC-rich sequences. The presence of GC-rich blocks equivalent to NOR sites, as evidenced by CMA3 staining, is practically a constant characteristic of fish species, including Astyanax.20,23
In the previous articles, A. mexicanus8,10 and Astyanax jordani 50 presented similar chromosomal locations of U2 snRNA and H3 histone clusters (one pair with U2 snDNA clusters and two pairs with H3 histone clusters; both gene clusters were located in different chromosomes) in comparison with the number of clusters in other species of Astyanax (two pairs with U2 snDNA clusters and two pairs with H3 histone clusters; both gene clusters were located in different chromosomes).8,10,50 The variation in these clusters may be related to the restricted location of A. mexicanus and A. jordani (Central America), which would facilitate the fixation of these changes, which may have occurred after the separation of this Cna group about 8.6 Mya.
The molecular results showed that A. schubarti would be the species with an ancient origin with an approximate derivation of 6.5 Mya inside the Hds group. This species showed change in the diploid number (2n = 36), and cytogenetic characteristics differed from those found in the ancestral karyotype, as 5S rDNA clusters in one m/sm pair and/or one st/a medium-sized pair, H3-histone/5S-rDNA clusters in one medium-sized pair, two medium-sized chromosomes with U2 snDNA clusters, and one medium-sized chromosome with H3 histone clusters.8,10 Curiously, Astyanax currentinus 12 and A. schubarti 8 presents 2n = 36 chromosomes and a similar distribution pattern of 5S rDNA clusters. Unfortunately, there are no available phylogenetic data for A. currentinus to support the hypothesis that it is phylogenetically related with A. schubarti.
Based on morphological and molecular data, Astyanax has been considered a polyphyletic group by several authors.6,52–55 According to the molecular analyses presented by Coutinho-Sanches and Dergam 46 and the molecular clock (present study), A. schubarti, A. fasciatus, A. bockmanni, A. marionae, and A. eigenmanniorum form a clade, as well as A. mexicanus is closer to North/Central American species. These data also are supported by Rossini et al. 6
The Cna group has diverged around 8.6 Mya from the Hds group. Some hypotheses are proposed for colonization of Mesoamerica by freshwater fishes from South America.55,56 Bermingham and Martin 56 and Perdices et al. 57 showed that, according to molecular clock estimates, the invasion of freshwater fish started when the Panamanian Isthmus began its uplifting around 4–7 Mya. The study of Ornelas-García et al. 42 reinforces the hypothesis of a period of Mesoamerican colonization-expansion of Astyanax species from South America about 7.8–8.1 Mya, similar to the divergence between Hds and Cna groups shown in this article (∼8.6 Mya—late Miocene).
Thus, the relationship between cytogenetic and molecular data permitted two conclusions. First, A. mexicanus has restricted location in North/Central America and represents a species that diverged a long time ago (∼8.6 Mya) from the Hds group, accumulating distinct chromosomal characteristics. Second, contrary to what was proposed by Morelli et al., 7 A. schubarti is the oldest species (∼6.5 Mya) among the Dhs group and may have a particular chromosome system compared with other species of the genus.
Footnotes
Acknowledgments
We are grateful to Francisco Langeani Neto and Ricardo Britzke for species identification. This study was supported by Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Disclosure Statement
No competing financial interests exist.