
Abstract
Abstract
Selection criteria for sperm cryopreservation are highly relevant in zebrafish since sperm quality is particularly variable in this species. Successful cryopreservation depends on high-quality sperm, which can only be ensured by the selection of breeders. Consequently, male selection and management are a priority to improve cryopreservation, and therefore, this study aimed to characterize optimal age and sperm collection frequency in zebrafish. For this purpose, males from wild type (AB) and from a transgenic line [Tg(runx2:eGFP)] were sampled at 6, 8, 12, and 14 months. For each age, sperm were collected at time 0 followed by samplings at 2, 7, and 14 days of rest. Sperm quality was assessed according to motility and membrane viability parameters. Quality assessment showed that Tg(runx2:eGFP) displayed significantly higher motility than AB and younger males showed higher motility in both lines. Sperm collection frequency affected membrane viability. While AB fish recovered sperm viability after 14 days of rest, Tg(runx2:eGFP) could not recover. Consequently, it may be important to study the sperm quality of each zebrafish line before sperm cryopreservation. Taking into consideration the results achieved in both lines, sperm collection should be performed between 6 and 8 months of age with a minimum collection interval of 14 days.
Introduction
Successful cryopreservation depends on several factors, the selection of high-quality sperm being one of the most important factors, since the freezing process induces damage that decreases sperm quality significantly. 1 Given that high-quality sperm is related to high-quality breeders, 1 the selection of male donors is essential for broodstock management and cryopreservation programs. Zebrafish is an established model species maintained in laboratories worldwide and extensively used in numerous research fields, including biological and biomedical research.2,3 As a consequence, in the past years, abundant and valuable wild type, mutant, and transgenic zebrafish lines were established, posing problems in terms of space and management. To solve this issue, sperm cryopreservation can be used to support zebrafish facility management and to safeguard all those valuable genetic resources. 4 Zebrafish sperm cryopreservation was achieved for the first time more than 30 years ago by Harvey et al. 5 However, until today, the most relevant issue for successful and reproducible results using cryopreservation is the lack of methodological standardization among laboratories and rearing facilities, which translates into high variability in post-thaw sperm quality and in vitro fertilization success.
Sperm quality is defined by the ability of sperm to successfully fertilize an egg,6,7 which is dependent on factors such as heritage, 8 spermiation period, favorable environmental conditions for activation of sperm motility, 9 parental age,10,11 and sperm output frequency.
It has been reported that the age of males affects both sperm production and quality,10,12 resulting in lower reproductive success. This phenomenon is associated to the accumulation of de novo mutations in germ cells,8,13 thus decreasing the genetic quality of gametes9,13 and altering sperm functionality. It has been reported that in humans, age is associated with lower sperm volume, motility, and percentage of normal sperm cells.11,12 Furthermore, advanced parental age in several species is associated with a decline in sperm competition.9,14 However, the decrease of sperm competition with age was not observed in teleosts, such as reported for sockeye salmon (Oncorhynchus nerka) 15 in in vitro fertilization experiments and for guppy (Poecilia reticulata) natural spawns. 16 Consequently, the effect of age on sperm quality is not similar in all vertebrates and should be investigated thoroughly in zebrafish to ensure the highest sperm quality for cryopreservation purposes.
From an animal welfare and practical point of view, the most convenient technique for sperm collection in a zebrafish facility is through abdominal massage, since it is a nonlethal technique. 17 In this way, sperm collection can be performed repeatedly on the same male. 18
The influence of sperm collection frequency on sperm quality has been assessed in teleost species such as trout (Salmo trutta)19,20 turbot (Scophthalmus maximus), 21 European sea bass (Dicentrarchus labrax), 22 and white fish (Coregonus peled). 23 However, it is commonly accepted that an inappropriate sperm collection frequency affects sperm quality 7 and this must be determined for each species. Consequently, the assessment of an appropriate sampling frequency that allows a full recovery of sperm quality in zebrafish is essential for assisted reproduction purposes.
Motility is the most widely studied quality parameter in fish sperm,24,25 and although other analysis are needed to guarantee the status of spermatozoa, it is a useful tool to infer the probability of successful fertilization and to assess the previous mentioned factors.25,26 Still, there are no universal sperm quality biomarkers, therefore, besides motility, other parameters are needed for an accurate quality analysis.6,27 The viability of the plasma membrane is an important feature in spermatozoa since it characterizes the integrity of the cell. 24 Membrane alterations in spermatozoa can affect motility initiation (motility is triggered by membrane signaling), motility maintenance (loss of intracellular adenosine triphosphate [ATP]), and the ability of the sperm nucleus to produce the first embryonic cell after fertilization.26,28,29 In this way, analysis of sperm quality is a useful tool to select the most appropriate conditions to collect zebrafish sperm for cryopreservation and assist reproduction purposes.
This study aimed to characterize the optimal age for sperm collection in zebrafish and to evaluate the effect of the frequency of noninvasive sperm sampling on motility and plasma membrane viability.
Materials and Methods
Zebrafish maintenance
Zebrafish AB (wild type) and Tg(runx2:eGFP), 30 with an AB background, were housed in a standard aquatic recirculation system (Zebtec®; Tecniplast) with 980 L of water, containing a biological filter (ceramic beads), mechanic filtration (50 μm), granular activated carbon filter, and ultraviolet sterilization (180,000 μWs/cm2) to maintain water quality. The water temperature (28°C ± 0.5°C), conductivity (750 ± 70 μS) and pH (7.5 ± 0.2) were constantly monitored through automatic probes, and water was partially replaced daily (10%). The fish room had a controlled photoperiod with a 14:10 h light:dark cycle, an independent air conditioning system (26°C ± 1°C) and an air extraction system to guarantee the air renewal in the room, maintaining the humidity close to 60%. Males and females were maintained separately in 3.5 L tanks. The fish were fed twice a day ad libitum with ZEBRAFEED® (SPAROS Lda., Olhão, Portugal) and Artemia nauplii (AF 480; INVE). Unconsumed food and fish debris were removed daily.
All animal manipulations were performed in compliance with the Guidelines of the European Union Council (86/609/EU), according to the directive's for protection of animals used for scientific research (2010/63/EU) and transposed to the Portuguese law for the use of laboratory animals on research (Decreto Lei n° 129/92 de 06 de Julho, Portaria n° 1005/92 de 23 de Outubro). All animal protocols and fish sampling procedures were performed by licensed researchers (Decreto Lei n°113/2013 de 7 de Agosto).
Sperm collection and quality analysis
On the day before sperm collection (16 h before the sampling), 31 males and females were placed in 1 L breeding tanks at a 1:1 sex ratio (Tecniplast) and maintained separately while sharing the same water, to promote hormonal stimulation for improved release of gametes. Males were anesthetized with 0.168 mg/mL of tricaine methane-sulfonate solution (MS-222; Sigma-Aldrich) prepared according to Westerfield, 32 and sperm was collected (1 h after the lights turn on) by abdominal massage using a glass capillary tube connected to a mouth piece. Sperm was immediately diluted with 10 μL of sterilized and filtered (0.20 μm) Hank's Balanced salt solution (HBSS) at 300 mOsm/kg, 32 to prevent motility activation, in accordance with previous studies. 33 After sperm collection, the samples were maintained at 4°C in the dark until quality analysis was performed (between 1 and 2 h after collection). Meanwhile, the males recovered from the anesthesia in clean system water and were returned to the rearing tanks. Sperm motility was evaluated using Computer Assisted Sperm Analysis (CASA; Proiser). Only samples with concentration higher than 3 × 107 sperm cells/mL were analyzed. 31 To evaluate motility parameters, 0.5 μL of sperm at room temperature was placed on a Makler chamber under a 10 × negative phase contrast objective (Nikon E200, Tokyo, Japan) and immediately activated with 5 μL of filtered and sterilized system water at 28°C ± 1°C. Motility was recorded every 10 s postactivation, during 1 min for each sample. The images were captured with a Basler camera A312f (Basler Afc). Total motility (TM, %), progressive motility (PM, %), curvilinear velocity (VCL, μm/s), straight line velocity (VSL, μm/s), and linearity (LIN, %) were determined to assess sperm quality. Only those spermatozoa with VCL >10 μm/s were considered motile.
To evaluate spermatozoa membrane viability, the percentage of viable cells was quantified using the fluorescent dyes propidium iodide (PI; Invitrogen) and SYBR 14 (Invitrogen). Before the addition of the fluorescent dyes, the sperm sample was rediluted (1:10) in HBSS, to reduce cell concentration. Incubation with 5 μM SYBR 14 and 220 μM PI was performed in the dark at 4°C for 5 min. Cell viability was quantified under an epifluorescence microscope (Nikon E200), equipped with triple excitation filter block DAPI-FITC-Texas Red (excitation filter wavelengths: 395–410 nm [bandpass, 403 center wavelegnt (CWL)], 490–505 nm [bandpass, 498 CWL], and 560–580 nm [bandpass, 570 CWL]). Dead cells with disrupted membrane labeled in red (PI stained cells) and live cells labeled in green (SYBR 14 stained cells) were counted, and the percentage of viable cells was determined. At least 100 cells per slide were counted, and 2 slides per sample and per condition were observed.
Effect of age and sperm collection frequency on sperm quality
Males from wild-type (AB N = 90) and Tg(runx2:eGFP N = 85) zebrafish lines were sampled for sperm collection as previously described. To study the effect of age, sperm motility index was evaluated from: (1) AB line at 6 (N = 26), 8 (N = 13), 12 (N = 27), and 14 (N = 24) months of age; (2) Tg(runx2:eGFP) line at 6 (N = 23), 8 (N = 17), 12 (N = 18), and 14 (N = 27) months of age. The analysis of membrane viability was performed for the same males: (1) AB line at 6 (N = 20), 8 (N = 13), 12 (N = 27), and 14 (N = 21) months of age; (2) Tg(runx2:eGFP) line at 6 (N = 23), 8 (N = 12), 12 (N = 18), and 14 (N = 27) months of age.
Sperm collection frequency was evaluated in terms of motility in AB (N = 90) and Tg(runx2:eGFP) (N = 85) zebrafish lines. From the 90 AB and 85 Tg(runx2:eGFP) males, a total of 78 (AB) and 69 [Tg(runx2:eGFP)] males had sperm samples above 3 × 107 sperm cells/mL and were used to establish 3 treatment groups [AB N = 26; Tg(runx2:eGFP) N = 23]. The first group was sampled 2 days after the first sampling [AB N = 13; Tg(runx2:eGFP) N = 17]. The second group was sampled after 7 days of rest [AB N = 26; Tg(runx2:eGFP) N = 18]. The third group was sampled after 14 days of rest [AB N = 24; Tg(runx2:eGFP) N = 23].
The analysis of membrane viability was performed for the same males in: (1) AB line at the first sampling (N = 23), 2 (N = 18), 7 (N = 23), and 14 (N = 11) days of rest; (2) Tg(runx2:eGFP) line at the first sampling (N = 22), 2 (N = 13), 7 (N = 22), and 14 (N = 22) days of rest. Not all the samples analyzed for motility were analyzed for membrane viability due to the low sperm volume.
Sperm motility parameters of the individual males were assessed through CASA system, every 10 s postactivation during 1 min, to determine TM, PM, VCL, VSL, and LIN. Viability of the plasma membrane was evaluated as previously described.
Data analysis
Due to the high number of variables related to sperm motility measured for each sample (5 motility parameters × 6 postactivation times = 30 motility-related variables), we started by evaluating their degree of redundancy. Principal Component Analysis (PCA) was used to assess whether it was possible to aggregate all 30 variables into a small number of variables, without significant information loss. After a preliminary exploratory analysis, we observed that the LIN variables displayed very low variance, except for sperm samples with extremely low motility and no particularly relevant linear correlation with the other variables. As such, we have no longer considered the LIN parameters for analysis. In contrast, all other parameters (TM, PM, VCL, and VSL) displayed a high degree of positive correlation among them, which was reflected by the fact that it is possible to aggregate these 24 variables into a single variable (PC1), which still retains 52% of observed variation (after mean-centering and autoscaling of these variables, to ensure that PCA does not give preference to higher variance variables) and which can be interpreted as a general “motility index.”
This motility index (i.e., the first component of the PCA) consisted of a weighted mean of these motility measurements (after standardization), which was used for further analysis of variance (ANOVA).
SPSS 18.0 software was used for statistical analysis. Data are expressed as means ±95% of confidence intervals of the mean, and normalized by arcsine transformation when results were expressed as percentages. Statistical differences between treatments were detected by ANOVA and Student–Newman–Keuls (SNK) multiple comparison post hoc tests (p < 0.05). A three-way ANOVA (SNK, p < 0.05) was performed on all motility (PC1) data. For each zebrafish line, a two-way ANOVA (SNK, p < 0.05) was applied to evaluate the effect of age and sperm collection frequency on sperm quality.
Results
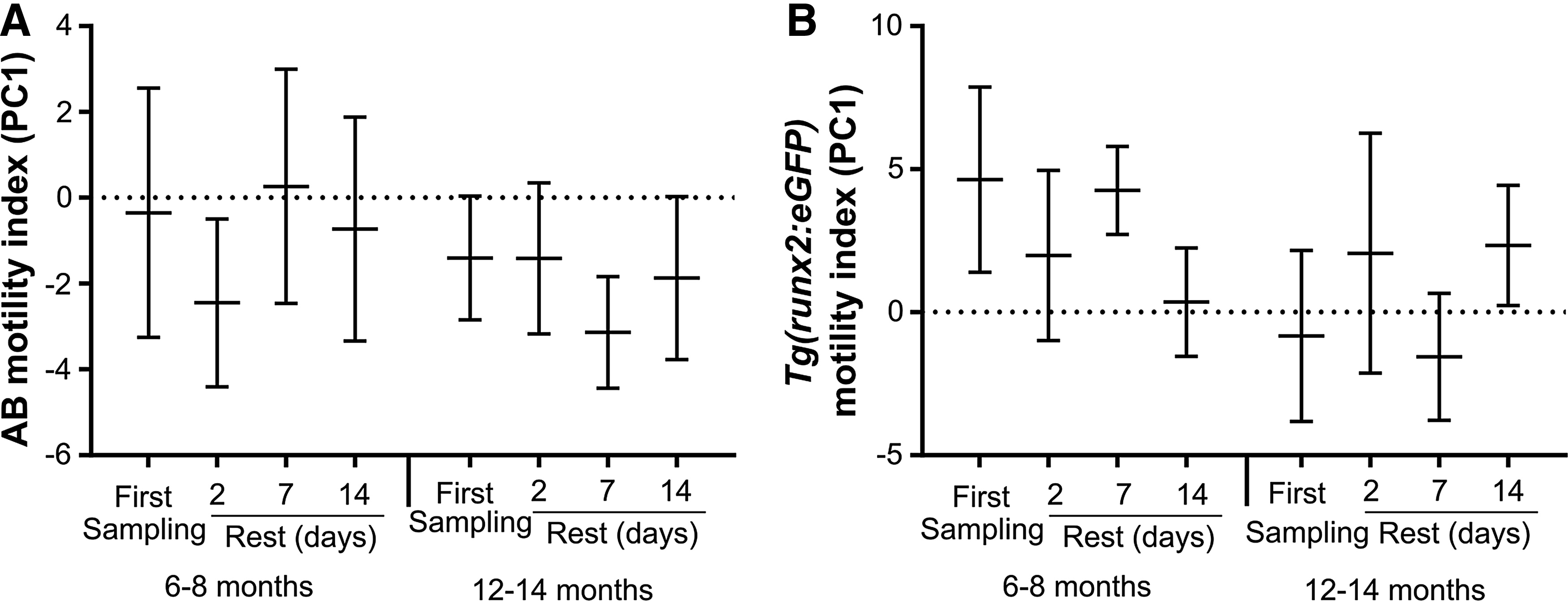
Sperm motility parameters from CASA analysis are presented in Supplementary Tables S1 (AB) and S2 [Tg(runx2:eGFP)], where we could observe a decrease of motility with aging and with low poststripping recovery times. A three-way ANOVA (SNK, p < 0.05) was performed on motility index (PC1) data, which showed that the zebrafish line was the factor with largest main effect on motility (Table 1), since Tg(runx2:eGFP) had significantly higher sperm motility when compared to AB (Figs. 1A, 1B, 2A, and 2B). Given the high number of significant interaction effects observed between factors (line, age, and stripping frequency) (Table 1), which impair interpretation of the main treatment effects, a two-way ANOVA (SNK, p < 0.05) was applied for each line independently, to study the effects of age and stripping frequency (along with possible interactions between these factors). Both lines showed a consistent main effect of age, where younger males (6 and 8 months) had significantly higher sperm motility when compared with older males (12 and 14 months) (Fig. 1A, B). The results of stripping frequency on sperm motility were also consistent in both lines, with no significant effect being observed (Table 1 and Fig. 2A, B). In both lines, there was an age interaction with frequency effect, which means that stripping frequency had an effect on sperm motility that depends on age (Fig. 3A, B).
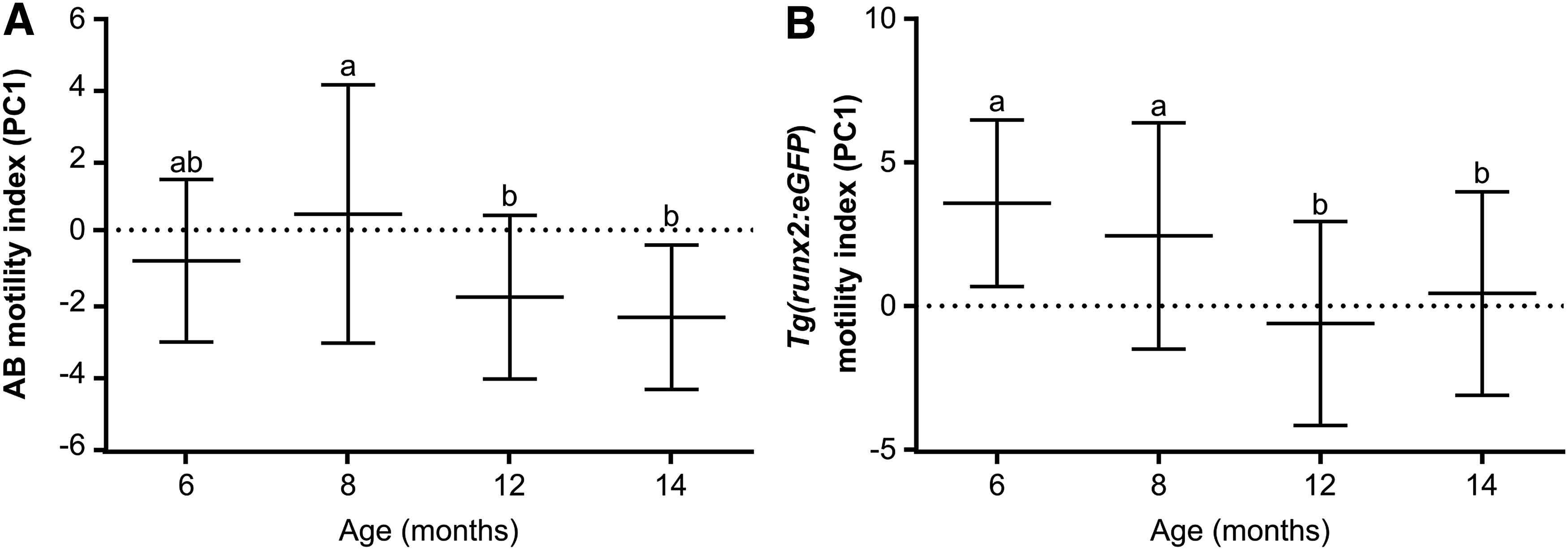
Zebrafish sperm motility index (PC1) according to age of:

Zebrafish sperm motility index (PC1) after different sperm stripping frequencies. The number of the stripping frequency is related to the time of rest between sperm collections. After the first sampling sperm was collected after 2, 7, and 14 days of rest between samplings. The analysis was performed in:
Effect of the interaction in zebrafish sperm motility index between males age and sperm stripping frequency in
Zebrafish Sperm Motility Analysis of Principal Component Related to Age, Stripping Frequencies, and Their Interactions in AB and Tg(runx2:eGFP) Line
Significant differences (three-way ANOVA [SNK, p < 0.05]) are represented.
ANOVA, analysis of variance; SNK, Student-Newman-Keuls.
The percentage of viable cells was analyzed with a three-way ANOVA (SNK p < 0.05) after arcsine transformation, with the factor zebrafish line displaying no effect on sperm viability (Table 2). However, the high number of interaction effects impaired a clear interpretation of the effect of age and stripping frequency on sperm viability, so, as previously, a two-way ANOVA (SNK, p < 0.05) was applied to each line independently to study the effect of age and stripping frequency. Sperm viability in the AB line showed no significant differences between all the studied ages (Fig. 4A, B), while in the Tg(runx2:eGFP) line a significantly higher sperm viability was observed at 8 months of age. In the AB line, males sampled for the first time had significantly higher sperm viability when compared to samples collected 2 and 7 days after first stripping, but it was not significantly different from 14 days after stripping (Fig. 5A, B). Consequently, AB males were able to recover membrane viability 14 days after stripping. On the contrary, Tg(runx2:eGFP) males were not able to recover membrane viability 14 days after stripping, since males in the first sampling point had significantly higher sperm viability compared with males after 7 and 14 days of recovery (Fig. 5B).
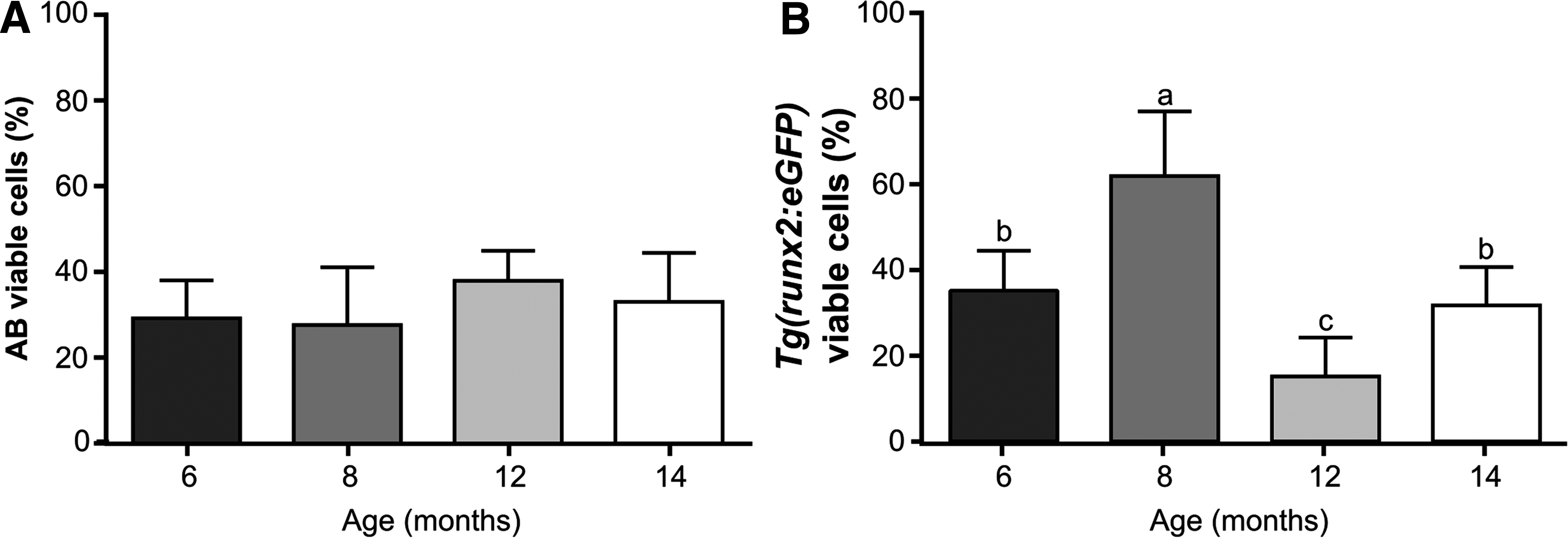
Zebrafish sperm viability (%) according to age of:
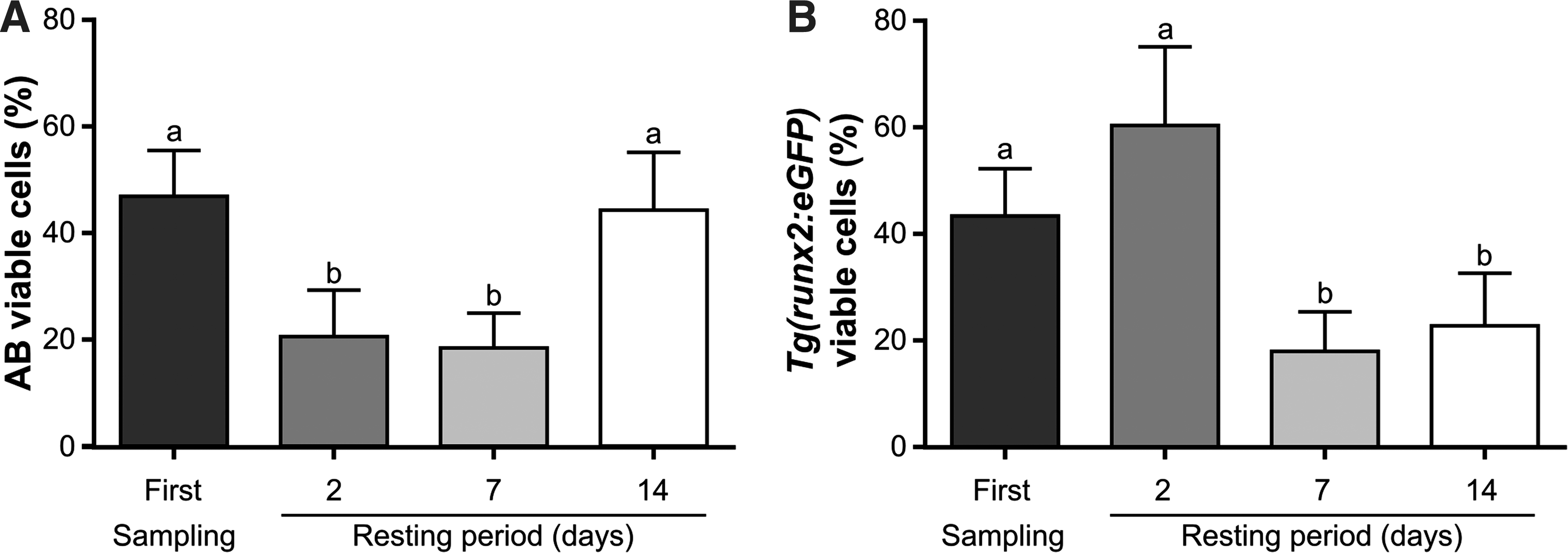
Zebrafish sperm viability (%) after different sperm stripping frequencies. The number of the stripping frequency is related to the time of rest between sperm collections. After the first sampling sperm was collected after 2, 7, and 14 days of rest between samplings. The analysis was performed in:
Zebrafish Sperm Membrane Viability Analysis Related to Age, Stripping Frequencies, and Their Interactions in AB and Tg(runx2:eGFP) Line
Significant differences (three-way ANOVA; SNK, p < 0.05) are represented.
Discussion
The selection of sperm donors is highly relevant for cryopreservation, since it can help in reducing post-thaw variability in zebrafish sperm used for in vitro fertilization. In captivity, zebrafish has a natural longevity of 42–66 months, depending on reproductive effort and caloric intake. 34 In our study, younger zebrafish males (6–8 months) showed significantly higher sperm motility when compared with older males (12–14 months). In agreement, Johnson et al. 35 observed that older zebrafish males had a decline in sperm production and motility, although displaying higher offspring survival. In guppy, male age affected negatively sperm morphology, velocity, and sperm number, but not membrane viability. Older males had slower sperm, with longer flagellum and higher sperm volume than younger males, 10 but these differences did not affect sperm competition success when compared with younger males. It is interesting to observe that both in zebrafish and in guppy, there is a decrease in sperm motility in older males, which might be associated to the accumulation of oxidized proteins and a decline in mitochondrial functionality. 36 Mitochondria aging-related alterations are among the most remarkable features observed in senescent cells. Mitochondrial oxidation is the major source of oxidation lesions accumulated with age, affecting mitochondrial energy metabolism, which is essential for male reproductive function. The damage produced by excessive reactive oxygen species in the sperm membrane causes reduced sperm motility and impairs its ability to fuse with the oocyte. 37 Oxidative stress reduces sperm motility and viability4,38–40 and could be one of the possible explanations for the lower motility in older zebrafish males.
Zebrafish are hierarchical fish with dominant subordinate relationships, which are related with body size and levels of aggression, associated to reproductive success. 41 The hierarchical relationships established among zebrafish are strongly connected to sperm competition, which is a postcopulatory selection that occurs when females breed with multiple males in the same reproductive episode. In this process, sperm from rival males compete to fertilize the oocytes. 42 In sperm competition for fertilization, there is a strong selection for spermatozoa quality parameters that enhance fertilization success, such as sperm quantity and quality.43,44 Therefore, the reproductive setup established to determine sperm quality is extremely important since changes in the social environment rapidly affects sperm competition and therefore sperm quality. Zajitschek et al. 45 found that under high sperm competition environment (two males and one female), males display higher sperm motility and velocity than reproductive setups, where one male was available to two females (low sperm competition environment). In our study, males of each age were permanently maintained separated from females and the reproductive setup was established in breeding tanks in a sex ratio of 4:4 to stimulate reproduction and collect the sperm on the following day.
Therefore, the results obtained with our experimental design emphasize the effect of age and sperm collection frequency in sperm quality, reducing biases associated to sperm competition effects.
Considering this information, it largely explains the differences observed between studies on the effects of zebrafish age on reproduction and sperm quality. Not only are the samples highly heterogeneous but they also manifest adaptations and different investments in gamete production according to the social environment and hierarchical relationships. A characterization of the effect of zebrafish age on sperm competition should be undertaken in the future.
Spermiogenesis is a complex and highly regulated process, where diploid cells called spermatogonia proliferate and differentiate onto mature spermatozoa through mitosis, meiosis, and spermiogenesis. 46 Zebrafish spermatogenesis has a cystic pattern with one of the teleosts fastest spermiogenesis cycle taking only 6 days to reach spermatozoa full maturation. 46 Reinardy et al. 47 observed that with stripping frequencies with a maximum of 7 days of rest, the DNA integrity was not altered, despite the fact that sperm concentration was affected. Consequently, it was necessary to determine the adequate stripping frequency that allows the full recovery of sperm quality. Our data showed that stripping frequency does not affect sperm motility, although it does affect membrane viability. The AB zebrafish line was able to recover the initial membrane viability after 14 days of rest. However, this recovery was not observed in the transgenic line, where sperm membrane viability was still decreased after 7 and 14 days of rest.
In our study, recovery time in younger fish is seemingly faster than in older fish, which is observed through the interaction effects of age and stripping frequency: older fish display lower sperm motility when the recovery time between collections is short, while sperm motility for younger fish seems insensitive to the recovery time between collections. Consequently, it is highly advisable to respect 14 days of rest between sperm collection events, particularly in older fish. The fact that younger fish are less susceptible to cellular distress, related to sperm collection events, reinforces the selection criteria of using younger fish for sperm collection.
The experimental design used in our work allowed a better comprehension of the interaction between treatments that would not be possible otherwise. Both zebrafish lines are commonly used in zebrafish facilities and the transgenic model was used as comparison between wild-type fish breeders with genetic modified zebrafish lines. The Tg(runx2:eGFP) line has an AB background and expresses the Tg(runx2:eGFP) transcription factor, which is related not only to osteoblast differentiation but also to the regulation of cell proliferation. Although most studies on zebrafish sperm cryopreservation and assisted reproduction are performed with wild-type lines, its application is most useful in transgenic and mutant lines, and their particularities are generally unknown or disregarded.
Throughout our experiments, the Tg(runx2:eGFP) transgenic line had systematically higher sperm motility when compared to AB line. The fact that Tg(runx2:eGFP) fish displayed significantly higher sperm motility, but lower capacity to recover membrane viability at 7 and 14 days after sperm collection, suggests the existence of relevant differences between zebrafish lines in terms of sperm quality and susceptibility to damage. Consequently, each zebrafish transgenic and mutant line should be investigated before the establishment of sperm cryopreservation programs.
The knowledge obtained by this work allows the determination of suitable zebrafish age and sperm collection frequencies to obtain the highest sperm quality possible to facilitate cryopreservation procedures, respecting the 3R's principle. Therefore, we consider that males between 6 and 8 months of age have the highest sperm quality, and at least 14 days of rest should be respected between sperm collection events.
Footnotes
Acknowledgments
P.D. acknowledges the financial support from the Portuguese Foundation for Science and Technology (FCT) through the doctoral grant SFRH/BD/97466/2013. This work was partly funded by projects: ASSEMBLE+ JRA2-H2020-INFRAIA-2016-2017 (N730984), EBB-EAPA_501/2016 (Interreg Atlantic Area), PEst-C/MAR/LA0015/2011 (FCT and European Commission-ERDF-COMPETE), and UID/Multi/04326/2013 (FCT). The authors acknowledge the technical support of Filipa Beça, Isa Quinzico and Rita Nogueira during the samplings; and Matthew Castaldi for article revision.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.