
Abstract
Animals communicate with each other through a variety of behavioral interactions, many of which are often complex due to the interplay of several ecological factors. Observations on dyadic interactions can help throw light on the more complex interactions observed among group living organisms and can help in understanding mechanisms of behaviors related to mating strategies, dominance hierarchies, and decision-making. This study focused on the assessment of several generally observed interactions among dyads of different sexes (female-female, male-male, and male-female) in wild zebrafish (Danio rerio). Temporal dynamics of these interactive behaviors were observed in 45 dyads across 3 time intervals of the day. We used generalized linear mixed models to investigate the effect of time, sex of dyad, and their interaction on specific behaviors. While the frequency of occurrence of some behaviors showed clear variation across time intervals of the day, these were further found to depend on the composition of the dyad. Contrary to previous reports, we found that same-sex dyads are equally aggressive and aggressive interactions did not vary temporally. Mating-associated interactions, as expected, were significantly higher in mixed-sex dyads and declined significantly from early morning to afternoon. Interestingly, we also found some mating-associated interactions in same-sex dyads. A fine line exists between social and mating-associated interactions in many organisms and so we speculate that these interactions could also be social interactions and not mating-related behavior. Our findings shed light on complex interactive behaviors among zebrafish, that are likely to be affected by time as well as sex composition of interacting individuals and thus has important implications for groups varying in sex ratios in the wild.
Introduction
Across the animal kingdom, a reduced risk from predation and an increased foraging efficiency driven by improved foraging success among group members has lead to the evolution of group living in many species. 1 It has been observed in several taxa that animals who live in groups enjoy antipredator benefits and improved foraging as well as additional benefits such as greater mating success and faster information transfer. 2 Within groups, a diverse repertoire of interactions allow information exchange that are vital for the survival of the group3,4 Such interactions are complex, dynamic, and give rise to a social structure within the group.5–7
While agnostic interactions give rise to dominance hierarchies, mating-associated interactions facilitate mating. Thus, interaction patterns among organisms within groups are likely to change across time, based on immediate needs of the organisms and surrounding ecological contexts. Ethograms describing such interactions provide insight into interaction patterns across animal societies.8,9 Here, using detailed ethograms, we studied wild zebrafish dyads of different sexes and across three time sessions over the course of a day to understand the dynamics of interaction patterns across time. We followed Kalueff et al.'s catalog for zebrafish behavior 10 and identified all behaviors visible to the naked eye.
While it is understood that there were several survival and other indirect benefits, there are costs associated with group living as well.11,12 A balance between costs and benefits decide grouping tendencies. To perform collective tasks, groups rely on behavior of individuals and interactions within members. Studies on group behavior in stickleback (Gasterosteus aculeatus) have shown that differences in individual behavior within groups drive collective behavior. 13 In mosquito fish (Gambusia holbrooki), individuals adapt to the larger groups by deviating from an individual's characteristic locomotion behavior, and thus opt for grouping. 14 In female baboons (Papio hamadryas), the tendency to groom changes depending on whether an individual is isolated (where it is an individual's grooming tendency) or is a part of a larger group (where it depends on the recipients response). 15 Thus, despite the fact that interactions in groups are made up of many dyadic interactions, group interactions are considerably different from interactions within dyads. Indeed, social behaviors such as shoaling tendencies have been shown to be different in zebrafish dyads when compared with shoals (comprising five individuals). 16 However, across taxa, dyadic interactions have widely been studied to understand the nature of interactions. Individuals, dyads, or small groups are easier to observe, and from such observations, we get some insight into behavior of groups in the wild. Specific details of interindividual behaviors are best observed in dyads. Dyads are widely studied to understand interactions. While typically, male-female (MF) dyads are studied to gain insight into the species' mating behavior,17–19 studies on aggression involve same-sex dyads (mostly male-male [MM] dyads).20–22
In fish dyads, aggressive behavior may involve indirect (noncontact) and direct (contact) interactions. Such interactions give rise to dominance hierarchies.23–27 Studies across several fish species have examined whether winners of agnostic encounters are more successful in courtship compared to losers, and furthermore, these could be correlated.28,29 Fish species have diverse mating strategies and often show diurnal or seasonal patterns in mating.24,30 Some studies characterizing courtship and mating interactions in fish species have been conducted within mating dyads and this has allowed to tease apart the discrete mating-related behaviors in the species.31,32
Zebrafish are small shoaling cyprinids found in stagnant to slow-flowing streams of India, Bangladesh, and Nepal. 33 In the wild, shoal sizes in Danio rerio can vary vastly across different habitats and have been observed to be between 5 and 500 individuals. 33 Shoaling commences from the larvae stage (∼12 days postfertilization) and zebrafish shoals can vary in cohesion depending on the predation pressure and food availability in their habitats. 34 Even as remarkable variation observed in social behavior among zebrafish shoals highlight the important role played by ecological factors, the exact mechanisms driving these variations and the determinants of social behaviors such as shoaling decisions are still waiting to be unraveled.
Interactions among individuals within zebrafish shoals could comprise interactions between two males, two females, and between a male and female. A variety of interactive behaviors can be observed between dyads. For instance, unlike what is commonly examined, aggressive interactions occur between dyads comprising of MF, or female-female (FF) individuals, and not just between MM dyads. Aggressive behavior involves biting, chasing, and can lead to formation of dominance hierarchies in both sexes. 35 Dyad studies have also identified courtship interactions in this species and have categorized courtship in this species into three phases: initiatory, receptive or appetitive, and spawning. 31
Here, we explored whether the sex of the dyad has an impact on the nature of interactions. An earlier related investigation on the temporal (diel) dynamics across mixed-sex dyads in this species revealed changes in the intensity of behavior across diel sessions. 36 Like Ghoshal et al., we also examine interactions between dyads over time (i.e., diurnal studies). Thus, our work focuses on unraveling differences in interactive behaviors based on composition of dyads over the course of a day. Unlike most studies, which address questions on specific interactions across dyads of particular sexes, this study aimed to address a broader question. Here, we examined behavioral interactions across three kinds of dyads (comprising of different sexes) over three discrete time sessions within a day in wild zebrafish (D. rerio), to investigate temporal patterns in same and mixed-sex dyads of this species. Specifically, the main objectives of this work were (1) to compare dyadic behavioral interactions among mixed and same-sex zebrafish dyads and (2) to explore temporal variations in interaction patterns within different kinds of dyads.
Methods
Maintenance of fish
For our study, adult wild zebrafish from Howrah district (West Bengal, India) was obtained from a local collector. In West Bengal, zebrafish populations occur in shallow water habitats varying in vegetation and water flow and have been found to differ in shoaling and aggressive behavior. 37 Fish were maintained in 30 × 30 × 60 cm glass aquarium stock tanks filled with aged, filtered water (through a faucet iron filter) of pH ranging between 7.5 and 7.75 and water conductivity ranged between 480 and 520 μS/cm. The holding tanks were kept at 22°C–24°C under a constant lighting (simple 12 W LED Light Panels) condition of 12 h dark:12 h light and fish were fed freeze-dried bloodworms once daily in the morning. Mixed-sex groups of ∼30 individuals were maintained in each such tanks and acclimatized in the above mentioned conditions for 3 weeks before the experiment.
Experiment
Three kinds of zebrafish dyads, MM dyads, MF dyads, and FF dyads, and a total of 45 dyads (n = 15 of each dyad type) were used in this study. On the day of the experiment, a zebrafish dyad was introduced to a 30 × 20 × 20 cm experimental tank with a substrate (a Petri plate with pebbles). In the wild, zebrafish have been observed to spawn preferentially in habitats with gravel substrata, as it provides protection to eggs and is known to increase egg survival. 38 The presence of a substrate has been shown to be important to fish for specific mating-related behaviors such as circles and leads. 36 As gravel substrate is preferred over silt substrates, 38 we provided pebbles placed on Petri plates as substrata. After 10 min of acclimatization to the tank, the dyad was video recorded (Sony DCRSX22) over three sessions, early morning (EM) from 9:40 AM to 10:40 AM, morning (M) from 11:40 AM to 12:40 PM, and afternoon (A) from 1:40 PM to 2:40 PM. 36 The first session (EM), beginning at 9:40 AM, was 15 min after the onset of “dawn” as lights turned on routinely at 9:25 AM in the laboratory. Each dyad was tested only for one set of sessions, after which individuals were returned to the stock tanks.
Zebrafish exhibit a wide repertoire of behaviors, and these behaviors have been categorized into anxiety-related behaviors, social behavior, sexual behavior, sleep behavior, and so on. 10 We followed Kalueff et al. and Ghoshal et al., and identified five distinct behaviors: leads, nudges, bites, chases, and circles. Our descriptions of individual behaviors are consistent with Ghoshal et al.. To prevent a possible misinterpretation of behaviors, we additionally measured the speed of individuals to distinguish between leads and chases. We calculated displacement (and thereby speed) of 120 interactions (leads or chases) in the X-Y plane, using an image-editing software (ImageJ 39 ). We established that chases have higher speeds (mean ± SE, 4.17 ± 0.23 cm/s) compared to leads (mean ± SE, 2.78 ± 0.18 cm/s). There was a significant difference (unpaired Wilcoxon test, W = 933.5, n = 60, p < 0.001) in speeds between these two kinds of interactions. Speeds varied based on the individuals constituting the dyad and on the sex of the dyad. No speed cutoff was set and in instances where the speed indicated either of the interactions (i.e., leads or chases), leads and chases were distinguished based on the observation that chases involved acceleration of individuals and often end in bites (pers. observation). Similarly, bites and nudges were additionally distinguishable as bites involved opening of the mouth (unlike nudges).Ten random 2-min time slots within each video (from 0:00 to 1:00) were analyzed. Occurrences of the selected behaviors observed in the time slots were counted and recorded manually. Behaviors of both individuals in the dyad were counted. All video analyses were carried out by the same observer, blind to the dyad sex and the session, to avoid any bias in recording counts. Following Ghoshal et al., we classified interactions into two categories. In our first classification, we combined interactions based on the nature of interactions; bites and chases as aggressive interactions and leads and nudges as mating-related interactions. Circling, a somewhat rare interactive behavior, is observed where individuals in the dyad move around each other in a circular pattern. 10 This behavior was not included in either category as it was sometimes aggressive and sometimes mating related, and it is often difficult to distinctly categorize this behavior as either and so, excluded in the analyses involving these categories, but was considered separately. As an additional investigation, we grouped the behaviors into a second classification based on the tactility of interactions; nudges and bites were considered contact behaviors, and chases, leads, and circles were considered as noncontact behaviors.
The study complied with the existing rules and guidelines outlined by the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), Government of India. The study adhered to the Institutional Animal Ethics Committee's (IAEC) rules and guidelines of IISER Kolkata. No animals were euthanized or sacrificed during any part of the study, and behavioral observations were conducted without any chemical treatment on the individuals. At the end of the experiments, no individuals were sacrificed and all were returned to stock tanks and continued to be maintained in the laboratory.
Statistical analysis
All statistical analyses were performed using R, version 4.0.2. 40 Generalized linear mixed models (GLMMs) were built using lmerTest package. 41 Separate models were built to predict each measured behavior, with the measured behaviors (individual or combined) as the dependent variables and dyad identity (fish id) incorporated as the random factor. Observation session (EM, M, A), dyad sex (FF, MM, MF), and their interactions (session × sex) were included as fixed factors. Model comparisons were performed using “Anova” in “car” package 42 and post hoc comparisons (Wilcoxon tests with Bonferroni corrections for multiple comparisons) were carried out for comparing effects of factors that were significant. Models were built for all behaviors together, behaviors individually and for the behavioral classes (i.e., combined behaviors) described in the Methods section. The level of significance was p < 0.05 for all statistical tests.
Results
The GLMMs conducted to predict aggression indicated that there was a significant effect of sex of dyad on aggressive interactions (bites and chases were combined together as aggressive interactions) (Wald type II χ 2 = 6.25, df = 2, p = 0.04; Table 1). When modeled alone, bites depended significantly on the sex of the dyad (Wald type II χ 2 = 6.17, df = 2,p = 0.04), while chases did not (Wald type II χ 2 = 5.58, df = 2, p = 0.06). Details of these models are given in the Supplementary Data S1. Mating-associated interactions (leads and nudges were combined as mating-associated interactions) depended significantly on the sex of the dyad (Wald type II χ 2 = 19.25, df = 2, p < 0.001), session (Wald type II χ 2 = 52.5, df = 2, p < 0.001), and their interaction (Wald type II χ 2 = 25.2, df = 4, p < 0.001; Table 2). GLMM model for leads taken separately depended significantly on the sex of the dyad (Wald type II χ 2 = 45.27, df = 2, p < 0.001), session (Wald type II χ 2 = 37.61, df = 2, p < 0.001), and their interaction (Wald type II χ 2 = 31.85, df = 4, p < 0.001), while GLMM for nudges when taken separately depended significantly only on the session (Wald type II χ 2 = 22.4, df = 2, p < 0.001). Details of these models are given in the Supplementary Data S2. Like nudges, circles also depended significantly only on the session (Wald type II χ 2 = 12.96, df = 2, p < 0.01; Table 3).
Generalized Linear Mixed Model Showing Estimates, Standard Errors, Degrees of Freedom, t Values and p-Values for Combined Aggressive Interactions (bitespluschases)
Indicates significance at p < 0.001, and *indicates significance at p < 0.05.
Df, degrees of freedom; EM, early morning; MF, male-female; MM, male-male; Pr, p-value.
Generalized Linear Mixed Model Showing Estimates, Standard Errors, Degrees of Freedom, t Values and p-Values for Mating-Associated Interactions (leadsplusnudges)
indicates significance at p < 0.01, and *indicates significance at p < 0.05.
Generalized Linear Mixed Model Showing Estimates, Standard Errors, Degrees of Freedom, t Values and p-Values for Circles
Aggressive interactions
In general, aggression within dyads differed based on sex composition of the dyads. Aggressive interactions were significantly more in FF (mean ± SE, 152.13 ± 13.82) than in MF dyads (mean ± SE, 84.75 ± 11.48) (Wilcoxon test, W = 538, n = 45, p < 0.001) and also significantly more in MM (mean ± SE, 141.88 ± 16.05) than in MF dyads (mean ± SE, 84.75 ± 11.48) (Wilcoxon test, W = 633.5, n = 45, p < 0.01). Aggressive interactions between FF and MM dyads were comparable (Wilcoxon test, W = 904, n = 45, p = 0.38) (Fig. 1). When considered separately, both bites and chases among dyads were found to show similar patterns (Supplementary Data S1).

Comparison of aggressive interactions (bites and chases) exhibited by zebrafish dyads of different sexes. MM and FF dyads show comparable aggressive interactions. MF dyads show significantly lower aggressive interactions. Statistically significant differences (p < 0.05) are indicated by different alphabets (a,b), while nonsignificant differences are indicated by same alphabet. (Comparisons were performed using Wilcoxon unpaired tests, n = 45). FF, female-female; MF, male-female; MM, male-male.
There was, however, no significant effect of session on either of the aggressive interactions (i.e., bites and chases), indicating that overall aggressive interactions did not vary vastly across the observed sessions during the day.
Mating-associated interactions
The GLMM conducted for prediction of mating-associated interactions (leads and nudges combined) revealed a significant effect of sex of the dyad, session, and sex of dyad interacting with session (Table 3). Although significantly lower than MF dyads, mating-associated interactions were observed in same-sex dyads as well (for details, see Supplementary Data S2). Mating-associated interactions were significantly more in MF dyads (mean ± SE, 30.08 ± 4.19) than in FF dyads (mean ± SE,14.17 ± 2.39) (Wilcoxon test W = 1398, n = 45, p < 0.01; Fig. 2) as well as in MM dyads (mean ± SE, 10.37 ± 1.47) (Wilcoxon test, W = 1489.5, n = 45, p < 0.001; Fig. 2). Mating-associated interactions were comparable between FF and MM dyads (Wilcoxon test, W = 904.5, n = 45, p = 0.38) (Fig. 2).

Comparison of mating-associated interactions in dyads based on their sex composition (MF, FF, MM). Mixed-sex (MF) dyads showed significantly more mating-associated than either of the same-sex dyads (MM or FF). Statistically significant differences (p < 0.05) are indicated by different alphabets (a,b) while nonsignificant differences are indicated by same alphabet. (Comparisons were performed using Wilcoxon unpaired tests, n = 45).
Mating-associated interactions within dyads were also significantly dependent on the session being observed. There were significantly more mating-associated interactions in EM (mean ± SE, 29.8 ± 4.32) than in M (mean ± SE, 14.28 ± 2.06) (paired Wilcoxon test, V = 780.5, n = 45, p < 0.001), significantly more mating-associated interactions in M (mean ± SE, 14.28 ± 2.06) than in A (mean ± SE, 10.55 ± 1.67) (paired Wilcoxon test, V = 653.5, n = 45, p < 0.01) and significantly more mating-associated interactions in EM than in A session (paired Wilcoxon test, V = 872.5, n = 45, p < 0.001) (Fig. 3). When considered separately, leads and nudges also showed similar patterns of interaction behavior (Supplementary Data S2)
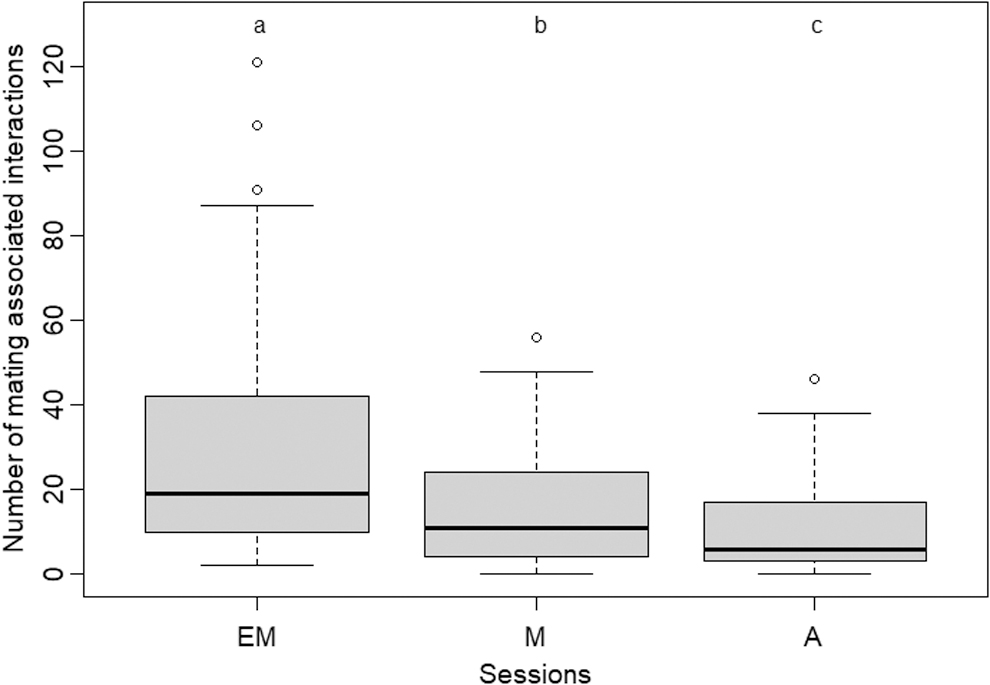
Comparison of mating-associated interactions in dyads across different time sessions (EM: early morning; M: morning; A: afternoon). Mating interactions were found to be highest in the EM session, compared to either of the two later sessions. Statistically significant differences (p < 0.01) are indicated by different alphabets (a,b) while nonsignificant differences are indicated by same alphabet. (Comparisons were performed using Wilcoxon paired tests, n = 45).
For same-sex dyads, there were significantly more mating-associated interactions in EM (mean ± SE, 18.3 ± 2.61) than in A (mean ± SE, 8.07 ± 1.22) sessions (Paired Wilcoxon Test, V = 41, n = 30, p < 0.001). Mating-associated interactions in same-sex dyads were comparable between EM (mean ± SE, 18.3 ± 2.61) and M (mean ± SE, 10.47 ± 1.6) (Paired Wilcoxon test, V = 99.5, n = 30, p = 0.02) and M (mean ± SE, 10.47 ± 1.6) and A (mean ± SE, 8.07 ± 1.22) sessions (Paired Wilcoxon test, V = 110, n = 30, p = 0.09) (see Supplementary Data S2, Supplementary Fig. S6).
In general, aggressive interactions and mating-related interactions among dyads were found to be negatively correlated (Spearman rank correlation test: r = −0.2, p = 0.021).
Circling behavior
Circling behavior was found to vary significantly across sessions (Wald type II χ 2 = 12.96, df = 2, p ≤ 0.01) (Table 3). There was significantly more circling behavior in EM (mean ± SE, 0.73 ± 0.24) than in M (mean ± SE, 0.08 ± 0.04) session (paired Wilcoxon test, V = 164, n = 45, p < 0.01) and A (mean ± SE, 0.20 ± 0.10 (paired Wilcoxon test, V = 120.5, n = 45, p < 0.01) sessions. Circling behavior between M and A sessions were not significantly different (paired Wilcoxon test, V = 5, n = 45, p = 0.28) (Fig. 4).

Comparison of circles exhibited by zebrafish dyads across different time sessions (EM: early morning; M: morning; A: afternoon). The number of nudges significantly decreases from EM to M and EM to A. The number of nudges is comparable between M and A. Statistically significant differences (p < 0.01) are indicated by different alphabets (a,b), while nonsignificant differences are indicated by same alphabet. (Comparisons were performed using Wilcoxon paired tests, n = 45).
Contact interactions and noncontact interactions
Session of observations was found to be the significant predictor factor for contact interactions (Wald type II χ 2 = 7.43, df = 2, p = 0.02) as well as for noncontact interactions (Wald type II χ 2 = 12.24, df = 2, p < 0.01). Details on post hoc comparisons for both these types of behaviors across dyads and sessions are provided in Supplementary Data S3.
Discussion
Our study aimed to understand the dynamic changes in behavioral interaction patterns among wild zebrafish dyads, and detailed observations on behavioral interaction between dyads across different time sessions revealed distinct patterns. While our results agree with the general patterns observed in MF dyadic behavioral patterns in Ghoshal et al., our observations further indicate that dyadic interactions in zebrafish are highly dependent on sex of the individuals constituting the dyads. Aggressive as well as mating-associated interactions were similar between same-sex (MM or FF) dyads, but the interaction dynamics change significantly when the dyad is a mixed-sex (MF) one. As expected, mating-associated interactions were significantly higher in a mixed-sex (MF) dyad. We also observed same-sex mating-associated interactions in zebrafish. In general and consistent with Ghoshal et al. and Hutter et al., we found higher number of aggressive interactions within mixed-sex (MF) dyads compared to other types of behavioral interactions. However, we found that the number of aggressive interactions within same-sex dyads was even more than mixed-sex (MF) dyads. Thus, our study revealed that behavioral patterns among dyads are driven by the sexual composition of the interacting pairs.
Like Ghoshal et al., we observed a distinct temporal pattern in mating-associated interactions. Our results show an equally high number of aggressive interactions across the three sessions. However, our results differed from Paull et al. as we did not observe a drop in aggression across the day, from early morning to afternoon. It is likely that these interactions could change if observations were conducted over a yet later session (for example, late afternoon or evening). Being a diurnal species, we did note a general reduction of activity in zebrafish in the later parts of the day (pers. observation), and therefore, and in keeping with the protocol followed in Ghoshal et al., we did not include this session in our study. As the general set-up of the observation tank (in terms of laboratory lighting, water temperature, basic tank set-up etc.) was similar to the holding tanks, the chance of habituation of the dyads to the observation tank and the possibility of this confounding the circadian activity (and behavioral) patterns is minimal. The recording of the first session (EM) began after a 10 min acclimation period, to avoid any bias due to potential stress of shifting the dyads from the holding tanks.
Aggressive interactions
Interestingly, aggressive interactions were the most frequently observed interactions in the zebrafish dyads. We found 87.29% of all interactions to be aggressive ones. Ecological factors such as vegetation, along with competition for resources, could be a reason for aggression being prevalent. Significantly higher aggressive interactions in same-sex dyads compared to mixed-sex dyads are likely to be due to MF dyads showing 2.5 times more mating-associated interactions (and thus, spending a greater proportion of time on mating-associated interactions).
Contrary to our expectations based on studies with other species and D. rerio, both the same-sex (MM and FF) dyads were found to be equally aggressive. Studies on intrasexual dyadic encounters in cichlids (Cichlasoma dimerus) also indicate males and females to be equally aggressive. 43 Sexually monomorphic electric fish species Gymnotus omarorum have been shown to exhibit similar levels of aggressive behavior irrespective of whether they are males or female dyads. 17 However, contrary to our findings, Paull et al., showed male only (MM) dyads to be more aggressive than female only (FF) dyads in zebrafish. Similar observations across a variety of taxa show that males are generally more aggressive than females.44,45 Environmental context could be an important determining factor for aggression. Aggression in wild zebrafish is known to vary across populations depending on predation pressure and vegetation. 46 While the extent of the role of vegetation on aggression is unclear, a low predation pressure might increase aggression levels as individuals can then set up dominance hierarchies based on aggression. 37 Equally high aggression levels in MM and FF dyads can be because the wild caught fish used in this study occur in habitats with relatively low predation pressure, and thus both sexes are likely to spend their energy on establishing social hierarchies to acquire maximum food resources. 47 In addition to ecological factors, aggression could also be determined by genetic factors. Long-term studies across generations would be needed to establish the role of genetic factors.
Studies on dominance hierarchies in zebrafish where aggression was measured in two sessions within same-sex dyads show aggression levels vary across the day, decreasing from morning to late afternoon. 35 Contrarily, we observed no such drop in aggressive interactions across our observation sessions, with aggression level in early morning session being comparable to those in the late afternoon session. This indicates that dyads in our study continue to be active and spend time foraging across the observed sessions. However, it is possible that observations on interactions conducted beyond our study sessions, perhaps closer to the evening, might have also shown a decline in aggression.
When observed separately, bites and chases also independently follow the same trend. It has been reported that aggression involving noncontact (indirect) behaviors, such as chasing is typical of females,47,48 and in cichlids, aggressive interactions involving physical contacts were greater in males compared to females. 42 In contrast to these findings, our current work with zebrafish shows comparable contact aggressive interactions (bites) and noncontact aggressive interactions (chases) in MM and FF dyads.
Mating-associated interactions
Mating-associated interactions were found to vary across sexes and sessions. Under natural conditions, zebrafish typically breed early in the morning (i.e., dawn) 33 and mating-associated interactions in the early morning session in the laboratory are, therefore, expected.31,33,36
Although mating-associated interactions were significantly higher in MF pairs, some mating-associated interactions were observed also among same-sex dyads. Same-sex mating-associated interactions have been observed in a variety of species, including guppies, dolphins, and chimpanzees.49,50 However, as these interactions drop from EM to A, it is possible that these interactions are exploratory activities and exhibited by the dyads irrespective of the sex of individuals in the dyads. Also, in some species, mating-associated interactions overlap with social behavior 51 and our observation of same-sex mating interactions could point to the fact that the leads and nudges we observed are not mating associated but are social interactions. Further experiments to separate exploratory from mating interactions would be needed in future studies to confirm this.
When observed separately, leads alone also show the same trend as mating-associated interactions. A negative correlation between mating-associated interactions and aggressive interactions found in our study is expected, as spending time in one behavior would decrease the time for the other behavior.
Contact and noncontact interactions
Contact interactions were found to be significantly more in same-sex dyads compared to mixed-sex dyads. This is likely because most contact interactions observed were bites. As aggression was higher in same-sex dyads, and contact interactions were correspondingly higher for same-sex dyads. Also, while contact behaviors vary across same-sex and mixed-sex dyads, noncontact interactions vary across sessions. Decline in noncontact interactions from early morning to afternoon session is likely due to the corresponding large proportion of leads (that comprise noncontact interactions). As mating declines from EM to M, so do noncontact interactions.
While this work examines temporal patterns in interactions in wild zebrafish, studies have been conducted that compare such interactions between wild-caught and laboratory bred strains. Differences among strains have been found to be behavior dependent as found in a study comparing behavior among a laboratory bred strain (Scientific Hatchery or SH strain) and two wild caught populations. 52 In terms of mating, wild caught fish have a greater temporal variation in mating behavior (egg production) compared to a laboratory strain (WIK or wild Indian karyotype). 53 As a further study to understand the genetic basis of these behaviors, it will be interesting to make a detailed comparison of temporal patterns in various behavioral interactions between wild and laboratory-bred strains.
In conclusion, this study suggests that zebrafish dyads can be identified as same sex or mixed sex based on their behavior. Distinct interaction patterns were observed for aggressive and mating-associated interactions across the dyads Our current work thus answers a broad question, and further studies are warranted to extend these studies to understand behavior of individuals within groups. Behavioral observation on groups differing in sex ratios could throw further light on combined effect of multiple individual interactions and effect of sex ratios on changes in group interaction patterns. These studies in the context of ecological factors such as predation or vegetation impact would be helpful in understanding behavior in nature.
Footnotes
Authors' Contributions
A.B. and I.M. designed the study. I.M. conducted the experiments and video analyses. A.B. and I.M. analysed the data. I.M. and A.B. wrote the article. All authors gave final approval for publication.
Acknowledgments
The authors thank the Indian Institute of Science Education and Research Kolkata (IISER Kolkata), India, for providing infrastructural and financial support. Helpful discussions with Aditya Ghoshal, Danita Daniel, and Rubina Mondal during this study are deeply appreciated. The authors also thank Prasenjit Pan for help in fish maintenance in the laboratory.
Disclosure Statement
No competing financial interests exist.
Funding Information
I.M. was supported by a junior research fellowship from IISER Kolkata. This work was supported by the Academic Research Funds provided by Indian Institute of Science Education and Research Kolkata (IISER Kolkata), India to A.B.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.