
Abstract
Accumulating researches suggest that the microbiota reside in the gastrointestinal system can influence neurodevelopment of brain and programming of behaviors. However, the mechanism underlining the relationship between shoals' behaviors and intestinal microbiota remain controversial and the roles of responsible neurotransmitters are still unclear. Here we show that shoaling behavior affected the color preference of shoals, indicating that shoals tended to choose a favorable color environment that benefited social contact. Meanwhile, administration of the selective D1-R antagonist, SCH23390, could disrupt the social interaction that led to the deficits of color preference in shoals. More importantly, the altered microbiota caused by an antibiotic oxytetracycline (OTC) exposure decreased the sociability and weakened shoals' preference for all color combinations. When given a supplementation of Lactobacillus rhamnosus GG after OTC exposure, fish maintained the same capability of social cohesion and color preference as normal fish. Our results support a role for dopamine in shaping the color preference in shoals. Our findings show that dopamine level of brain could mediate both social recognition and color preference, and offer a possibility that the production of dopamine is coordinated through gut microbiota.
Introduction
Sociability that is found across the animal kingdom comprises a wide range of cooperative, neutral, or antagonistic behaviors. 1 Some animals are highly social when they live in cooperative communities (e.g., termites and primates), whereas others are mostly solitary (e.g., bear). Social behavior is observed only when animals gather into a group. Some of these behaviors are beneficial for animals, such as division of labor, cooperative care, shared risk, and increased immunity, whereas others are deleterious for groups, such as conflict, dominance, and coercion. 2 As a vital component of social behavior, the change of social interaction may have negative impacts on the whole fitness of animals. 3
It has been found that a mixture of antibiotics can reduce the social cohesion of zebrafish, resulting in severe tissue damage. 4 Because the antibiotics often negatively affect social interaction, probiotic supplementation can have a beneficial effect. For instance, it has been reported that the social deficit can be reversed through treatment with Lactobacillus reuteri, 5 and zebrafish with the probiotic strain Lactobacillus rhamnosus IMC 501 increase shoaling behavior. 6 In addition, microbes are implicated in determining social cues in animals, and the altered gut microbiota may affect the social interactions between animals. 5 For example, germ-free mice, in which the microbiota is absent, display deficits in social cognition and social recognition.7,8 Accumulating evidences suggest that the mechanism of antibiotic/probiotic administration regulating shoaling behavior may be attributed to the function of the intestinal microbiota.
Animals are usually better able to perceive colors that stand out from their surroundings,9–11 and their color preferences (defined as the ability to discriminate among color stimuli and the tendency to spend more time in particular colors12,13) are either most represented in the environment 14 or contrast with the background. 15 For example, the parasitic wasp prefers yellow, which is the most commonly encountered flower colors. 16 Bumblebees prefer blue flowers that show obvious contrast with the complex background. 17 As is well known, zebrafish are social animals and tend to travel in shoals (by forming and remaining in a tight group called the cohesion of shoals). 18 Meanwhile, social interaction in a shoal is an important behavior for fish to sense danger and stay away from predators; thus, it is beneficial for a fish to stay tight within a shoal. Furthermore, zebrafish have a rich color vision, whose retina contains four different types of cone cells and is capable of processing four regions of the color spectrum (namely red, green, blue, and ultraviolet). 19
In addition, plenty of researches have proven that zebrafish have preference toward different colors (including blue, yellow, red, and green),20–24 although the conclusion remains controversial. For instance, some researches show that zebrafish have a strong preference for blue,20,21 whereas others report a clear aversion for this color.22–24 It is still not clear whether the shoaling behavior affects the color preference. We hypothesis that the characteristic of background color may influence the efficiency of social contact within a shoal and consequently the shoal may prefer certain background colors to keep the social contact. Recently, Park et al. 25 tested the color preference in shoal of zebrafish larvae [5 days postfertilization (dpf)], although shoaling behavior usually starts to develop after 7 dpf and gradually increases with maturity.26–28 As far as we know, few researches have been published on the color preference of juvenile fish (26 dpf or 47 dpf) either, which have strong shoaling behavior.
The cohesion of shoals has been found to be associated with the whole brain dopamine level. 29 Dopamine is one of the major neurotransmitters in the central nervous system of the vertebrate brain that plays important roles in a variety of cerebral functions, such as mood, attention, reward, and memory.30–32 Abundant evidences have shown that dopamine is associated with the neurobehavioral functions in zebrafish.33,34 It has been found that the dopamine and its metabolite 3,4-dihydroxyphenylacetic acid (DOPAC) levels increase in the brain of the adult zebrafish by strong social stimuli, 33 whereas short-term isolation could reduce the DOPAC level of zebrafish. 34 The social interaction of shoals in zebrafish can also be affected by the dopaminergic system through influence of the dopamine level. 35 According to related studies, we selected a D1 dopamine receptor antagonist (SCH23390) to disrupt social preference in zebrafish by reducing dopamine level in the dopamine system.36,37 D1-receptor is one of the most abundantly expressed dopamine receptor subtypes in the brain of zebrafish. 38
In this study, we reveal the influence of antibiotic [oxytetracycline (OTC)] and/or probiotic [L. rhamnosus GG (LGG)] on the social interaction and color preference with zebrafish as a model animal. Meanwhile, we assess the impact of SCH23390 that affects brain dopamine level on the social interaction and color preference. By analyzing the dopamine levels, we establish the correlation between OTC and/or LGG and behaviors through neurotransmitter of the brain. We also evaluate the intestinal microbiota through 16S rRNA sequencing after OTC and/or LGG administration to establish a possible correlation between intestinal microbiota and social behaviors through neurotransmitters in the brain. Further understanding of how intestinal microbiota influence brain will be meaningful for elucidating the causal mechanisms underlying behavior of shoals and for generating new behavioral paradigms for social disorders, such as social anxiety and depression.
Materials and Methods
Zebrafish husbandry and embryo collection
Zebrafish (AB strain) of 4 months old were obtained from breeding center at University of Science and Technology of China, with an average body length of 3.12 ± 0.49 cm and an average body weight of 1.56 ± 0.21 g. Zebrafish were maintained at 28°C ± 0.5°C with a 14 h light/10 h dark cycle (room fluorescent light, 08:00 am–22:00 pm). The pH and conductivity in circulating water of the aquarium were 7.0–7.4 and 1500–1600 μs/cm, respectively. All adult zebrafish were fed fresh brine shrimps twice a day, at 09:00 am and 14:00 pm. Before the experiment, these zebrafish were bred for at most five times, and the embryos were collected from spawning healthy adults. To obtain the normal fertilized embryos for tests, the dead embryos and sundries were cleared. The methods were performed in accordance with relevant guidelines and regulations and approved by the Ethical Committee of Hefei University of Technology.
Antibiotic and SCH23390 exposure, probiotic culture, and exposure
OTC, as a common antibiotic, was purchased from Aladdin (Shanghai, China). It was dissolved in water with a concentration at 100 mg/L as a stock solution. Based on the available literature on the concentration of antibiotic that can affect shoaling behavior, 4 the concentration of OTC used in this study was set at 500 μg/L. The D1-receptor antagonist SCH23390 (R−(+)-8-chloro-2,3,4,5-tetrahydro-3-methyl-5-phenyl-1H-3-benzazepine-1-ol; Cat. No. D054; Sigma-Aldrich) was applied to treat individuals, which significantly reduced the amount of dopamine in the brain of zebrafish. 35 Before experiment, fish were placed in drug exposure beaker and remained in D1-receptor antagonist solution (0.1 mg/L, 500 mL in volume) for 1 h, sufficiently long for the drug to reach the zebrafish brain through the vasculature of their gills and skin. 35 All the animals in the beaker were offered the same conditions (including illumination, temperature, and dissolved oxygen) identical to the standard aquarium.
The probiotic LGG was obtained from Bnbio (Beijing, China), and was cultured in MRS medium at 37°C for 24 h. Then, the LGG in MRS medium was stored at 4°C for no more than 24 h before use. The liquid culture was centrifuged at 10,000 rpm for 2 min at 4°C. The pellet was collected and washed twice with sterile phosphate-buffered saline (PBS) buffer and resuspended. The final concentration of LGG was 106 CFU/mL. To maintain constant OTC or LGG concentration for the period of exposure, the OTC or LGG solution was renewed with freshly prepared solution every day.
Experimental design
Zebrafish larvae (5 dpf) were randomly divided into six groups. Among them, the zebrafish in Ctrl group (normal), OTC group (500 μg/L OTC), and LGG group (106 CFU/mL LGG) were bred for 3 weeks (5–25 dpf), respectively, and were tested on day 26 (Fig. 1A). SCH group were bred with no treatment of OTC or LGG during 5–25 dpf, but were treated with SCH23390 (0.1 mg/L) for 1 h at 26 dpf and was tested on day 26 (Fig. 2A). Ctrl group (normal) and OTC+LGG group [500 μg/L OTC (3 weeks) +106 CFU/mL LGG (3 weeks)] were tested on day 47 (Fig. 1A). The behaviors including locomotion, shoal cohesion, and color preference were examined on days 26 and 47, respectively. Then, the methods were used according to published protocols,35,39 on the 26th or 47th day, the fish were washed five times with sterile water to remove the bacteria loosely attached to the skin. Fish were killed with tricaine (MS-222) at 200 mg/mL, and the brains and intestines from fish in each group were aseptically dissected on ice, and frozen in a microcentrifuge tube (30 brains or intestines per tube) at −80°C for dopamine content analysis or gut microbiota analysis.
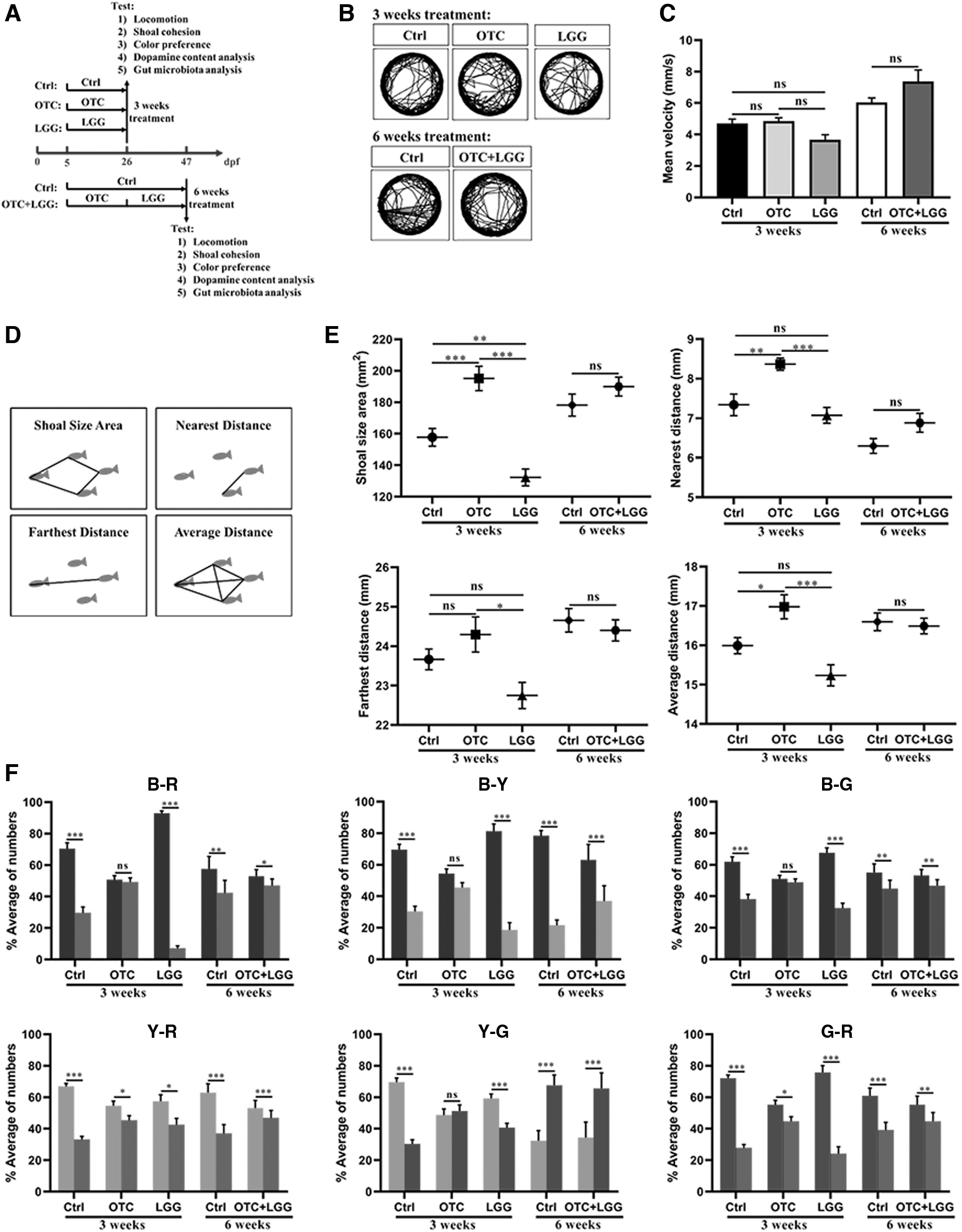
Different treatments affect social interaction and color preference in zebrafish.
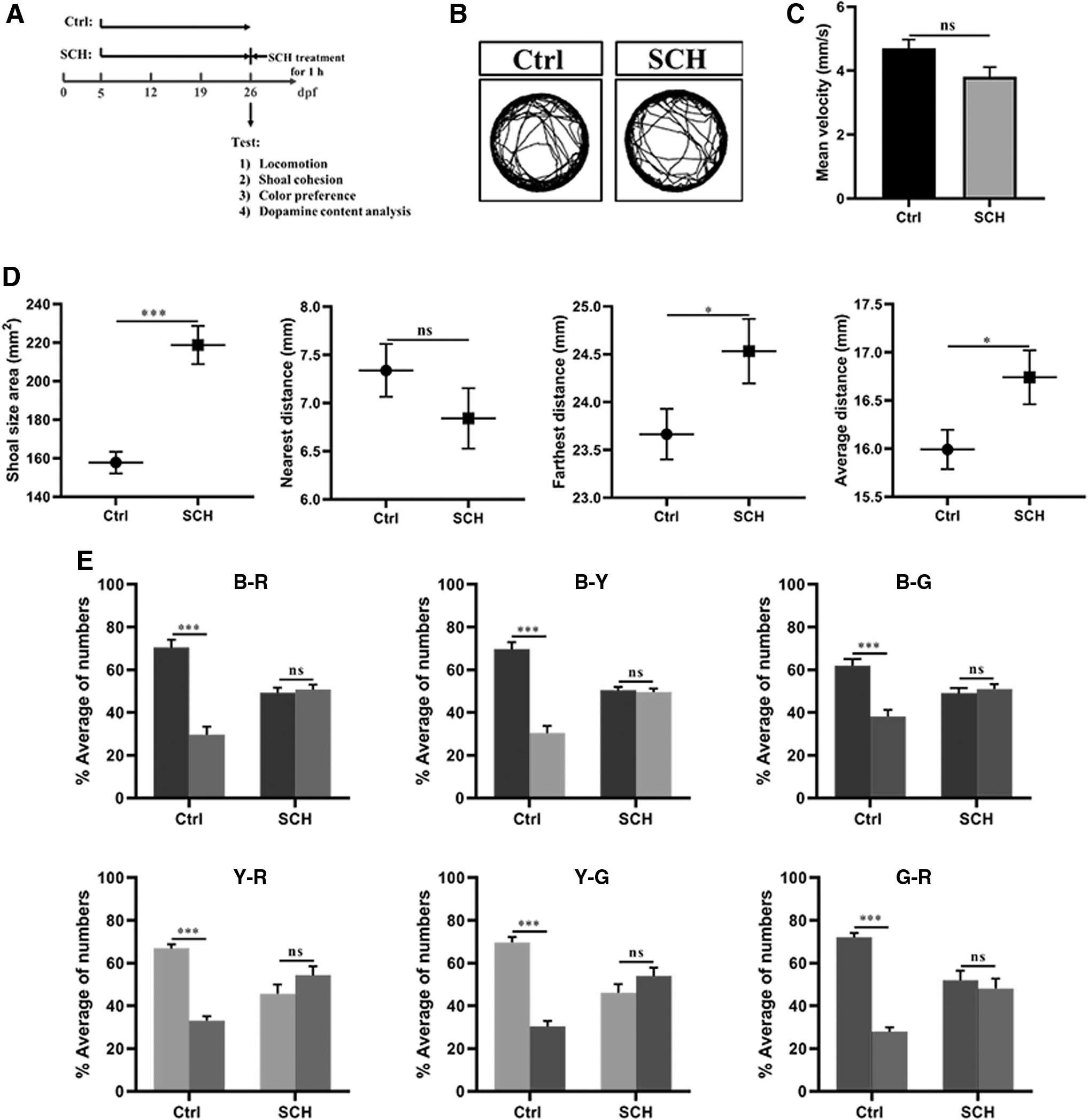
SCH23390 affects shoal interaction and color preference in zebrafish.
Locomotor activity test
Locomotor behavior was assessed according to the published protocol. 40 The fish were transferred into 12-well plates with one fish and 2 mL system water in each well. The individuals were acclimated to the recording condition for 5 min. After the acclimation, locomotor behavior was tested in a 10-min period. The trajectory of the fish was recorded by an industrial camera (Guangdong, China) and the videos were analyzed using LSRtrack and LSRanalyze software (Zhou et al.). 40 The velocity was also recorded to assess motor function.
Shoaling behavioral assay
To test the social behavior, a four-fish assay was used to measure the social interaction of zebrafish in a six-well plate according to Kyzar et al. 41 In brief, 24 normal fish were randomly selected from each treatment and transferred to the six-well plate with 4 fish and 5 mL water in each well. After the acclimation of 5 min, the shoaling behavior was video-recorded for 10 min, and analyzed using 10 screenshots that were made every 1 min during the recording period. Each screenshot was calibrated to the size of the six-well plate and the distances between each fish in the group were measured using ImageJ software. The parameters including shoal size area (SA), nearest distance (ND), farthest distance (FD), and average distance (AD) were calculated to assess the fish shoaling behavior (Fig. 1D).6,42
Color preference test
The color-enriched conditional place preference (CPP) apparatus is a customized fish tank (20 cm length × 10 cm width × 10 cm height), colored with four color combinations [red (R), green (G), yellow (Y) and blue (B)]. To examine the preference between two colors, the CPP tank was divided into two compartments that were covered with the corresponding colors on all sides except the top. A video camera (NVH-589MW; Wang Shi Wu You Corp.) was placed above the CPP tank for vertical video tracking. The color preference apparatus was placed over the LED light panel to ensure the light source could cover the whole tank. The detailed apparatus has been described in our previous work. 43
To test the color preference of zebrafish shoals, the experimental tanks were filled with 500 mL fresh fish water. There was no physical barrier between the two compartments and the 20-larval zebrafish (a shoal) could swim freely in the entire tank. After 5-min adaptation, the proportion of numbers that stayed in each colored zone was recorded every 20 s for 10-min experiment. Larger numbers of fish spent on one color side indicated a preference for this color. The outside of color-enriched CPP tank was opaque to prevent external visual interference from all directions. To minimize the effect of noise, the experimental room was closed and kept quiet, and the experimenter was not visible to the fish during the recording.
Dopamine content analysis
The level of dopamine was measured by enzyme-linked immunosorbent assay (ELISA) according to the manufacturer's (Wuhan Xinqidi Biological Technology Co. Ltd.) instructions. In brief, the precooled PBS (1:9, m/v) was added to the brain tissues of fish, the tissues were ground and centrifuged for 2 min at 5000 g to obtain the supernatant. The standards and the supernatant of each sample were added to each well of 48-well plates precoated with primary antibodies. The plates were incubated at 37°C for 60 min after adding enzyme conjugate, and then rinsed with distilled water. During 30 min of the chromogenic reaction, the optical density (O.D.) was measured at 450 nm by a microtiter plate reader (Thermo). The concentration of dopamine (pg/mg brain tissue) was calculated by comparing the O.D. of the samples to the standard curve.
Bacterial genomic DNA extraction
To analyze the bacterial composition, total bacterial DNA was isolated from 30 intestines per treatment group by using the E.Z.N.A.® Soil DNA Kit (OMEGA). DNA yield was measured by a NanoDrop 2000 Spectrophotometer (Thermo).
Illumina high-throughput sequencing of barcoded 16S rRNA genes
The V3–V4 region of the 16S rRNA gene was amplified with the primers 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′). PCR reactions were performed in a reaction mixture containing (20 μL) 4 μL of 5 × Fast Pfu buffer, 2 μL of 2.5 mM dNTP, 0.8 μL of 5 μM forward primer, 0.8 μL of 5 μM reverse primer, 0.4 μL of FastPfu polymerase, and 10 ng of template DNA. The PCR consisted of an initial denaturation time of 3 min at 95°C followed by 29 cycles (95°C for 30 s, 55°C for 30 s and 72°C for 45 s), and the final extension period lasted for 10 min at 72°C. PCR products were purified and were subjected to Illumina-based high-throughput sequencing (Majorbio Bio-Pharm Technology, Co., Ltd., Shanghai, China). The Vsearch 8.0 software in the QIIME was used to detect and filter chimeric sequence to obtain valid data. Based on the valid data, the sequence data were analyzed using QIIME with an Operational Taxonomic Unit (OTU) definition at a similarity cutoff of 97%.
Bioinformatics analysis
Alpha diversity was calculated by using Phylogenetic Distance over OTU counts, 44 and the beta diversity was estimated by using Unifrac metric. 45 Taxonomic richness and diversity estimators were determined based on OTU abundance matrices in Mothur (version v.1.30.1). Chao was used to assess the community richness, and the diversity was reflected by Shannon. Principal coordinates analysis (PCoA) was the distance matrix and performed a program using MATLAB R2016a environment. One-way analysis of variance (ANOVA) followed by Tukey's test was used for the data analysis of Shannon, Chao, and relative abundance of species level in the 3-week treated groups (i.e., Ctrl, OTC, and LGG). The two-tailed t-test was used for the data analysis of Shannon, Chao, and relative abundance of species level in 6-week treated groups (i.e., Ctrl and OTC+LGG).
Statistical analysis
All experimental results were expressed as means ± standard error of the mean (SEM). Data analysis and graphs were performed using GraphPad Prism version 7.0 (GraphPad Software). One-way ANOVA followed by Tukey's test was used for the data analysis of locomotion, social cohesion, dopamine, and microbiota in the 3-week treated groups (Ctrl, OTC, and LGG). The data analysis of 6-week treated groups (Ctrl and OTC+LGG), SCH group, and color preference of all the six groups was performed by the two-tailed t-test. Significance was set at p < 0.05 for all the experiments.
Results
Analysis of social interaction and color preference in zebrafish through OTC and/or LGG treatment
In this study, to assess the influence of OTC or LGG on shoal interaction and color preference, the larval zebrafish (5 dpf) were exposed to OTC (500 μg/L) and LGG (106 CFU/mL) for 3 weeks, respectively (Fig. 1A). More importantly, the natural microbiota has a small amount of probiotic lactic acid bacteria, beneficial for social interaction. 5 However, it is not clear whether probiotic can improve the deficiency of color preference induced by antibiotic treatment. To address this issue, zebrafish larvae (5 dpf) were treated with OTC (500 μg/L) for 3 weeks, and then probiotic LGG (106 CFU/mL) was applied daily for 3 weeks (Fig. 1A).
To investigate the locomotor behavior of fish that may affect shoaling behavior and color selection, the fish were transferred into 12-well plates to measure locomotion. The behavioral trajectory and the results of average movement speed (i.e., velocity) were not significantly changed after OTC and LGG exposure (Fig. 1B, C). The results showed that OTC or LGG may not cause the locomotor change in zebrafish, and the LGG strain could not change the behavioral trajectory and mean velocity after OTC exposure.
As demonstrated in Figure 1E, after the 3-week treatments, for fish in OTC group, compared with those in the Ctrl group, although FD was not significantly different, SA (p < 0.001), ND (p < 0.01) and AD (p < 0.05) significantly increased. In addition, shoaling behavior of fish was significantly different between the Ctrl and LGG groups. Although ND did not change between the two groups (p > 0.05), FD (p = 0.09) and AD (p = 0.08) were decreased, and SA (p < 0.01) was significantly decreased in the LGG group. Together, these data indicated that the shoal cohesion in fish was decreased by the OTC exposure, and then increased by the LGG exposure. We next set out to identify the social cohesion by LGG treatment after OTC exposure, as given in Figure 1E; after 6 weeks of treatment, shoaling behavior showed no significant difference between the Ctrl and OTC+LGG groups. These data indicated that probiotic LGG could enhance the ability of shoal cohesion that was reduced by the treatment of OTC.
As given in Figure 1F, after the 3-week treatments, in Ctrl group, there was a distinct color preference among two color combination (B-R, B-Y, B-G, Y-R, Y-G, and G-R), and the order of BYGR preference was B > Y > G > R. However, the OTC group lost their color preference (B-R, B-Y, B-G, Y-G) or attenuated their color selection (Y-R, G-R). Unlike the OTC group, the LGG group maintained their color preference except that the preference of Y-R was slightly decreased, and the order of BYGR preference was the same as that of the Ctrl group (B > Y > G > R). Hence, our results showed that the color preference of shoals was lost or weakened by the OTC treatment, while it was retained by the LGG treatment. Next, we investigated the variation of color preference of the fish exposed to OTC and LGG simultaneously (OTC+LGG group). Although the selections of colors among B-R, B-G, Y-R, and G-R were slightly weakened, the order of BYGR preference was the same as that in the Ctrl group (B > G > Y > R). These data suggest that LGG could maintain the existence of the color preference in shoals that was lost after the OTC treatment.
SCH23390 affects shoal interaction and color preference in zebrafish
From the above results in Figure 1E, F, the strength of social cohesion may also affect the color discrimination in shoals of zebrafish. To identify the mechanism of shoaling behavior's effect on color preference, we investigated a D1-receptor antagonist SCH23390, which could disrupt social preference and decrease social interaction in zebrafish. 35 To start the experiment, the SCH23390 (SCH) group (26 dpf) remained in the SCH23390 solution for 1 h (Fig. 2A). It is assumed that since the dopaminergic system is associated with a number of brain functions (e.g., motor function), the D1-receptor antagonist may potentially lead to the altered shoaling behavior as a result of abnormal motor or activity level. However, our results demonstrated that the fish in SCH group did not display abnormal locomotory activity or posture patterns and their mean velocity was also unaltered (Fig. 2B, C).
From the results in Figure 2D, shoal cohesion of fish treated with SCH23390 was significantly different from that in the Ctrl group. The results for SA (p < 0.001), FD (p < 0.05), and AD (p < 0.05) were significantly decreased in the SCH group, but ND did not alter between the two groups (p > 0.05). These data suggested that SCH23390 significantly decreased social behavior in zebrafish. From the results of color preference, in contrast to the shoals without SCH23390 treatment, the shoals treated by SCH23390 lost their interest in color combinations in the color-enriched CPP tank (Fig. 2E). These results suggest that the color preference of shoals required the participation of social interaction.
Dopamine level analysis of the treatment groups
As the D1-receptor antagonist mediated the social interaction and affected color selection in shoals of zebrafish (Fig. 2D, E), we hypothesized that changes in social interaction and color preference of the exposed fish may be related to dopamine level in their brain. To identify the dopamine level, we tested the dopamine concentration of the exposed groups using ELISA. As given in Figure 3A, OTC treatment significantly decreased the dopamine level (p < 0.05), whereas LGG treatment significantly increased the dopamine concentration (p < 0.05). Meanwhile, LGG treatment enhanced the dopamine level after the OTC treatment (p > 0.05). In addition, our data verified the results that SCH23390 can significantly decrease the dopamine level (p < 0.001) in the brains of fish (Fig. 3B). These results suggest that dopamine, which was associated with social cohesion, contributed to the color preference of the fish.

Dopamine level analysis of the treatment groups.
Gut microbial communities of zebrafish in OTC or LGG treatment group
Considering the close relationship between antibiotic/probiotic and intestine, intestinal health is very important to fish. The gut microorganisms of the OTC and LGG groups were identified using 16S rRNA gene sequencing of the total DNA extracted directly from the two groups. According to a Venn chart at the OTU level for the similarity of each group with 152 shared members, the OTC group showed low similarity compared with that in the Ctrl group (Fig. 4A). The community diversity (Shannon) and richness (Chao) were evaluated as given in Figure 4B and C. Compared with that of the Ctrl group, the OTC group had lower bacterial diversity (p = 0.06), whereas the LGG group had higher bacterial diversity (p < 0.01) (Fig. 4B). The microbial richness from the OTC group had a significant decrease compared with that of the Ctrl group (p < 0.05), whereas the LGG group showed an increasing trend but not statistically significant (p > 0.05). PCoA was used to characterize the distribution of variations in community composition among the three groups, with most of the variations explained by the first two coordinates (22.5% and 36.7%, respectively; Fig. 4D). Samples from the OTC and the LGG groups were distinctly separated from the Ctrl group, indicating that the OTC or LGG treatment had a great influence on gut bacterial communities.
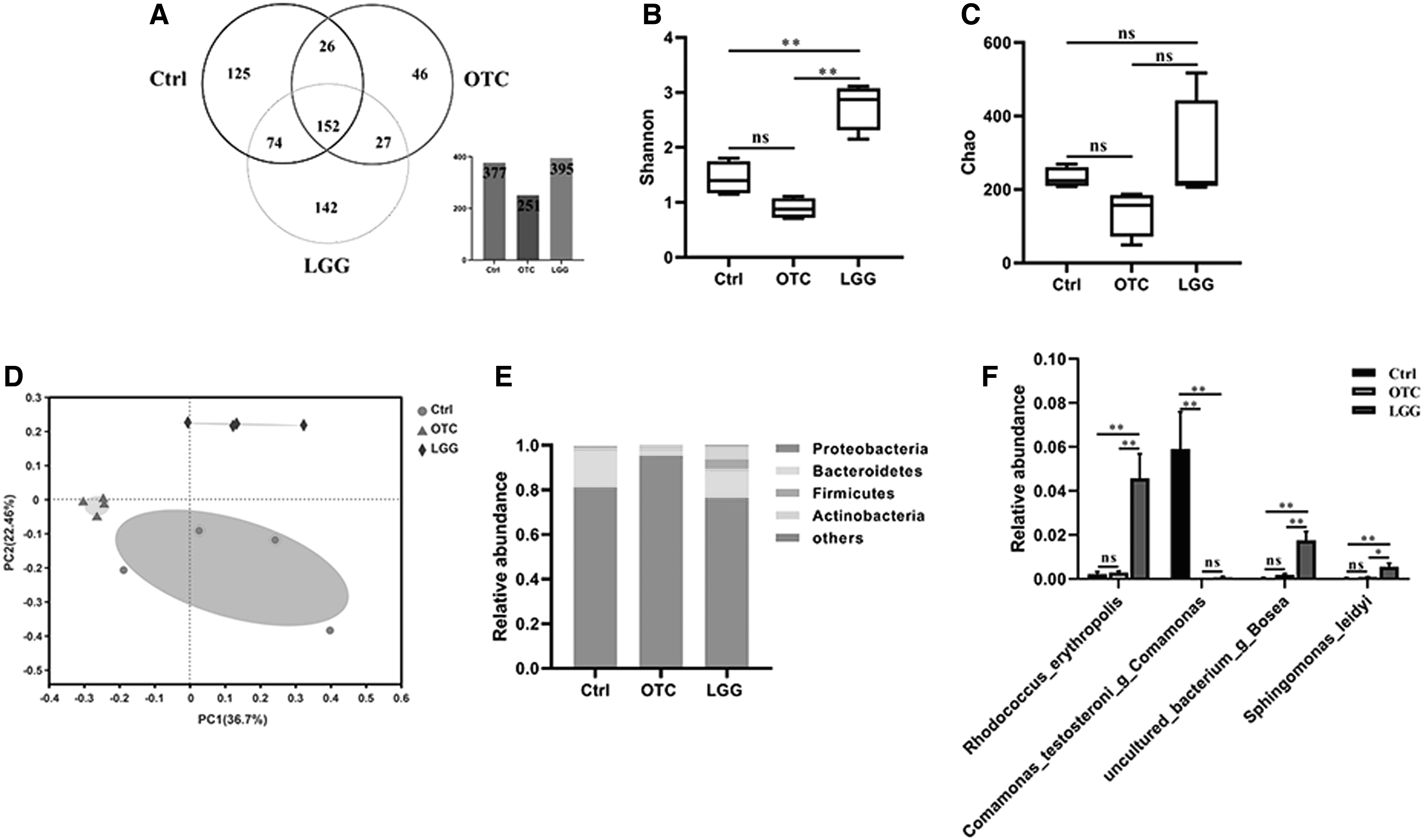
Effect of OTC or LGG exposure on the intestinal microbial composition.
The sequencing results of the gut microbiota revealed that Proteobacteria was the predominant phylum. In the Ctrl group, the relative abundances of the four phyla (Proteobacteria, Bacteroidetes, Firmicutes, and Actinobacteria) were 81.0%, 17.7%, 0.8%, and 0.4%, respectively. In the OTC group, the relative abundances of the four phyla were 95.0%, 3.8%, 0.3%, and 0.9%, respectively. In the LGG group, the relative abundances of the four phyla were 76%, 6.4%, 6.6%, and 9.8%, respectively. To assess the relative abundance of species level, the relative abundances of the four species are given in Figure 4F. Rhodococcus_erythropolis was the most abundant bacterium in the LGG group and had a significant increase compared with the Ctrl group. Comamonas_testosteroni_g_Comamonas showed a significant decrease in the OTC or LGG group compared with that in the Ctrl group. A significant increase of relative abundance in uncultured_bacterium_g_Bosea was observed in the LGG group. In addition, Sphingomonas_leidyi also showed a significant increase in the LGG group. Hence, these data indicate that the OTC or LGG treatment could affect the intestinal bacterial communities.
Gut microbial communities of zebrafish in OTC and LGG treatment group
We also analyzed the composition of zebrafish intestinal microbiota by LGG treatment after OTC exposure. As given in Figure 5A, the similarity between the Ctrl group and the OTC+LGG group were 162 shared members in a Venn diagram at the OTU level. From Figure 5B, C, the microbial richness (Chao) and community diversity (Shannon) from the OTC+LGG group showed a decreasing trend with the Ctrl group but not statistically significant (p > 0.05). Samples from the Ctrl and OTC+LGG groups clustered together in PCoA (Fig. 5D), indicating that the OTC+LGG group had a lower influence on gut bacterial communities.
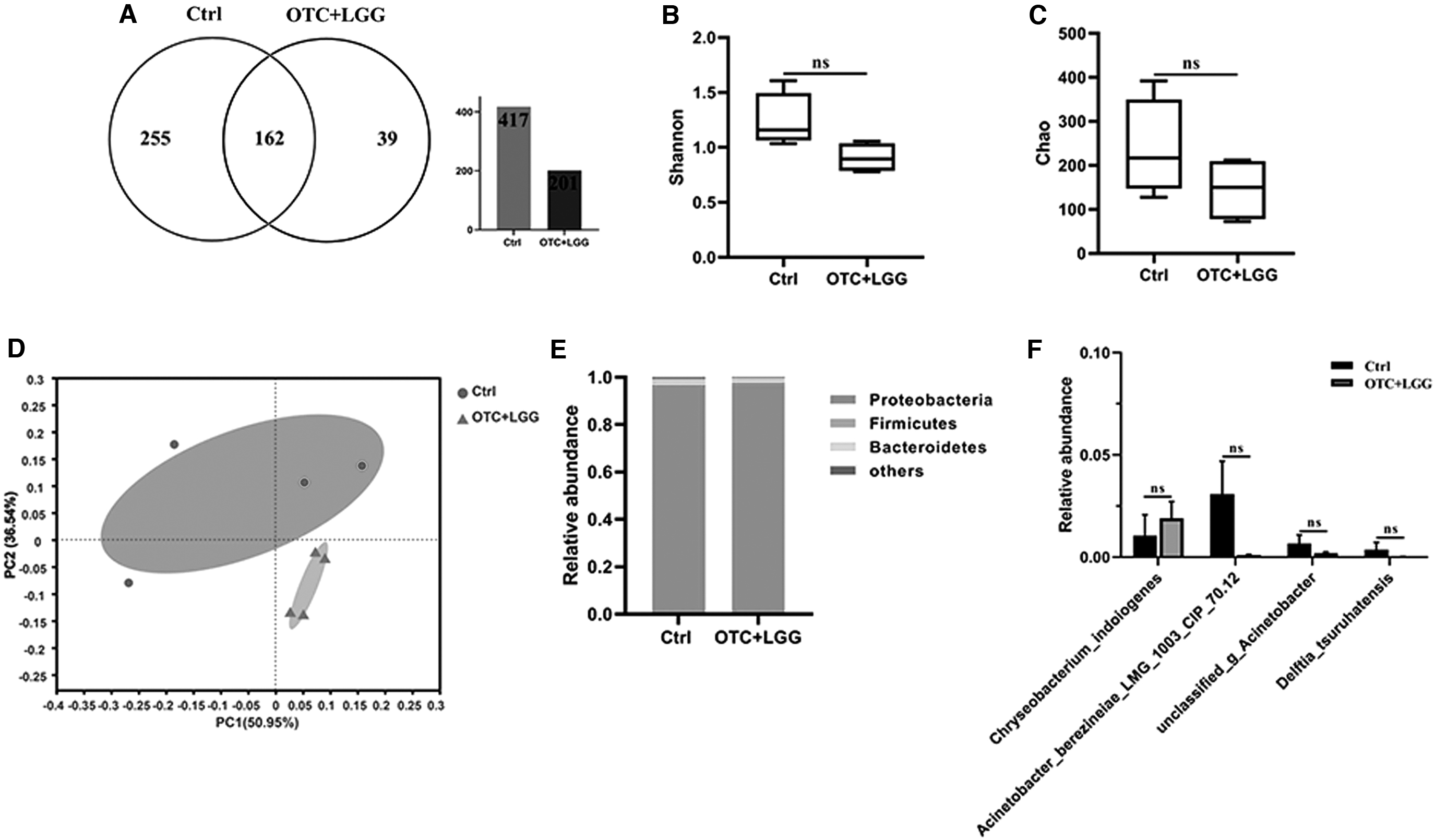
Effect of OTC and LGG exposure on the intestinal microbial composition.
The relative abundances of gut microbiota between the Ctrl group and the OTC+LGG group at phylum level are given in Figure 5E. The dominant phyla in the two groups were Proteobacteria, Bacteroidetes, and Firmicutes. In the Ctrl group, the relative abundances of the three phyla were 96.5%, 2.2%, and 2.3%, respectively, whereas the relative abundances of the three phyla in the OTC+LGG group were 97.4%, 2.1%, and <0.1%, respectively. In addition, the relative abundances of four species in the Ctrl and OTC+LGG groups were also assessed as given in Figure 5F. The results showed that the relative abundances of four species did not have a significant difference between the two groups. Therefore, these results indicate that the enhanced social cohesion and maintained color preference of zebrafish by the probiotic LGG after OTC exposure may be through its influence on intestinal microbiota.
Discussion
The shoaling behavior is one of the most robust and consistent behavioral features of zebrafish because it has been observed both in nature 46 and in the laboratory. 47 Fish often form aggregation in lakes, puddles, ponds, rice fields, ditches and small watercourses. 48 Social cohesion varies remarkably under different living conditions. As a commonly used antibiotic for human and other animals, 49 OTC has been frequently detected in surface water, groundwater, and seawater. 50 Probiotic as a living and supplementary microorganism is found in foods (e.g., yogurt, snacks, breakfast cereals, and infant formulas). 51
From our results, the social cohesion of zebrafish was decreased by the OTC treatment, and increased by the LGG treatment (Fig. 1E). More importantly, the probiotic LGG could enhance the social cohesion after the OTC treatment (Fig. 1E). Our data of the decreased shoaling behavior were consistent with the study by the antibiotic exposure. 4 Moreover, many studies on probiotics (such as L. reuteri, L. rhamnosus IMC 501) have shown their potential role in regulating social behavior.5,6 Therefore, it is reasonable that changes in living conditions caused by OTC or LGG may affect sociability and social cognition of zebrafish, and probiotic LGG supplementary could increase the social cohesion that is reduced after the OTC exposure.
The intricate living conditions usually contain abundant colors, and the color preference would spur shoal on to emigrate to the environment with a favorable color background that is beneficial for social contact. To investigate color preference of zebrafish shoals in different exposure conditions, we measured the color selection of fish shoals with different treatments. According to Figure 1F, the fish in the Ctrl group show a clear and strong color preference in shoals, and the order of RYGB preference was B > Y > G > R. In addition, we found that the OTC group lost or weakened their color preference. By contrast, the LGG group maintained their color preference in shoals. To our surprise, probiotic LGG supplementation could maintain color selection after the color deficiency caused by OTC (Fig. 1F). Although color selections among some color combinations were slightly weakened, the order of BYGR preference was the same with that in the Ctrl group (B > G > Y > R).
Some results of color preference were consistent with the findings reported by Park et al. 25 and Peeters et al. 52 They found that zebrafish preferred B over R, and B over Y, and B over G. In this work, the fish by different treatments did not exhibit abnormal locomotor behavior or behavioral trajectory (Fig. 1B, C). A possible explanation for contradictory conclusions in existing literatures may be that the 5 dpf larvae have not developed shoaling behavior yet 26,27 or the microbiota changes affect behavior of color preference. Besides, from our results, the order of color preference for Y-G combination was different between the two Ctrl groups at different developmental days. Color is perceived primarily through cones in the retina of zebrafish, and fish do not possess yellow cones for sensing yellow light. 53 Therefore, we hypothesized that the deficits of yellow cones may cause the observed difference during color selection.
From our results in Figure 2, the D1-R antagonist disrupted the social interaction, leading to the deficits of color preference in shoals. The fish treated by SCH23390 did not exhibit abnormal motor or posture (Fig. 2B, C), and the results of locomotor behavior were consistent with the study by Scerbina et al. 35 The dopaminergic system is involved in several brain functions, which has been found to be associated with the shoaling tendencies. 54 Dopamine receptors distribute in different brain regions. Clearly, the specific areas are involved in cognition, including hippocampus, the prefrontal cortex, the amygdale, and the ventral and dorsal parts of the striatum. There are four different dopamine receptor subtypes (D1, D2, D3, and D4) in the brain of zebrafish.38,55,56 Among them, the excitatory D1 receptor (D1-R) subtype is the most predominately expressed in the brain regions. 57 D1-R activates the production of intracellular 3′-5′-cyclic adenosine monophosphate through adenylyl cyclase induction and regulates intercellular calcium signaling or protein kinase activity. 58 Therefore, these results suggest that social interaction can be affected by dopaminergic system that could be responsible for eliciting the preference for colors.
We next identified the dopamine levels of different treatments that showed clear effect on social cohesion and color discrimination of zebrafish. From our results it is shown that the impact of OTC or LGG on dopamine level contributes to the changes in social cohesion and color preference. The decrease in dopamine levels implies the reduced dopamine production and/or increased dopamine degradation in response to the applied SCH23390. 59 Yung et al. 60 have shown that D1-R localized in the postsynaptic terminals of neurons in the basal ganglia. The blockade of postsynaptic neurotransmitter receptors may impair signaling downstream and reduce neurotransmitter release.
The antagonist of D1 receptor could increase the concentration of dopamine in the synaptic cleft, leading to reuptake and leakage to extra-synaptic areas. The increased extra-synaptic dopamine could activate dopaminergic autoreceptors on the presynaptic neuron and inhibit the dopamine synthesis. 61 To sum up, these studies and our work have shown that dopamine levels have an impact on social interaction and color preference in shoals, and the lack of sociability and color discrimination is related to the decreased dopamine level in the brain of zebrafish.
The altered microbiota in the gastrointestinal system may influence neurodevelopment and programming of social behaviors. 1 Indeed, the antibiotic OTC treatment was associated to the reduction in gut microbiota diversity (Shannon) and richness (Chao) (Fig. 4B, C). Moreover, under condition of OTC exposure, the relative abundance of microbiota on phylum level or in species level was influenced (Fig. 4E, F). By contrast, probiotic LGG could promote gut microbiota diversity and richness (Fig. 4B, C), and the relative abundance of microbiota on phylum level (Bacteroidetes, Firmicutes, Actinobacteria) or on species level was also increased by the LGG treatment (Fig. 4E, F). Although antibiotic administration often perturbs microbiota, modulation of gut bacteria through probiotic strain LGG supplementation can have a beneficial result (Fig. 5).
Proteobacteria including potentially pathogenic bacteria was the dominant member of the gut microbiota. The relative abundance of Proteobacteria was less represented in the LGG group (76%) than that in the Ctrl (81%) and in the OTC group (95%), although the difference was not statistically significant. A higher proportion of Proteobacteria that is correlated with enterocolitis severity characterized the gut microbiota dysbiosis of zebrafish. 62 Although LGG was not detected in species level, the relative abundance of Firmicutes phylum (6.6%) increased in the LGG group. Our results were consistent with previous studies reporting increased abundance of Firmicutes in zebrafish after probiotic treatment. 6 The higher abundance of Firmicutes phylum might be related to the LGG administration and LGG might be symbiotic relationship between three species levels, which had a higher abundance in the LGG group.
The intestinal microbiota signaling to the brain may affect behavioral processes of sociability through numerous pathways, including activation of the vagus nerve, the production of microbiota metabolites (including neurotransmitters), and sensory pathways (such as olfactory system). 63 In the route of vagus nerve, the vagus nerve fibers are enriched with receptors (including 5-HT3, Toll-like receptor 4, free fatty acid receptors, and gut peptide receptors), which may transmit signals from gut lumen to the brain. 64 For example, the L. rhamnosus JB-1 lost its ability to modulate anxiety-like behavior and γ-aminobutyric acid-mediated neurotransmission in mice through vagotomy. 65 Similarly, the anxiolytic-like effects of Bifidobacterium longum NCC3001 were absent in mice that underwent vagotomy. 66
In addition, Gut microbiota produced vast metabolites through multiple pathways of microbial metabolism, such as volatile carboxylic acids, esters, neurotransmitters (e.g., dopamine), and fatty acids, some of which may affect brain physiology and behavior. 67 In addition, in vitro studies have indicated that certain bacteria have the capacity to produce neurotransmitters such as noradrenaline, dopamine, and γ-aminobutyric acid.68,69 Some researches have shown that the gut microbiota can influence serotonergic neurotransmission by regulating the availability of its precursor, tryptophan. 70 Thus, these data and studies suggest the possibility that microbiota play critical roles in the social behaviors and brain functions through the microbiota–gut–brain axis.
So far, numerous discoveries linking gut microbiota to behavior have been obtained from studies on rodent animals.5,7,8 Compelling and accumulating evidences reveal that the presence and activity of gut microbiota strongly affect the behavior and host physiology.6,71 However, the mechanism of the relationship between shoals' behaviors and intestinal microbiota still remains controversial and the roles of responsible neurotransmitters are unclear. Taken together, our data reveal that specific social behavioral, neurochemical, and gut microbiota changes are induced by antibiotic OTC/probiotic LGG treatment, and the correlation between OTC and/or LGG and behaviors through neurotransmitter (dopamine) of brain has been established. In addition, the potential evidences for the relationship between gut microbiota and dopamine level of brain through microbiota–gut–brain axis may also be found.
Despite the above findings, further studies are necessary to connect these domains in a causal manner. Another limitation of this study, as in most other studies investigating microbiota–gut–brain relation, is that the mechanism of the relationship between gut microbiota and behaviors is incomplete. A deeper understanding of the mechanism will be crucially important for development of any probiotic-based therapeutic strategy for psychiatric diseases in human and animals.
Footnotes
Disclosure Statement
The authors have no conflicts of interest to declare.
Funding Information
This study is supported by the National Key Research and Development Plan of China (2017YFC1600603), and Funds for Huangshan Professorship of Hefei University of Technology (407–037019).