
Abstract
The rapid increase in zebrafish use needs to be accompanied by research into the refinement of procedures. The European (EU) Directive lists three possible euthanasia methods for fish: anesthetic overdose, electrical stunning, and concussion. However, for small fish such as zebrafish, concussion and electrical stunning are difficult to perform, leaving anesthetic overdose as the most used method. Our aim was to test the efficacy and side effects of anesthesia overdose using different anesthetics and the rapid cooling method to euthanize adult zebrafish. Adult mixed-sex AB zebrafish were randomly assigned to: 250 mg/L MS222; 20 mg/L propofol +100 mg/L lidocaine; 6 mg/L etomidate; 50 mg/L clove oil; and rapid cooling (water at 2°C–4°C). Two minutes after opercular movement ceased, animals were transferred into clean water for 20 min and recovery assessed, or decapitated and used for biochemical analysis of the gills, muscle, liver, and brain; for the histological analysis of the gills and muscle; or for the assessment of cortisol levels. No animal recovered; rapid cooling was the quickest and etomidate overdose was the slowest method to cease the opercular movements. There were no major differences between euthanasia methods regarding the biochemical or histological data. Cortisol levels were higher in the rapid cooling group, but only when compared with the propofol/lidocaine group. The use of a physical method of euthanasia, such as rapid cooling, is essential when chemicals, such as anesthetics, may interfere with postmortem analyses. Although anesthetic overdose can be used without major effects on the analyses conducted in this work, rapid cooling can be another option with the advantage of being simple to administer, easily available, affordable, and very quick; this decreases the potential duration of suffering, being more humane. Therefore, a change in EU legislation should be considered to include additional humane options for euthanasia, such as rapid cooling, for zebrafish and other small tropical fish.
Introduction
According to the latest statistics from the European (EU) Commission regarding the use of animals for scientific purposes, 1 fish are the second most used group of animals, with one single species, zebrafish (Danio rerio), representing half of this use. Thus, it is of utmost importance to refine routine procedures in this context, such as euthanasia/humane killing. Euthanasia is necessary in many situations: terminal tissue collection for research; when the levels of pain, distress, and suffering are likely to exceed the designated level; when animals have an incurable disease; or when animals are no longer serving the purpose for which they were reared. 2
A humane method of euthanasia should fulfill several criteria: it should cause minimal distress and pain for the animal, afford rapid unconsciousness and death with minimum restraint, be irreversible, and provide a reliable central nervous system (CNS) depression. In addition, it should be reproducible, simple, and practical to perform; safe to the operator; aesthetic (not unpleasant to the operator's perception, thus avoiding an emotional burden and a poorly performed technique); economically viable; adapted to animal species, strain, age, number, and health; and, when applicable, in the case of research, be used without compromising postmortem analyses.2,3 Finally, it should consider the public's perception of the method.
The EU Directive 2010/63 lists three possible euthanasia methods for conscious fish: anesthetic overdose, electrical stunning, and concussion. Regarding the physical methods, the killing of a small fish, such as zebrafish, via concussion requires a skilled operator and does not allow the collection of an intact brain, whereas electrical stunning requires special equipment, which is still being optimized for zebrafish to avoid the spasms caused by the electrical current, resulting in hemorrhage. 4 Both methods only allow the killing of one fish at a time. Thus, the most used euthanasia method in adult zebrafish is anesthetic overdose in a water bath.
Tricaine methanesulfonate (MS222) is the most commonly used anesthetic in this setting. This agent blocks sodium ion channels, leading to unconsciousness, immobility, and ultimately vascular collapse and death caused by hypoxia. 3 A previous work has shown that fish display signs of distress in the presence of MS222, regardless of whether it is buffered or not. 5 Over the years, different alternative anesthetics have been suggested for fish euthanasia.6,7 Clove oil is highly lipophilic and is absorbed through the skin and gills rapidly, thus entering the bloodstream. Its action on the brain induces CNS depression, death by respiratory inhibition, and, ultimately, cardiac arrest. 8 This agent is widely available and has a low cost and wide range of efficacy. 9 Etomidate is a nonbarbiturate hypnotic agent that acts by depressing the CNS. It does not provide analgesia, but it is a relatively quick-acting anesthetic. 2
In turn, propofol/lidocaine is a new anesthetic protocol that has been proposed by our group with the potential to be used in zebrafish. 10 Propofol is a short-acting, rapidly metabolized hypnotic, 11 which when overdosed causes respiratory depression in mammals and fish. 12 Lidocaine acts as a local anesthetic by binding to and blocking sodium channels, thus preventing the self-propagating depolarization signal to travel along the length of the axon. 13 Similar to that observed for MS222, lidocaine leads to unconsciousness and eventually death in zebrafish via this mechanism of action. 3 Moreover, it is relatively cheap and easy to obtain. 14
The lack of an easily available physical method of euthanasia for zebrafish that avoids the pharmacological effects on the experimental output has led researchers to propose the use of rapid cooling, as discussed in a EuFishBiomed workshop (now the European Zebrafish Society), 15 and in the study by Valentim et al. 4
Rapid cooling (or hypothermic shock or rapid chilling) is a method that is accepted in the United States for euthanasia in adult zebrafish, according to the newest guidelines of the American Veterinary Medical Association (AVMA). 16 However, it cannot be used on conscious fish (in general) under the present EU legislation, although some studies revealed it to cause less distress to zebrafish compared with MS2225,9,17 and to have the advantages of affording tissues that are free of chemicals for postmortem analysis and posing minimal risk to the operator compared with the use of chemical or other physical methods. 5 Nevertheless, concerns remain about whether this method induces unconsciousness, with the signs varying according to the species. For instance, catfish and eel18,19 showed low brain activity and loss of response to pain, whereas turbot 20 exhibited a decrease in total power magnitude, but still responded to pain stimulus, with no shift in the electroencephalogram to lower frequencies, thus probably not achieving unconsciousness. These data are still lacking for adult zebrafish.
The aim of this study was to clarify the efficacy and assess the putative side effects of different euthanasia protocols (anesthetic overdose using different agents and rapid cooling) in adult zebrafish, to identify specific methods that are adequate to this small tropical fish.
Materials and Methods
Ethics statement
All procedures were carried out under personal and project licenses (014703/2017-06-16) approved by the National Competent Authority for animal research (Direção-Geral de Alimentação e Veterinária, Lisbon, Portugal) and by the Institutional Animal Care and Use Committee (IACUC) of IBMC (2016-27) for a project in which this study protocol was described. All experimental procedures were performed in accordance with the European Directive 2010/63/EU on the protection of animals used for scientific purposes, and its transposition to the Portuguese law (Decreto Lei’ 113/2013).
Animals and housing
For the pilot and main study, 215 adult mixed-sex AB zebrafish (D. rerio) (8–12 months old) bred in the Animal Facility of the institute (i3S, Porto, Portugal) were used between 2 p.m. and 6 p.m.; for the cortisol analysis, this period was shorter (2–4 p.m.). In the annual health screening of the zebrafish facility, Pseudoloma neurophilia was detected (subclinical disease), but no pathogen was identified based on the histology of the fish used in this work. The fish were not subjected to previous experiments.
They were maintained in groups inside 3.5-L tanks (maximum of 5 fish per liter), placed in a recirculating water system, which was connected to the reverse osmosis system and to a central unit of water purification (Multi-linking WTU; Tecniplast, Buguggiate VA, Italy). Thus, water parameters were controlled (27°C ± 0.5°C, pH of 7 ± 0.5, conductivity of 700–715 μS), and the facility was maintained in a 14 h:10 h light:dark cycle. Adult fish were fed three times a day with a commercial diet (GEMMA micro 300; Skretting, ME) supplemented with artemia (after 21 days postfertilization).
Euthanasia methods: pilot study
Three adult zebrafish per concentration of anesthetic were used to test which concentration of each anesthetic protocol led to death, to choose the lowest effective dose for irreversible euthanasia for the main study. The three animals that had received the concentrations chosen for the main study were included in the final analysis, to keep the number of animals used in the main experiment to a minimum. Choosing the lowest effective concentration of anesthetic allows the use of a lower amount of the agent, thus being more economical and with less chemical residuals being retained in the biological samples. The detailed procedure used in this pilot study was that described below for the main study.
Euthanasia methods: main study
Mixed-sex zebrafish (53% male:47% female ratio) were randomly assigned to five different groups using the Rand () function of Excel (n = 12): one group was euthanized with 250 mg/L of MS222 (MS222 group), one group was euthanized with 20 mg/L of propofol combined with 100 mg/L of lidocaine (P/L group), one group was euthanized with 6 mg/L of etomidate (Et group), one group was euthanized with 50 mg/L of clove oil (CO group), and one group was euthanized with cold water (between 2°C and 4°C) (RC group).
The order of group testing was also randomized. The buffered MS222 solution was prepared by adding MS222 powder (E10521; Sigma–Aldrich) to system water, to prepare a stock solution of 10 g/L buffered with sodium bicarbonate until pH 7.0 was reached. Clove oil was prepared in a 9:1 ratio of ethanol to clove oil solution (C8392; Sigma–Aldrich). Propofol (Lipuro 2%, B. Braun, Barcarena, Portugal), lidocaine (2%; B. Braun), and etomidate (Lipuro 2 mg/mL; B. Braun ) did not require any previous preparation.
An anesthetic water bath of 200 mL was always freshly prepared using system water inside a 1-L tank, that is, each solution was always prepared before the beginning of the experiment. For the rapid cooling method, ice was added to a 1-L tank filled with system water until the water temperature reached 2°C–4°C. An internal tank with a perforated bottom (ZB10BTI; Tecniplast, Ultragene, Santa Comba Dão, Portugal) was then placed on top of the ice to avoid contact between the animals and the ice. Zebrafish were individually placed in the prepared water bath (with anesthetics or cold water), and the time to the loss of equilibrium was recorded when the fish stayed for more than 3 s in lateral or dorsal recumbency.
After loss of equilibrium was achieved, the caudal fin was gently pinched with forceps until the animal had no response to this painful stimulus. The response to the painful stimulus was evaluated every 30 s. After the loss of the painful stimulus, the opercular movement was monitored. After 2 min without opercular movement, cessation of the opercular movements was recorded. Thus, the latency for this movement to stop was considered to have occurred 2 min before the recorded time. When no opercular movements were detected for 2 min, the tested individuals were placed into another tank with clean water (nonmedicated water from the water system at ∼27°C) for 20 min.
If no signs of recovery were noticed, the animals were deemed to be dead. Thus, the total time of exposure to the anesthetic solutions or to cold water was the latency to the opercular movements to cease plus 2 min. The water bath (with the anesthetic or cold water) was not re-used, and a new solution/bath was prepared for each zebrafish that was euthanized individually. Thus, the animal was the experimental unit, with the exception of the biochemical analyses, as the biological material of one animal was not sufficient to perform these analyses, in which case three animals/organs per sample were pooled.
Side effect of euthanasia methods
The most promising protocols were chosen for further analysis in a cortisol assay, biochemical analysis, and histological assessment. Only etomidate was not tested in these analyses because of the long time it required to euthanize an animal using this agent. The following analyses were performed to check if any of the methods of euthanasia mentioned above would interfere with these parameters, leading to nonreliable research outcomes. Moreover, cortisol levels and oxidative stress parameters were tested to evaluate the physiological stress and the general welfare and health of the animals.
The biochemical parameters evaluated here are often used in zebrafish research and can be altered by anesthetics.21,22 Parameters related to metabolism and reactive oxygen species (ROS) are particularly important to study the hypoxia that can be induced by high anesthetic concentrations 23 and cause stress, thus compromising fish welfare. The level of acetylcholinesterase (AChE) in the brain can also be an indicator of oxidative stress. 24 Moreover, anesthetics interfere with neurotransmission, and AChE activity was measured as an indicator of this process. 25 In addition, AChE is involved in muscle contraction. To check if it was altered by the rapid cooling, during which fast muscle contractions (bending) were observed, AChE activity was also assessed in the muscle. 26
Histological changes were evaluated in the gills, which are the organs of anesthetic uptake and are in close contact with these chemicals, as well as in the muscle, to confirm if the bending observed in some animals could lead to muscle fiber disruption.
Cortisol assay
A different batch of zebrafish was euthanized using the protocols described for the main study (with the exception of etomidate overdose) (n = 12). Seven females and five males were included in the RC, MS222, and P/L groups, whereas the CO group comprised six animals from each sex. At 2 min after the last opercular movement, fish were decapitated, their sex was identified, and the body was collected to measure cortisol levels as described previously, 27 with minor modifications. Eggs were removed from abdomen of females, and the bodies of the fish were weighed. The body of each fish was placed in a 15-mL centrifuge tube at −20°C containing phosphate-buffered saline (PBS, pH 7.4) for later analysis.
Briefly, the samples were thawed, cut into small pieces, and transferred to a 2-mL centrifuge tube before being homogenized twice using a TissueLyser II equipment (Qiagen, Hilden, Germany), at 30 Hz for 90 s. After a brief centrifugation (3 min at 7000 rpm, 4657 g, Biofuge Pico centrifuge; Heraeus Instruments, Germany), the aqueous layer was transferred to a 10-mL centrifuge tube and 5 mL of diethyl ether were added to extract cortisol. After vortexing, the samples were centrifuged (10 min at 3500 rpm, 1164 g), the organic layer was transferred to new tubes, and the extraction procedure was repeated once.
The organic layers were collected in a single tube and allowed to dry at room temperature under a fume hood until a yellow oil containing cortisol remained. Five hundred microliters of PBS was added to the tubes, and cortisol levels were assayed using a cortisol enzyme-linked immunosorbent assay kit (Salimetrics assay #1-3002; Salimetrics, State College, PA) as per the manufacturer's instructions, using spectrophotometry (Power Wave XS2 plate reader; BioTek) at 450 nm. Cortisol concentrations are expressed as nanograms per gram of body weight after normalization for body weight.
Biochemical analyses
At 2 min after the last opercular movement, fish were decapitated and the liver, brain, gills, and muscle dissected in cold PBS; three fish/organs were pooled as one sample (n = 5), homogenized, processed, and analyzed as earlier described previously. 28 The overall generation of ROS in each organ was assessed using DCFH-DA (2′,7′-dichlorofluorescein diacetate) at 485 nm (excitation) and 530 nm (emission). To assess the antioxidant response, the activities of Cu/Zn-superoxide dismutase (SOD) and catalase (CAT) were evaluated based on the photochemical reduction of nitroblue tetrazolium at 560 nm and the decomposition of H2O2 at 240 nm, 29 respectively.
Standard curves that were prepared using SOD from bovine erythrocytes (0–15 U/mL) and bovine CAT (0–15 U/mL) were employed to calculate specific activities. A method based on the disappearance of NADPH at 340 nm using the extinction coefficient of 6.22 mM−1 cm−1 was used to calculate the activity of glutathione peroxidase. The activity of glutathione-S-transferase was estimated based on the formation of the glutathione–dinitrobenzene conjugate at 340 nm and using the extinction coefficient of 9.60 mM−1 cm−1. Lactate dehydrogenase was assayed at 340 nm based on the oxidation of NADH to NAD+ and using the extinction coefficient of 6.22 mM−1 cm−1.
For the determination of AChE activity, a method based on Ellman's reagent (405 nm) was applied using the 5-thio-2-nitrobenzoic acid extinction coefficient of 13.6 mM−1 cm−1. The inorganic phosphate (Pi, 0–20 mM) released by ATP hydrolysis at 660 nm 30 was used to assess the Na+K+-ATPase activity. The reduced (GSH) and oxidized (GSSG) glutathione forms were derivatized with ortho-phthalaldehyde and measured at excitation and emission wavelengths of 320 and 420 nm, respectively. Concentrations were estimated based on GSH and GSSG standard curves (0–125 μM), which allowed the calculation of the glutathione redox ratio (oxidative stress index).
For the assessment of lipid peroxidation, malondialdehyde (MDA) concentrations were measured via the spectrophotometric quantification of the MDA-TBA adducts at 530 nm and based on a standard curve of MDA (0–30 μM). A DNPH (2,4-dinitrophenylhydrazine)-based photometric assay at 450 nm 31 was used for the quantification of carbonyls based on an extinction coefficient of 22 mM−1 cm−1 Activities/levels were normalized to protein concentration via absorbance measurements performed at 280 nm in a BioTek Take3 microvolume plate (Bio-Tek Instruments). All samples were tested in duplicate and measured against a reagent blank in the corresponding microplate. Enzymatic assays were carried out with 10 μL of the sample at 30°C using a PowerWave XS2 microplate scanning spectrophotometer (Bio-Tek Instruments) or a Varian Cary Eclipse (Varian) spectrofluorometer equipped with a microplate reader.
Histology
At 2 min after the last opercular movement, 10 animals per group were euthanized using the methods described previously; the head and the body of the fish were then separately fixed in buffered formaldehyde (3.7%–4%; Panreac, Spain) for 24 h. Subsequently, the samples were dehydrated in a graded ethanol series, embedded in paraffin, and sectioned (at a thickness of 3 μm) in a microtome (Leica RM 2135; Leica Biosystems, Germany). The head was laterally oriented to obtain longitudinal sections of the gill filaments, although several transversal cuts were also observed. The body was sagittally sectioned for skeletal muscle evaluation. The sections were then stained with hematoxylin–eosin and observed under a light microscope.
On average, 30 gill filaments were analyzed per animal. A qualitative evaluation was performed, and the histopathological changes were recorded in each individual. The prevalence index of each histopathological change was defined as the percentage of fish per treatment group that exhibited that specific change. 32 Histological changes were evaluated according to the semiquantitative methodology described in the study by Monteiro et al. 32 The extent of each histopathological change was assessed using the severity gradation scale (SGS) presented in Table 1 and was defined as the percentage of filaments with a histopathological change in each fish sampled.
Definition of the Severity Gradation Scale to Assess Histopathological Damage
As some animals made abrupt movements with pronounced bending during the euthanasia procedure, the trunk muscle obtained was used to evaluate the possible rupture of the skeletal muscle fibers. Although some bending was observed in other groups, the bending was more pronounced and more frequent in the animals that were subjected to rapid cooling.
Microscopical sections were observed using an inverted microscope (IX 51; Olympus, Japan) equipped with a CCD camera (Color View III; Olympus, Hamburg, Germany). The images were analyzed and photographed using the Olympus CellˆA 2.6 software.
The organ collection and data analyses (clinical and postmortem analyses) were performed by blinded experimenters. However, the experimenter could not be blind to the method of euthanasia during the monitoring of death because of the necessary visual observation of vital signs and the cessation of opercula movements during euthanasia.
Number of animals
To calculate the number of experimental units (single animal or pool of three organs for biochemical analysis) to be used, G*Power 3.1 (University of Düsseldorf, Germany) was used: α = 0.05, power of 0.85, and an effect size of 0.50, as determined based on previous data, 33 and considering clinical parameters as the primary outcome.
Statistical analysis
First, data were analyzed for normal distribution (Shapiro–Wilk test) and the homogeneity of variances between the groups (Levene's test). Data with a normal distribution are presented as the mean ± standard deviation (SD), whereas the remaining data are presented as the median [interquartile range].
The data pertaining to the time to the loss of equilibrium, reflexes to a painful stimulus, and opercular movements were log transformed, after which they exhibited a normal distribution and homogeneity of variances between the groups. One-way analysis of variance (ANOVA) with Tukey's test as a post hoc test was then used to compare these parameters between the treatment groups.
To analyze cortisol levels, data were log-transformed to achieve normal data distribution. Subsequently, two-way ANOVA using treatment groups and sex as factors and Tukey's post hoc test were used.
To assess the prevalence and the SGS gradation scale of the histopathological changes, the Kruskal–Wallis test was used to determine differences between the groups.
One-way ANOVA or the Kruskal–Wallis test was used to analyze the biochemical parameters and compare them between the groups. When data were normally distributed but did not exhibit homogeneity of variances between the groups, Welch's test was used.
All hypotheses were two-tailed tested, and statistical significance was set at p ≤ 0.05. Data were entered into Microsoft Excel™ 2010 (Microsoft Corporation, Redmond, WA) and analyzed using IBM SPSS™ 20 for Windows (SPSS, Inc., Chicago, IL). GraphPad Prism 7 for Windows (GraphPad, Inc., San Diego, CA) was used for graphical representations of the parameters without log transformation (raw data).
Results
Pilot study
All concentrations tested in the pilot study, as well as the number of dead animals, are presented in Table 2. The doses selected for the main study are indicated in bold. The first concentration clove oil tested based on the anesthetic concentrations 34 resulted in the death of all three animals; therefore, no further clove oil concentration was tested.
Anesthetic Concentrations Tested in the Pilot Study
The number of dead animals in each protocol is presented. The protocols selected to be used in the main study are presented in bold.
Max, maximum.
Euthanasia methods: main study
Regarding the loss of equilibrium, a significant difference was detected between the groups [F (4, 50) = 36.18, p < 0.001]. Rapid cooling induced a faster loss of equilibrium (8.0 [3.5] s) compared with the other treatments: Et group, 21 [5.8] s, p < 0.0001; CO group, 24.5 [11.0] s, p < 0.0001; P/L group, 25 [12.0] s, p < 0.0001; and MS222 group, 31 [11.0] s, p < 0.0001. MS222-treated animals exhibited a longer time to the loss of equilibrium than did zebrafish treated with etomidate (p = 0.004) or with propofol/lidocaine (p = 0.011) (Fig. 1A).

Latency to lose equilibrium
Moreover, the treatment groups had different latencies to the loss of reaction to a painful stimulus (Fig. 1B) [F (4, 50) = 140.39, p < 0.001]. Rapid cooling induced a faster loss of this reflex (27.5 [17.5] s) compared with the other protocols: MS222 group, 170 [98] s, p < 0.0001; CO group, 180.5 [114.0] s, p < 0.0001; P/L group, 260 [129.3] s, p < 0.0001; and Et group; 929.5 [1128.5] s, p < 0.0001. Etomidate-treated animals took longer to lose the painful reflex than did those treated with MS222 (p < 0.0001), clove oil (p < 0.0001), and propofol/lidocaine (p < 0.0001). In addition, MS222-treated animals lost this reflex faster than did propofol/lidocaine-treated animals (p = 0.025).
Regarding the cessation of the opercular movements (Fig. 1C) [F (4, 50) = 286.0, p < 0.001], RC induced a faster loss of these movements (30 [22] s) compared with the remaining treatments: MS222 group, 679 [390.3] s, p < 0.0001; CO group, 795 [250.3] s, p < 0.0001; P/L group, 1113 [588.75] s, p < 0.0001; and Et group, 1355 [995] s, p < 0.0001. Etomidate-treated animals took more time to stop breathing than did MS222- and clove oil-treated animals (p < 0.001).
Some zebrafish in all the groups still had a heartbeat after opercular movement ceased; however, none recovered or regained opercular movements when placed in clean water for 20 min. After this period, a heartbeat was not detected in any of the animals.
Cortisol assay
There was no interaction between the factors of sex and group, and no significant difference between sexes in relation to the cortisol values. The only difference detected between the treatment groups [F (3, 40) = 4.099, p = 0.013] was that the cortisol level was higher (p = 0.031) in the animals subjected to rapid cooling (10.4 [9.79] ng/g of body weight) compared with animals from the P/L group (5.9 [4.30] ng/g of body weight) (Fig. 2).
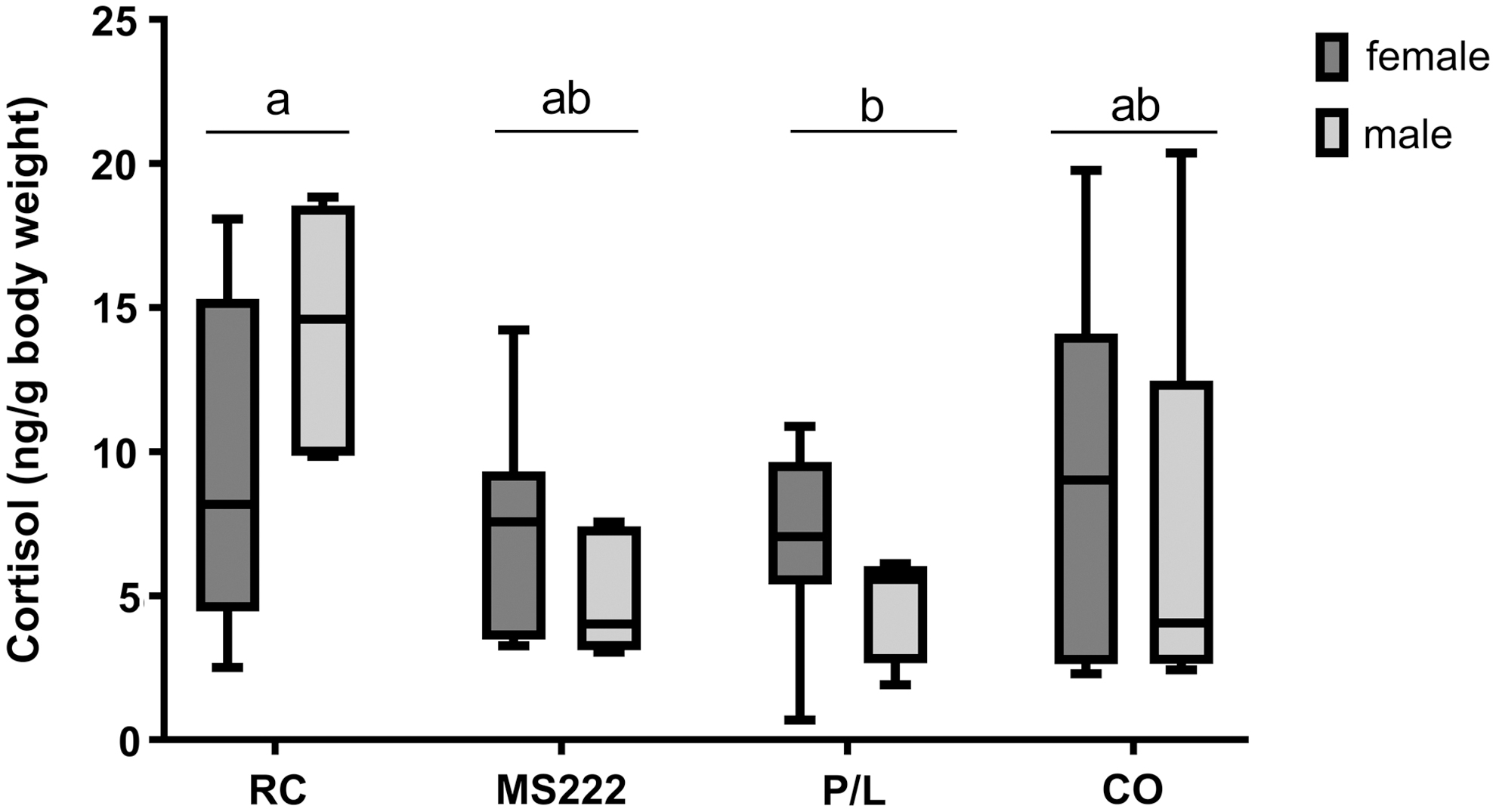
Whole-body cortisol levels (ng/g of fish body weight) determined in adult zebrafish after euthanasia. Animals exposed to cold water at 2°C–4°C (RC), 250 mg/L of MS222 (MS222), 20 mg/L of propofol combined with 100 mg/L of lidocaine (P/L), or 50 mg/L of clove oil (CO). The raw data are presented here as median and 5th to 95th data percentiles; n = 12: 7 females and 5 males in the RC, MS222, and P/L group, 6 females and 6 males in the CO group. Different letters indicate significant differences between the groups, p < 0.05, using one-way ANOVA, followed by Tukey's post hoc test using log-transformed data.
Biochemical analysis
One sample from the MS222 group was excluded from the analysis because it presented values out of the biological range; thus, for this group, n = 4 was used. Almost all the biochemical parameters assessed were similar among the groups (Supplementary Figs. S1–S4). The muscle analysis alone revealed one difference in ATPase activity between the groups [F (3, 15) = 3.793, p = 0.033]: a lower ATPase activity was detected in the CO group (2.7 ± 0.23 μmol Pi/mg protein) compared with the MS222 group (4.2 ± 1.12 μmol Pi/mg protein) (p = 0.046) (Fig. 3).

ATPase activity in the muscle of adult zebrafish after euthanasia. Animals exposed to cold water at 2°C–4°C (RC), 250 mg/L of MS222 (MS222), 20 mg/L of propofol combined with 100 mg/L of lidocaine (P/L), or 50 mg/L of clove oil (CO). ATPase activity is expressed in μmol of the inorganic phosphate (Pi) released by its hydrolysis per mg of protein. Data are presented as median and 5th to 95th data percentiles; n = 5, except for MS222 (n = 4); each sample is a pool of three organs. Different letters indicate significant differences between the groups, p < 0.05, using one-way ANOVA, followed by Tukey's post hoc test.
Histology
The main histopathological changes observed in the gills were edema, lifting, filament epithelium proliferation (FEP), and lamellar epithelium proliferation (LEP) (Fig. 4). Moreover, the prevalence of these changes was low and similar among animals from all the groups. Edema was observed in ∼30% of the animals (although an artifact of fixation or embedding cannot be ruled out), whereas epithelial lifting, FEP, and LEP were observed in less than 10% of the animals that were submitted to MS222, CO, and P/L. Regarding the SGS, no differences were recorded among treatments. In fact, the median severity degree of lifting, FEP, LEP, and edema was zero. Table 3 indicates the number of animals presenting each severity gradation by the total number of animals analyzed.
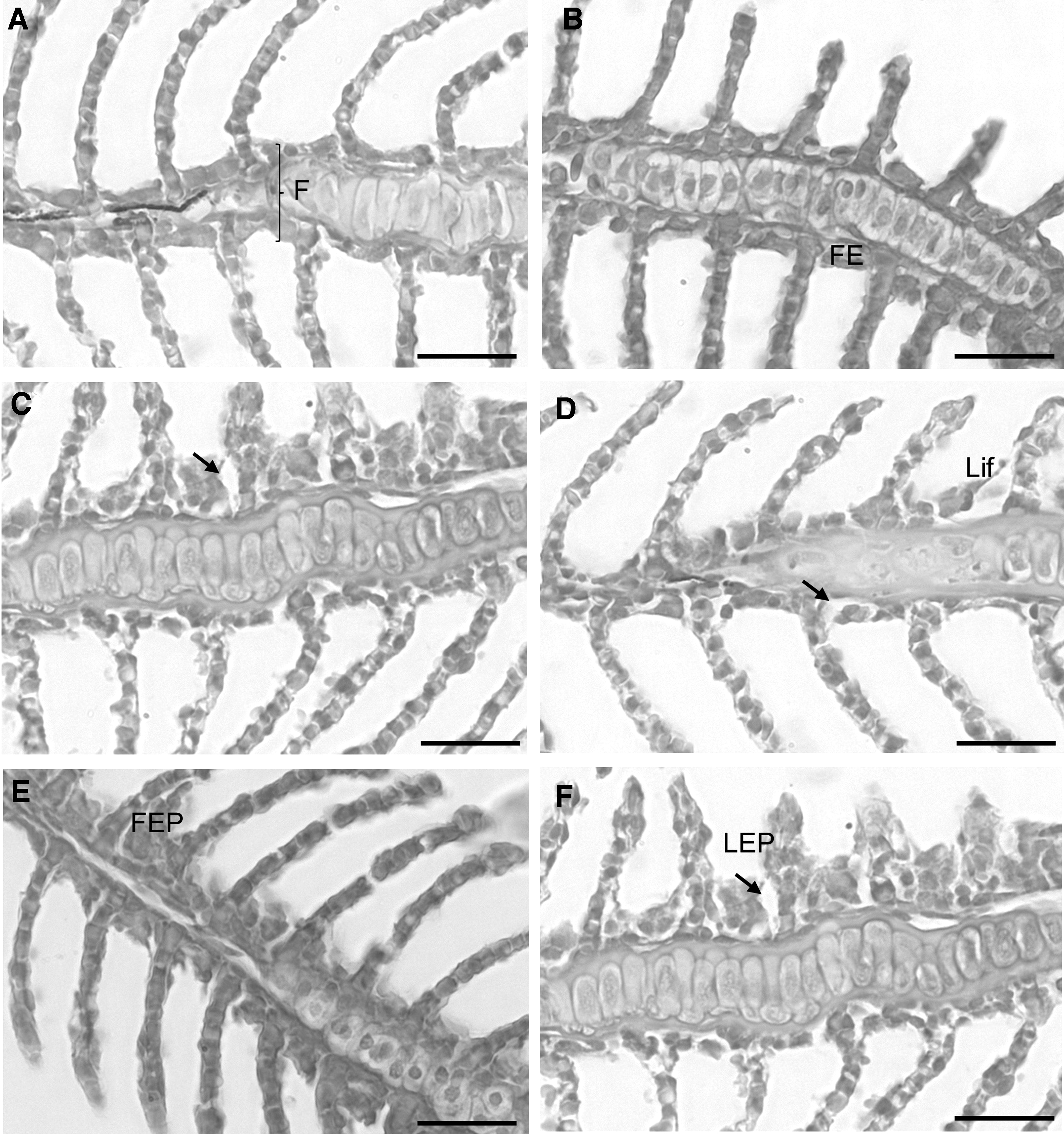
Images representative of the histopathological changes observed in the gill epithelia of fish.
Number of Animals Showing the Severity Gradation per Total Number of Animals Tested for Each Histopathological Alteration in Each Group
None presented severity gradation higher than 2. The animals not represented here had a degree of 0, that is, no relevant changes. Severity gradation: 1 = less than 25% of the filaments affected with the histopathological changes; 2 = 25%–50% of the filaments affected with the histopathological changes.
CO, 50 mg/L of clove oil; FEP, filament epithelium proliferation; LEP, lamellar epithelium proliferation; Lift, lifting; MS222, 250 mg/L of MS222; P/L, 20 mg/L of propofol combined with 100 mg/L of lidocaine; RC, cold water at 2°C–4°C.
The histological observations of the skeletal muscle sections revealed a normal fiber structure in fish from the different test groups, revealing the absence of histopathological changes or myofiber breakdown (Supplementary Fig. S5). The myofibers had an apparently uniform size and shape, fitting in a mosaic pattern. The fibers were separated by a thin sheet of endomysium. The flattened myofiber nuclei were normally located at the periphery of the cells, and the sarcoplasm was uniformly distributed. In longitudinal sections, striations produced by the sarcomeres were clearly visible.
Discussion
In this study, although all protocols used were efficacious to achieve euthanasia, rapid cooling was consistently the fastest method, with low variability among individuals; hence, it was more efficient. Moreover, none of the animals exhibited relevant side effects regarding the histological and biochemical values. All the groups had cortisol values similar to the most used euthanasia method, that is, an overdose of MS222.
In detail, etomidate was quick to induce a loss of equilibrium, although it was the slowest to induce death. As expected, etomidate-treated zebrafish took more time to lose the response to painful stimuli than did the remaining animals, as etomidate does not have analgesic properties. Nevertheless, this reflex was lost after around 30 min of exposure to etomidate because we used a very high concentration of this agent to euthanize zebrafish, and the loss of the pain reflex occurs when the animal is about to die. Regarding the latency to kill adult zebrafish, rapid cooling was significantly the fastest method (maximum latency of ∼0.98 min), followed by a similar performance recorded for the propofol/lidocaine combination, MS222, and clove oil; whereas etomidate was significantly slower (maximum latency of ∼40 min) than MS222 and clove oil in inducing death.
Furthermore, rapid cooling induced fewer variable responses (lower SD, variance, and interquartile range), and etomidate caused the least consistent responses among animals (higher interquartile range). Anesthetics are expected to be more prone to individual variability than is rapid cooling, as their effect will be affected by differences in animal size and in the amount of lipids at the sites where anesthetics may accumulate. 14 Variable results would be expected after clove oil treatment because of the variability in the eugenol content. 35 However, in our study, this was not observed as all the clove oil used came from the same batch (from Sigma–Aldrich). Rapid cooling is also expected to be slower in inducing death in larger zebrafish, although this was unlikely to be noticeable in the present study because we used adult zebrafish from the same strain, and the rapidity of the method renders small size differences irrelevant.
It is also worthy to point out that this study was conducted using the AB strain exclusively, which is one of the strains most used in research; thus, we advise running a pilot study when using other strains, to ensure the efficacy of the euthanasia method. Moreover, the identification of differences between sexes was not the aim of this study; therefore, sex was disregarded in almost all the parameters assessed. In fact, the sex of the animal was only considered in the cortisol analysis, as it is known that sex may affect cortisol release. 36
In this study, none of the animals recovered after 2 min without breathing. The AVMA guidelines 16 advise that adult zebrafish should be exposed to the euthanasia treatment for at least 10 min after opercular movement cessation. Although, in this study, none of the animals recovered after 20 min in clean water, the period of exposure included in the guidelines adds a safety margin to avoid accidental recovery. Performing a second/confirmatory method of euthanasia (e.g., decapitation or exsanguination) solves the question of the waiting time that is necessary after the last opercular movement to ensure the death of a fish. Heartbeat detection is not an indication that the fish will recover when placed in clean water, as we detected a heartbeat in many animals from all the groups after the 2-min period. This was an expected observation as the heartbeat is controlled by an intrinsic pacemaker system that continues to contract the heart after brain activity has ceased and only stops when oxygen is depleted. 37
Rapid cooling is accepted in the United States, 16 but not in countries following the European Directive 2010/63/EU, for the euthanasia of fish. This Directive does not distinguish between euthanasia methods for different fish species, and cold water is not effective for killing cold-water fish or large fish. In contrast, as we have demonstrated here, it rapidly kills zebrafish, which is a tropical species. However, we still need to determine what happens to zebrafish during a hypothermic shock, to fully understand if rapid cooling induces a “good death.” Studies have shown that the transient receptor potential V1 of the trigeminal ganglion neurons was activated by cold temperature in zebrafish larvae. 38 However, TRPA1 knockout zebrafish showed no behavioral alterations in response to cold, 39 thus, this mechanism warrants further research.
Although zebrafish do not need to prevent freezing in their natural habitat, they express antifreeze proteins 40 that are involved mainly in developmental processes. 41 Nevertheless, other defensive compensatory mechanisms have been detected in zebrafish under cold stress, such as alterations of gene expression. 42 We found a very quick loss of equilibrium in rapidly cooled zebrafish (average of 9 s), which leaves a very small window of time for the fish to express a stress response compared with the anesthetic overdoses tested.
Moreover, the preoptic area and pituitary pars distalis in the diencephalon, which are associated with temperature detection, 43 have not been directly implicated in the pain or fear/avoidance behaviors that are more related to telencephalon regions.44–46 In summary, the evidence that is available to date is not sufficient to understand what zebrafish feel during rapid cooling euthanasia; therefore, additional electrophysiological studies and other brain activity assessments would be important to understand if a true loss of consciousness is achieved or if the animal is only immobilized while still being able to experience stress.
To address this particular concern of stress, we evaluated the cortisol levels in animals that were subjected to the different euthanasia methods described previously, which did not seem to be affected by sex. Etomidate was not further evaluated because of the variability and long time required to induce death, which render it an impractical method. Rapid cooling resulted in increased cortisol levels, but only when compared with propofol/lidocaine overdose; thus, at least compared with the other methods, rapid cooling did not cause more stress to the animals. In carp, which is a temperate species, hypothermia was found to slow the drop in brain temperature by reducing the blood flow and oxygen to the CNS, with the exception of the areas associated with temperature detection and the endocrine stress response.
In fact, the plasma cortisol level was increased at 5 min after the temperature drop. 43 In the present study, the drop in temperature was much higher and death happened under 5 min; thus, the response to the potential stress induced by rapid cooling was probably not activated. It has been described that the cortisol levels in adult zebrafish start to increase within the first minutes following an acute stressor, although the peak is observed at around 15–30 min, depending on the stressor.47,48 Thus, it may be difficult to detect the release of cortisol caused by the rapid cooling stimulus, as the fish stopped responding to stimulus after 27.5 s on average. However, zebrafish subjected to an anesthetic overdose may have shown alterations in cortisol levels, as the animals had lost the response to all stimuli at 6 min after anesthetic exposure. Thus, cortisol levels were not increased in the anesthetic overdose groups because cortisol required more time to be released/detected or because the anesthetic overdose is not an effective stressor.
We cannot also rule out a possible blockage of cortisol release by an indirect action of the anesthetics, as they may inhibit important neurotransmitters that modulate the hypothalamic–pituitary–adrenal axis. 49 Regarding the cortisol levels, our results were similar to those obtained by measuring the serum cortisol levels of zebrafish euthanized with MS222 and rapid cooling. 50 However, in contrast with to our outcomes, Davis et al 35 showed that, in zebrafish, MS222 euthanasia induced higher levels of serum cortisol than did clove oil euthanasia, although the concentrations used in that study were rather high compared with the ones reported here (2300 mg/L for MS222 and 100 mg/L for clove oil). Nevertheless, other studies using rapid cooling showed few occurrences of behavioral indicators of distress in young adults, 17 or at least fewer occurrences compared with MS222 exposure in adult 5 zebrafish.
Propofol has been tested for euthanasia in zebrafish using higher concentrations (40 or 120 mg/L) and longer periods of exposure (20 or 10 min, respectively) than those used in our study, which could be impractical and increase stress 51 ; however, cortisol was not evaluated. Nevertheless, those authors did not record aversive behaviors. Lidocaine was also previously used to kill adult zebrafish successfully, with almost no aversive behaviors described.3,52 These results are in agreement with the low levels of cortisol detected in our study in animals treated with the propofol and lidocaine combination. The use of this combination allows the potentiation of the effect of each drug and the decrease in the concentration of each drug, thus preventing potential side effects, which is crucial when postmortem analyses are required.
Oxidative stress biomarkers are emerging as indicators in the evaluation of animal welfare, as exemplified in the study by Arechavala-Lopez et al, 53 because it can also be related to physiological stress and general health. 54 In our study, none of the oxidative stress parameters was altered between the groups, supporting the lack of relevant physiological stress differences. It can be postulated that no alterations of this type would be expected in a dying organism, where euthanasia methods render the animal dead within minutes; however, metabolic alterations were detected when different methods of euthanasia were used in mice. 55
Furthermore, no histological changes were detected in the gills induced after the treatments. This is very important as the gills are the organ that absorbs the anesthetics and is, thus, in close contact with the chemicals, which could induce damage, suffering, and interference with postmortem analysis. The proliferation observed in gill epithelia was probably not related to the euthanasia methods, as the cell cycle takes longer to be completed than does the process of euthanasia. However, assessing this feature was important to ensure that the fish studied did not have any structural damage in the gills, which could have been a variable in the biochemical analysis.
The muscle histology revealed an intact microarchitecture, excluding the possibility of muscle rupture in the animals that exhibited sudden movements of pronounced bending, which occurred especially when the rapid cooling method was used. Phenotypically, these bending movements were similar to C-start (Mauthner reflex), 56 which occurs when the animal is subjected to a stimulus, in this case cold water, and the trunk bends from the contraction of fast-twitch muscle fibers. However, in this work, fast swimming did not occur, as the animal stopped moving and death was rapidly achieved.
MS222 induced an increase of Na+/K+-ATPase activity in the muscle compared with clove oil. Clove oil and local anesthetics, such as MS222 and lidocaine, have been described to block sodium channels, among others. 57 The blockage of sodium channel prevents the influx of sodium ions into the muscle, thus preventing depolarization and impairing muscle contraction. 58 Therefore, the NA+/K+-ATPase activity is expected to decrease, leading to a decrease in ATP consumption. In fact, it has been reported that NA+/K+-ATPase activity decreased with the increase of the concentration of the inhibitor, such as lidocaine. 59
However, benzocaine, which has a similar structure to that of MS222, showed a biphasic effect, as it increased the activity of the ATPase up to a certain concentration, followed by a decrease at higher concentrations in mammals. 59 Thus, MS222 could still stimulate ATPase activity at the concentration used in the present study. In summary, the differences in ATPase activity detected here may be related to the chemistry of the different anesthetics and their concentrations. These ATPase activity alterations did not affect any other parameters; thus, it does not seem to cause a problem to the animals during euthanasia regarding the results of our postmortem analyses.
An optimal method of euthanasia is expected to comply with several criteria related to animal welfare; quality of scientific results when postmortem analyses are conducted; and the efficacy, safety, and practicability (procedure, costs, and availability) of the method. Regarding animal welfare, the P/L group alone exhibited lower values of cortisol compared with RC, and no major alterations were detected in the oxidative stress parameters or gill histology. All methods were considered efficacious as they provided irreversible euthanasia in all animals.
Regarding practicability, all the protocols allowed the euthanasia of several animals at the same time, with no need for special equipment, although etomidate took too long to induce death. MS222 and clove oil are slightly less practical to use, as these anesthetic solutions need to be prepared in advance: MS222 is a powder that must be diluted and buffered in a stock solution, and clove oil must be mixed with ethanol. As anesthetic baths need to be prepared with the appropriate dose, the margin for human error is higher when using these agents compared with the use of cold water.
Although all anesthetics used here can be easily bought, ice for rapid cooling is, of course, cheaper, thus representing an attractive method to aquarists/general public to use in ornamental zebrafish. For safety reasons, MS222 should be prepared in a fume hood. Although the risks associated with clove oil exposure have not been clarified, some of its components may have carcinogenic effects. 60 Rapid cooling has minimal occupational health and safety risks compared with chemical and other physical methods of euthanasia. 5
The purpose for which fish are killed is also important when choosing the euthanasia method. Although we did not detect relevant alterations in the parameters studied, if biochemical/molecular analysis needs to be performed or if the procedure already involves the use of other chemicals, a physical method, such as rapid cooling, might be better than anesthetic overdose, depending on the parameter to be tested. For example, based on the results of this study, to reduce the effect of euthanasia on cortisol levels, P/L overdose would be a better method as it induced lower cortisol levels than did the rapid cooling method. However, both groups had cortisol levels similar to the standard anesthetic overdose used (MS222 group).
Similar to the findings of Wilson et al, 5 who did not detect the formation of ice crystals in the tissues after the rapid cooling method, we did not detect any histological damage in this context. However, because rapid cooling is not a legally approved method in all countries, it cannot be used alone, and anesthetic overdose remains the most used method of fish euthanasia. To minimize the effect of anesthetic agents on the results of postmortem analyses, the concentrations used and the period of exposure to these agents may be reduced to provide loss of equilibrium, which can be followed by rapid cooling or decapitation. This protocol complies with the EU Directive, as other methods than those listed in the Annex IV may be used “on unconscious animals, providing the animal does not regain consciousness before death.”
Considering the issues studied here, rapid cooling seems to be a method that fulfills most of the criteria to properly kill adult zebrafish: rapid unconsciousness and death, requirement of minimum restraint, causing minimum stress (regarding the oxidative stress parameters and the cortisol levels compared with the standard MS222 method), reliability, reproducibility, irreversibility, simplicity of administration, and safety for the operator; moreover, it is economically viable. 2 Furthermore, rapid cooling did not interfere with the postmortem analyses performed here. Although propofol/lidocaine induced the lowest cortisol levels, more time was needed to induce death in the zebrafish; thus, the animals were exposed to the anesthetic for ∼18 min (on average), which may compromise the molecular analyses.
As rapid cooling is not allowed in the EU, we suggest that, after the loss of equilibrium induced by anesthesia, this method can be used to accelerate euthanasia, thus minimizing the duration of exposure to chemicals that may affect postmortem analyses, without requiring a special permit. In this situation, the anesthetic bath should be set at a temperature similar to the holding tank, so that the fish will face a real hypothermic shock when placed in the cold water and die rapidly, as described previously. The longer latency to lose equilibrium observed in the groups treated with anesthetic overdose compared with the rapid cooling group may increase the duration of the stress in zebrafish; however, we do not know what occurs in the brain during rapid cooling, mentioned previously.
If we choose a precautionary approach and assume that the loss of equilibrium in the rapid cooling group does not mean that the animal is unconscious, we can check the latency to the loss of the response to a painful stimulus (average of 27.5 s) or to the cessation of opercular movements (average of 30 s) as hallmarks of conscious experience loss in this group. These values were similar to the latency to lose equilibrium in the groups exposed to the anesthetic overdose (average of 24.5 s), which implies that they may all have experienced the same duration of euthanasia.
One of the limitations of this type of studies is the lack of a control group and/or baseline values regarding the parameters analyzed, as the animals have to always be killed to perform these biochemical, physiological, and histological analyses; thus, all values reported in the literature are from animals that have been killed using a specific method of euthanasia.
Further studies evaluating brain activity during euthanasia and other potential alterations induced by the euthanasia methods in postmortem analyses are needed. Although all the euthanasia methods studied here seemed to be reliable for this type of postmortem analyses, additional parameters can be affected by chemical compounds. Moreover, the potential stressful stimulus of euthanasia ended six times faster in the rapid cooling group (considering death) compared with the use of the standard MS222 overdose (considering loss of painful stimulus), with no cortisol increase, thus probably avoiding the activation of the stress response. In addition, rapid cooling has the advantage of being simple to perform, easily available, and affordable. Thus, there is a potential benefit for animal welfare and research in general if legislation is changed to include rapid cooling as a method for euthanasia for small tropical fish.
Footnotes
Authors' Contributions
J.M.F. conducted the experiments performed, analyzed data, and wrote the first draft of the article. L.F. and S.M.M. conducted experiments, analyzed data, and contributed to the preparation of the article. S.J. helped with data collection and reviewed the article. I.A.S.O. reviewed the article, providing critical feedback. A.M.V. conceived the experimental design, conducted the experiments, analyzed data, and gave important input and close supervision to the article writing. All authors discussed the results, contributed to the final article, and approved the article for publication.
Acknowledgments
The authors would like to thank José Bessa and the staff of the Zebrafish Facility for the maintenance of the tanks and care of the animals.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Norte Portugal Regional Operational Programme (NORTE 2020), under the PORTUGAL 2020 Partnership Agreement, through the European Regional Development Fund (FEDER) (Project Norte-01-0145-FEDER-000008—Porto Neurosciences and Neurologic Disease Research Initiative at I3S), and co-funded by FEDER funds through the Operational Competitiveness Programme—COMPETE 2020 and the Operational Competitiveness Programme and Internationalization (POCI-01-0145-FEDER-029542), Portugal 2020, and by National Funds through FCT—Fundação para a Ciência e a Tecnologia under the project PTDC/CVT-CVT/29542/2017. Individual funding was also provided by the doctoral fellowship SFRH/BD/135811/2018 issued by FCT to J.M.F.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.