
Abstract
Live food is necessary for the proper development of zebrafish larvae, providing nutrition in a form that is easily digestible and available to the larvae. Live food is commonly enriched to increase the dietary content of certain nutrients. However, little research has been done on protein-based enrichments, especially those of plant origin. This study sought to examine how different quality protein enrichments affected the composition of live food as well as growth and digestive tract development of larval zebrafish, Danio rerio. Larval zebrafish were fed from 3 to 22 days posthatch (dph) with one of six live food (rotifers Brachionus plicatilis and Artemia spp.) treatments: (1) live food with no enrichment (starved; control); (2) live food enriched with commercially used Spirulina spp. algae; (3) live food enriched with soybean meal (SBM); (4) live feed enriched with soy protein concentrate (SPC); (5) live feed enriched with a fishmeal hydrolysate; and (6) live feed enriched with intact fishmeal (FM). Proximate composition of live food was significantly affected by enrichment, in particular, protein content of rotifers was significantly increased by enrichment with SBM. Zebrafish fed SBM-enriched live food showed longer total body length than all other groups, except SPC. Zebrafish in the SBM group also showed increased gene expression of chymotrypsin in the intestine, possibly indicating improved intestinal development and extracellular digestion, which likely contributed to improved growth. Conversely, zebrafish fed hydrolysate-enriched live food showed reduced gene expression of alkaline phosphatase, possibly indicating a less developed intestinal tract, correlating with reduced growth compared to SBM group. Overall, plant protein was shown to be a promising source of live food enrichment for improving larval zebrafish growth.
Introduction
Most teleost fish species require use of live food at first feeding such as algae, copepods, rotifers (Brachionus spp.), and/or Artemia nauplii 1 since utilization of nutrients found in formulated pelleted diets is highly impaired by digestive tract immaturity1–3 and digestion of dietary protein is limited. 4 Ingestion of formulated feed particles can be difficult in these early stages, since physical properties such as lack of buoyancy, and lack of visual and chemical stimuli reduce ingestion compared to live food. 5
The early stage of larval fish development is a critical time and hence, it is imperative that the right diet characterized by high level of easily digestible protein required for optimal growth and survival is provided. The larval digestive tract is not fully developed or completely differentiated and undergoes drastic changes during the metamorphosis of larva into a juvenile.6,7 During this development, the presence or absence of exogenous and endogenous digestive enzymes will signal the maturity status of the digestive tract. Specifically, young larvae that do not have a fully developed digestive capacity will express higher levels of enzymes involved in intracellular digestion. 8 As the digestive tract develops, these intracellular enzymes will decrease in abundance as extracellular digestion takes over. 8 Larval fish grow fast during the first days/weeks of their life, and hence require delivery of highly available building blocks (amino acids) for efficient tissue protein synthesis.9,10
Fish larvae have evolved to feed on live organisms, which are able to deliver partially hydrolyzed nutrients that are more easily digestible compared to intact nutrients in formulated feeds. 1 Starved rotifers (Brachionus plicatilis) and freshly hatched Artemia contain 47% and 54% of water-soluble protein-derived nitrogen, 11 which is more available and easily taken up by the larval primitive intestinal wall, as opposed to intact nutrients found in formulated feeds, which require that digestive organs and enzymes are in place at the time of feed ingestion for proper nutrient breakdown.11,12 Live feed ingestion may also directly stimulate the release of digestive hormones and enzymes in the larval fish, which aid in the digestion process. 13
Although easily digestible and readily consumed, protein and amino acid content of live foods can vary,14,15 and is not consistent compared to formulated feeds. Differences in rearing methods and diets provided to rotifers will also alter their nutritional composition. 16 To overcome these nutritional deficits, live food is commonly enriched with various additives to compensate for their absence and/or to enhance fish health.17,18 Artemia nauplii enriched with plant oils have resulted in higher survival, total length, weight, and higher polyunsaturated fatty acid (PUFA) content of Sailfin molly (Poecilia latipinna) fry compared to those fed unenriched Artemia. 19
Furthermore, haddock (Melanogrammus aeglefinus) larvae fed either rotifers enriched with algae or Artemia enriched with commercial PUFA emulsion had significantly higher weight and survival than larvae fed formulated microdiets. 20 In general, a variety of live food enrichment strategies have been tested in larval fish nutrition.17,21 However, most of these studies focus on enrichment using lipid and fatty acid sources to meet requirements for those particular essential nutrients in the young fish and are not aimed at amino acid delivery for enhancement of protein tissue synthesis. Comparatively, a limited number of live food studies have been conducted using a protein-based enrichment. Among those, Johannsdottir et al. 22 found that Atlantic cod (Gadus morhua) can achieve a more distinguished organ structure when fed rotifers and Artemia nauplii enriched with a fish protein hydrolysate through bioencapsulation (the coating of feed particles with a waterproof polymer).
In addition, when Pejerrey (Odontesthes argentinensis) larvae were given Artemia enriched with a hydrolyzed yeast product, they achieved significantly higher weight and specific growth rate compared to those given unenriched Artemia. 23 Thus, the available evidence seems to suggest that the use of live food enriched with a protein source provides an efficient stimulation of the digestive tract toward proper maturation and positive growth performance. The use of plant protein (PP) is of particular interest, given the fish culture industry's desire to use alternative protein sources to those of marine animal origin. 24
In general, protein sources can be divided into two different groups: high and low quality. High-quality protein sources are characterized by high protein content, high digestibility, high palatability, balanced amino acid profile, and lack or low level of antinutritional factors that have been associated with inducing intestinal inflammation. Examples of those include fishmeal (FM) and its derivatives, as well as refined PP sources such as soy protein concentrate (SPC) or wheat gluten. Low-quality protein sources are often of plant origin and include unrefined plant meals that present opposite qualities to those of high quality (i.e., imbalanced amino acid profile, lower digestibility, and high concentration of antinutritional factors).
However, they are more cost-effective and widely available. Therefore, the objective of this study was to determine the effect of live food enrichment with different quality PP sources on the growth, survival, digestive tract development, and proximate composition of larval zebrafish (Danio rerio). Specifically, SPC, FM, and fishmeal hydrolysate were used as high-quality PP sources, while soybean meal (SBM) was tested as a low-quality PP. Spirulina was also included since it is commonly used as a feed source for rotifers. Unenriched rotifers served as a control in the study. We hypothesized that low-quality PP would be a suitable live food enrichment source for larval zebrafish, which would result in improved growth performance and digestive tract development status compared to fish fed unenriched live food.
Materials
Fish and culture system
This experiment was performed in accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of Southern Illinois University-Carbondale (SIUC), and all protocols (protocol #18–007) were approved by SIUC Institutional Animal Care and Use. All fish were reared indoors in a closed recirculating system (Pentair Aquatic Ecosystems, Cary, NC) consisting of racks of 3 L clear plastic tanks. Every treatment and parameter was tested in triplicate.
Water in the system was pumped through a carbon filter, treated with UV light and a biofilter before being aerated, and pumped back into the fish tanks. Water temperature during the experiment was an average of 26.73°C ± 0.07°C. Sodium bicarbonate and solution of salt were automatically added into the system to maintain the pH at an average of 7.01°C ± 0.08°C. Conductivity was at an average of 2479.76 ± 68.26 μS cm−1. The higher conductivity level was chosen to increase lifespan of the live food during feeding in the experimental tanks. 25
Zebrafish were SIUC domesticated animals that originated from broodstock stocked to the SIUC system in 2018. Zebrafish were of a nonstandard genetic line and were sourced from a local pet store (Petco, Carbondale, IL). Males and females were separated and fed ad libitum 2–3 times per day with a combination of Artemia nauplii and formulated commercial feed (Otohime, Japan). The fish were then all combined into one tank with a mesh frame and fake plant to act as an artificial substrate and as a barrier between the eggs and spawning adults to prevent cannibalization. Adult fish were removed after spawning, while eggs were left in the tank to hatch. Once larvae hatched and reached the swim up phase at ∼2 dph, they were randomly distributed 10 at a time into new 3 L tanks, with an initial density of 95 larvae per tank.
Exogenous feeding began at 3 dph, larvae were fed approximately every half to 1 h for the first two days ad libitum during daylight hours. The photoperiod was set to 10 h light:14 h dark. The illumination provided was at 245 lux and the distance between the surface of the water and the light source was 10 cm. This feeding regime continued from 3 dph to 5 dph, after which larvae were fed every 2–3 h for the duration of the experiment. The feeding trial lasted until fish reached juvenile stage at 22 dph. The total biomass of each tank was weighed at 23 dph (fish were starved for 24 h before handling) by gathering every fish in the tank into a net and then padding onto a paper towel thrice to remove excess water before being weighed on a scale.
Twenty fish from each tank were randomly chosen to be measured for total body length. Three replicates were used per treatment, resulting in 60 fish in total, which were measured for length, and three replicates of total biomass per treatment was measured. These fish were euthanized in an overdose of MS-222 (100 mg/L) before being measured, after which they were patted dry, placed in a tube, and stored at −80°C until further use in analyzing gene expression and proximate composition. A total count of all fish in each tank was also made to calculate the average weight of each fish in the tank and to assess survival.
Experimental treatments
The larval fish were divided into six groups and the dietary treatments included the following: (1) live food with no enrichment (starved; control); (2) live food enriched with commercially used Spirulina spp. algae (Earthrise, Irvine, CA) commonly used for feeding rotifers; (3) live food enriched with soybean meal (SBM; solvent extracted; Premium Feeds, Perryville, MO; low-quality PP option); (4) live food enriched with soy protein concentrate (SPC; The SausageMaker, Inc., Bufalo, NY; high quality PP option); (5) live food enriched with a fishmeal hydrolysate (hydrolysate; produced in house from muscle rendered from wild Bighead Carp); and (6) live food enriched with intact FM (mechanically extracted carp meal; Illinois River, Marseilles, IL). Both FM sources were considered high-quality marine protein options. Treatments were randomly assigned to tanks and three replicate tanks per treatment group were used. 26
The live feed feeding regime consisted of rotifers Brachionus plicatilis from 3 dph to 5 dph, followed by a combination of rotifers and Artemia nauplii (6–9 dph) and Artemia nauplii feeding only (10–22 dph). All groups were fed to apparent satiation to ensure high feed intake by larval fish.
Live food enrichment
Rotifers were reared in 22 L buckets at ∼15 ppt salinity with constant aeration at 25°C. Twice a day, rotifers were fed a mix of Spirulina spp. algae (Earthrise, Irvine, CA) and yeast (Lesaffre, Mexico), which had been previously homogenized to sustain the culture. Dry Artemia cysts were hatched during 24-h incubation under constant light in 6 L McDonald jars with 30 ppt salinity (Instant Ocean Sea Salt, Blacksburg, VA) aerated water at 25°C.
For rotifers and Artemia, equal volumes were each separated into individual McDonald jars to be enriched. Enrichments were made by homogenizing the protein source with de-ionized water at a ratio of 1:20 by weight of protein source to water and then straining through a 150 μm filter. Each enrichment was poured into a sterile plastic tube for storage and frozen at −20°C until ready for use. The night before each feeding, one tube of enrichment was moved from the freezer to a refrigerator to defrost overnight.
The next morning, the thawed enrichments were briefly homogenized once again to ensure proper mixing and to remove larger solids, if any, and then added to each jar containing the live feed. A minimum volume of enrichment was added such that the protein concentration of each jar was at least 88 mg/L. 22 Composition of raw materials used to make enrichments is presented in Table 1.
Proximate Composition of Enrichment Products
Composition (% dry matter) of raw materials used to make enrichments.
Solvent-extracted soybean meal; Premium Feeds, Perryville, MO, USA.
Soy protein concentrate; The SausageMaker, Inc., Buffalo, NY.
Bighead Carp, produced in-house from muscle rendered from wild Bighead Carp; Illinois River, Marseilles, IL.
Mechanically extracted carp meal; Illinois River, Marseilles, IL.
Food grade; Earthrise, Irvine, CA.
Before starting the feeding trial, a preliminary examination of rotifers and Artemia nauplii provided with different enrichments proved that the enrichment was ingested. 27 Five hundred milliliter beakers of rotifers and Artemia nauplii were enriched with SBM and left with an air stone. For 2 h, starting after the initial time of enrichment, the live food was sampled at 30-min intervals. The live food was pipetted onto a slide and examined under a microscope (Nikon Instruments, Tokyo, Japan) and visually inspected to determine its viability and if the enrichment had been taken up. Changes in coloration in the body and visual identification of enrichment in the guts were confirmed and compared to unenriched food. The optimal enrichment length established was 2 h. Live food viability was also assessed after the enrichment by visual observation of rotifers and Artemia nauplii in their culture.
Biochemical analysis
Enriched and unenriched rotifers and Artemia nauplii were siphoned onto a 50 μm mesh filter and allowed to dry, then scraped into a conical Falcon tube, and freeze dried for later proximate composition analysis (Freezone 6; Labconco). For proximate composition of whole zebrafish body, 15 fish from each tank, which had been previously frozen at −80°C, were gradually thawed before being weighed (sample wet weight), and then homogenized (Powergen 1000; Fisher Scientific) in 3 mL of de-ionized water. The samples were then freeze dried (Freezone 6; Labconco) before weighing again (sample dry weight).
An FP-528 Nitrogen/Protein Analyzer (LECO Corporation, St. Joseph, MI) was used to measure crude protein content in dry live food and fish samples by combustion in accordance with AOAC International (2000). Using a modified method described by Folch et al., 28 lipid content was analyzed in samples using a chloroform-methanol extraction.
Gene expression analysis
The relative expression of the following genes was analyzed: alkaline phosphatase (alpi.1), chymotrypsin like (ctrl), trypsin (prss1), elastase II like (ela2l), and tumor necrosis factor alpha (tnfa). The analysis of gene expression followed the methods of Molinari et al. 29 Briefly, five zebrafish from each tank, previously frozen at −80°C, were gently thawed and their heads and tails were removed, and the remaining portion of the body was processed with TRIzol Reagent (Ambion, Foster City, CA). RNA was extracted with On-Column PureLink™ DNase Treatment (Pure-Link RNA Mini Kit and PureLink DNase; Invitrogen, Carlsbad, CA) following the manufacturer's instructions. Using a spectrophotometer (Nanodrop 2000c; Thermo Fisher Scientific, Waltham, MA), nanograms/μL of each RNA sample was obtained.
Two μg from each of these samples was then reverse transcribed using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA) to make a 20 μL cDNA solution, which was then added to 380 μL of water. The qPCR mixtures for each tank contained 400 μL of 2 × SYBR qPCR mix and 5 μL each of forward and reverse primer (20 μM stock). Each qPCR reaction was run in duplicates on a 96-well plate. The qPCR cycle consisted of 95°C for 10 min, followed by 40 cycles of 95°C for 20 s and 60°C for 35 s, culminating in a dissociation curve. A reference gene (ef1a) was used to normalize target gene expression. Relative expression for target genes was calculated using the 2ΔΔCt.
Statistical analysis
The average weight and average total body length were each grouped by treatment and then analyzed using a one-way ANOVA followed by a Tukey's multiple comparison test to determine if significant differences were present between treatment groups. A value of p < 0.05 was considered significant. Survival for each tank was calculated as the percentage of fish remaining from the initial stocking of 95 fish per tank. Survival values are presented as the average of three survival rates per treatment. Survival was statistically analyzed using a Tukey's multiple comparison test to determine the presence of any significant difference between groups. The same statistical analysis was used for gene expression, using the ΔCt value for each gene to calculate any significance between groups. Statistical analysis was done with R software (RStudio, PBC, Boston, MA). All values are presented as the average ± standard deviation.
Results
Fish growth performance
Enrichment of live food with SBM significantly increased body length (p < 0.05) relative to control, hydrolysate, FM, and Spirulina groups, but not compared to SPC (p > 0.05). SPC enrichment increased body length (p < 0.05) compared to hydrolysate and control groups. Enrichment with hydrolysate, FM, or Spirulina did not increase body length (p > 0.05) compared to control. No difference in body weight was found between any group (p > 0.05).
Live food proximate composition
Proximate composition of the live food is presented in Table 2. SBM-enriched rotifers had the highest protein content compared to the nonenriched group (control) and SPC-enriched group (p < 0.05). Rotifers enriched with other treatments were excluded from proximate analysis due to low rotifer count at sampling. SBM-, SPC-, and Spirulina-enriched Artemia did not have protein content different from control (p > 0.05). The FM-enriched Artemia nauplii had the highest protein content compared to all other groups (p < 0.05). Protein content was the lowest in Artemia enriched with hydrolysate and was significantly different from control (p < 0.05)
Live Food Proximate Composition
Proximate composition (% dry matter) of Artemia and rotifers enriched with various additives. Rotifers and Artemia nauplii were enriched for 2 h. Values are presented by enrichment as averages with standard deviations. Different letters indicate statistical significance at p < 0.05.
SBM, soybean meal; SPC, soy protein concentrate.
The lipid content of SPC-enriched rotifers was significantly lower than control (p < 0.05), while SBM group did not differ from control (p > 0.05). FM and Spirulina were the only enrichments to increase lipid content in Artemia relative to control (p < 0.05).
Ash content of rotifers was not affected by SPC (p > 0.05), but was lowered by SBM compared to control (p < 0.05). The ash content of Artemia was significantly different between all groups (p < 0.05; Table 2).
Fish survival
Enrichment of live food with SBM did not significantly increase survival compared to control or any other treatment group (p > 0.05; Table 3), nor did the use of SPC, hydrolysate, or FM increase survival relative to control (p > 0.05). A significant reduction in survival (p < 0.05) compared to control was found in the Spirulina group.
Average Body Weight, Total Body Length, and Survival of Zebrafish
Values are presented by diet as averages with standard deviations. Different letters indicate statistical significance at p < 0.05.
Fish whole-body proximate composition
No significant difference (p > 0.05) between treatments was found in whole zebrafish body lipid and protein contents (Table 4). Due to little sample material obtained after freeze-drying, moisture and ash were not analyzed.
Zebrafish Whole-Body Content
Average whole-body protein and lipid content (% composition) by treatment. Values presented are the average of whole-body lipid and protein content per treatment with standard deviations.
Relative gene expression
Digestive enzymes
The genes analyzed and their primers are presented in Table 5 and relative expression is presented in Figure 1. Enrichment of live food with SBM did not increase relative expression of ela2l relative to control or any other treatment group (p > 0.05). Use of SPC, hydrolysate, or Spirulina did not cause a significant difference (p > 0.05) in ela2l relative expression compared to control, while FM resulted in a significant reduction of relative expression (p < 0.05) compared to control. SBM, SPC, or Spirulina did not significantly affect (p > 0.05) relative expression of alpi.1 compared to control. FM did not affect (p > 0.05) alpi.1 expression either; however, hydrolysate caused a significant reduction (p < 0.05) compared to control.
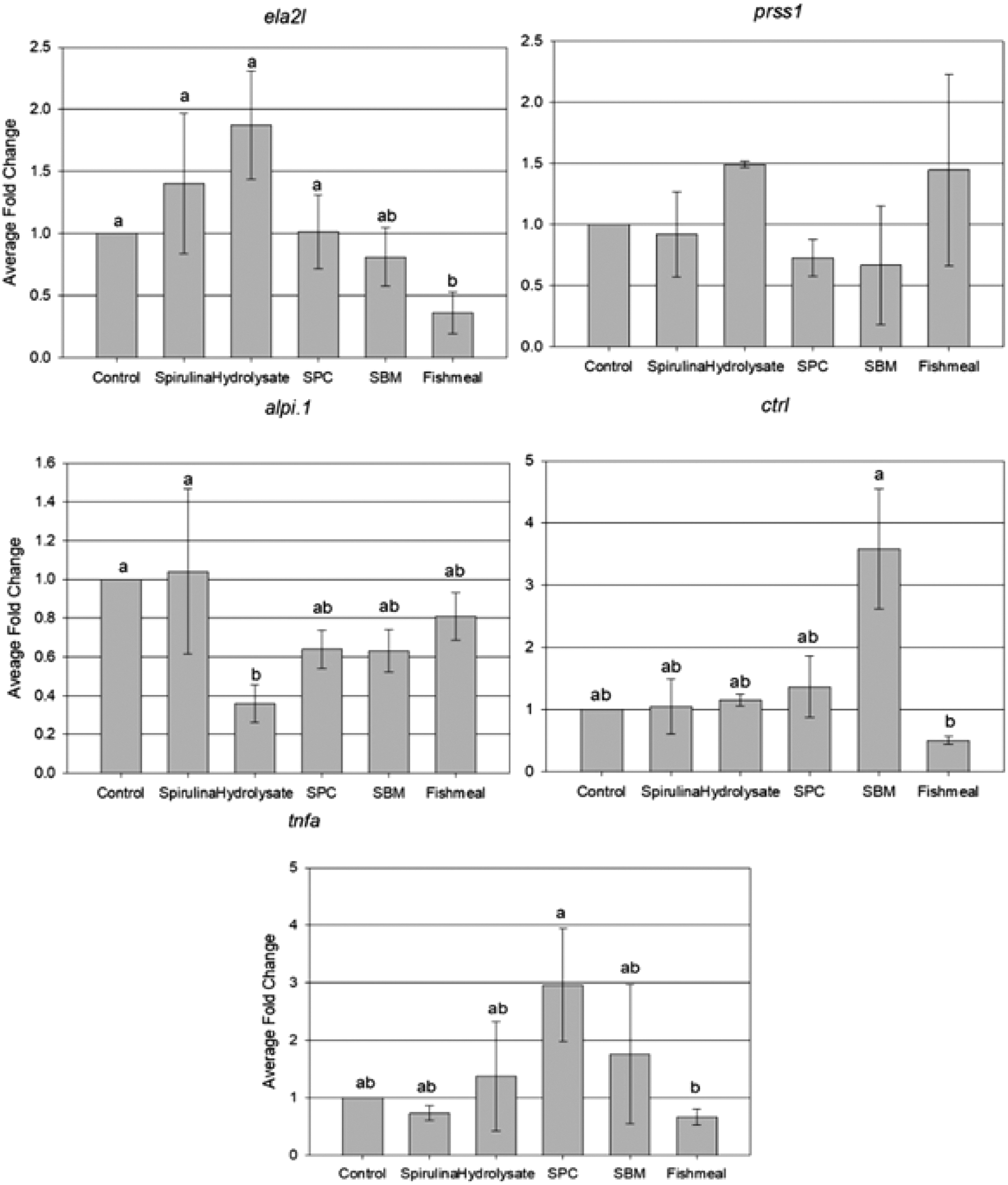
Relative fold changes of genes analyzed in zebrafish. Presented are elastase, trypsin, alkaline phosphatase, chymotrypsin, and tumor necrosis factor alpha coding genes: ela2l, prss1, apli.1, ctrl, and tnfa in each treatment group relative to the control group. A reference gene (ef1a; fold change = 1) was used to normalize target gene expression. Values are mean fold change with standard error of the mean. Significant differences between groups (p < 0.05; presence of significance determined by one-way ANOVA and least square difference test) are represented by different letters. Different letters indicate statistical significance at p < 0.05.
Primers Used in Gene Expression Analysis
Primers used for qPCR analysis of zebrafish digestive tract.
Enrichment with SBM resulted in a threefold increase in expression of ctrl compared to control. This increase was not significant (p > 0.05) compared to control, but was significant compared to FM (p < 0.05). Expression of ctrl did not differ from control (p > 0.05) in SPC, hydrolysate, FM, and Spirulina treatments. When analyzing the data for ctrl expression, two data points were identified as outliers. A replicate in the SBM and FM groups had relative fold changes of over 90% and 216%, respectively. No other replicate resulted in a fold change of ∼5%, and only two replicates were above 2%. With these outliers included, no significant difference between treatments in ctrl expression was detected (p > 0.05). No significant difference between groups was found in the relative gene expression of prss1 (p > 0.05).
Inflammatory cytokines
Use of SBM or SPC did not significantly affect relative expression of tnfa compared to control, nor did the use of hydrolysate, FM, or Spirulina enrichments (p > 0.05). FM enrichment caused a significant reduction in tnfa expression compared to SPC (p < 0.05); however, this was only achieved after an outlier in the data was removed (similar to the outlier discussed in relation to ctrl). This replicate of the SBM group showed an individual fold change of ∼59%. Only four replicates across all groups showed an individual fold change greater than 2%.
Discussion
Growth performance and proximate composition
This zebrafish study showed that fish larvae fed live food enriched with low quality SBM had significantly longer total body length compared to the control (starved) or live food enriched with hydrolysate, FM, or Spirulina, but did not differ from high-quality SPC. We believe this observed improved growth performance of zebrafish larvae fed SBM-enriched live food resulted from higher crude protein content of rotifers after the enrichment process. The higher protein content of rotifers specifically is important, since this is the first food provided to the fish when larval protein requirements and growth rate are at their highest.9,10
Effective growth during the larval stage requires high levels of protein in the diet and increased protein content in enriched rotifers during the first days of feeding likely provided a head start on growth for the SBM group, allowing them to achieve a longer final length. While Artemia enriched with other products (Spirulina and FM) had similar or higher protein levels than SBM, this was at later stages in zebrafish development where growth is comparatively slower and so the protein utilization differs as well.9,10 These results would therefore indicate SBM's potential as an enrichment product for first feeding larvae. Providing high dietary protein as early as possible may be a significant advantage in achieving longer body length later in life.
Live food enrichment and its positive effect on fish growth have been reported before30–32 ; Putra et al. 33 enriched Daphnia sp. with SBM and found it contained significantly higher levels of protein than those in other enrichment groups (manure and coconut dregs flour), resulting in significantly higher weight gain and specific growth rate in climbing perch (Anabas testudineus) larvae fed for 15 days. This is similar to results obtained in this zebrafish study, which found that SBM-enriched rotifers had significantly higher protein content compared to the nonenriched (control) and SPC-enriched groups. Øie et al. 34 reared first feeding turbot (Scophthalmus maximus) larvae in tanks either with or without algae, feeding them either enriched (with commercial products) or unenriched rotifers.
Rotifers exposed to algae increased their protein content over 72 h, and regardless of algae presence in the water, the highest larval growth rate was achieved in groups fed rotifers with the highest protein content. The concept of live food enrichment has been shown to effectively alter the nutritional composition of live food and henceforth affect fish growth. The effects on growth due to dietary PP can vary depending on inclusion level and species,35,36 but the higher growth of our zebrafish on SBM-enriched live food would suggest that SBM is not hindered as a live food enrichment source, despite being considered “low quality.”
Body length results similar to SBM were found in the “high-quality” SPC treatment. SPC has higher protein content (crude % by mass; Table 1) and is much more refined, lacking the same level of antinutritional factors present in SBM, which are known to impede the growth of fish. 37 A typical SPC production process will remove oligosaccharides and saponins, and inactivate trypsin inhibitors.38–40 However, SPC can have a protein content over 60%, but protein dispersibility in water under 6%. 41 It is possible that when SPC enrichment was prepared using the method presented in our study, these SPC proteins were not fully dispersed in the enrichment tanks (although not visually observed), and were therefore not fully available to the live food. This may explain the lower protein content of rotifers enriched with SPC compared to SBM and the unenriched control, despite the higher protein content of SPC itself.
Similarly, the Artemia enriched with high-quality hydrolysate had a lower protein level compared to SBM-enriched Artemia and resulted in lower total body length. High inclusion of hydrolyzed proteins in larvae has been shown to not always be beneficial. In Senegalese sole (Solea senegalensis) larvae, the highest dietary inclusion level of protein hydrolysates did not generate any noticeable positive effect to the larvae. The intact protein diet and the diet with moderate inclusion level of hydrolyzed protein both showed better growth at different stages. 42
The authors suggested that the high amount of hydrolyzed protein was likely saturating the intestinal peptide transport system and decreasing the overall protein uptake. 42 Hydrolysates are composed of low-molecular-weight peptides and individual amino acids, which are highly hydrophilic, and hence, poor growth of larval fish associated with high dietary inclusion level of hydrolysates is often associated with nutrient leaching.43,44 It is possible that the water-soluble components of the hydrolysate leached out from the enrichment before their ingestion by the live food or the high assimilation rate of low-molecular-weight peptides prevented proper utilization of hydrolysate enrichment by the zebrafish larvae.
In contrast, another high-quality marine protein source, FM, seems to have been readily taken up by Artemia, having the highest protein content of all groups. This did not translate to longer larval body length, again indicating that perhaps increased protein content is most effectively utilized for growth, and therefore more important, earlier in development. This is reflected again by high protein content of SBM-enriched rotifers.
The Spirulina enrichment did not increase total body length compared to the unenriched control, although specific algae enrichments and their different combinations have been shown to be effective in other studies. Barramundi (Lates calcarifer) larvae that were fed rotifers enriched with a 50:50 ratio of Nannochloropsis oculate and Chlorella vulgaris algae showed longer total body length compared to those fed rotifers enriched with other ratios of the algae.
Fish larvae fed live food enriched with the 50:50 mixture also had greater depth of body (measured vertically from the anal vent) at 10 and 13 dph compared to other groups. 45 Different mixtures of algae may be required for effective fish growth, especially considering the lower survival of Spirulina group compared to control. A mixture of algae that produces a higher protein content than Spirulina alone may be beneficial. Dicentrarchus labrax larvae fed algae-enriched Artemia nauplii achieved higher total length when the algae used for enrichment had higher protein and fatty acid content. 46
Differences in both lipid and ash content of our live food were found; however, these trends did not seem to correlate with growth, and it appears that total dietary lipid or ash was not a limiting factor on our zebrafish. The lipid content of our control and SBM-enriched rotifers did not significantly differ from each other, and were both above 10% total lipid. The FM-enriched Artemia was the only group to be above 20% lipid.
Larval cod has shown increased standard body length when fed enriched rotifers with lipid content of 7.8% ± 0.7%, 47 possibly suggesting that lipid source and fatty acid composition are more of an important factor than total lipid amount. The SBM-enriched rotifers had the lowest ash content compared to control and SPC-enriched rotifers, although this difference was small. Rotifer ash content by percent dry matter is unaffected by enrichment with Chlorella algae, Chlorella with zinc, DHA protein Selco, 48 or starvation. 49 Ash content of rotifers enriched with lipid emulsion, zinc, or combination of the two did not reach above 3%, 48 while in this zebrafish study, ash content in rotifers was above 16% in all groups. The ash content of Artemia in this study is similar to Artemia enriched with soybean oil and fish oil treatments by Morais et al. 50
Fish fed Spirulina-enriched live food had the lowest survival and was the only group to differ from control. Our survival results (81.05% ± 4.23% for all groups averaged together) are comparable to other reports in the literature on the same species. Carvalho et al. 51 found that zebrafish larvae fed from 6 to 27 days postfertilization (dpf) with unenriched Artemia nauplii had survival of 86% ± 5%, significantly higher than survival of zebrafish fed on commercial and purified diets. Zebrafish larvae raised in water mixed with algae and fed rotifers showed survival of 75% at 12 dpf and 80% at 22 dpf. 25
Goolish et al. 52 found first feeding zebrafish reared on live paramecia and Artemia can achieve survival of 69% after 21 days, and up to 94% depending on water turnover regime and food availability. Overall, we do not believe that the enrichment of live food presents a risk to survival greater than what already exists in the literature. This study found no significant differences in final average weight or whole-body composition among zebrafish groups after feeding from 3 dph to 22 dph.
Digestive enzyme gene expression
Alkaline phosphatase
The relative expression of alpi.1 in SPC, SBM, and FM groups did not differ from another group. The relative expression of alpi.1 found in zebrafish fed hydrolysate-enriched live food was significantly lower than those fed control and Spirulina-enriched live food. Using alpi.1 as a marker of intestinal maturity, the hydrolysate group seems to present a less mature status in terms of digestive capacity compared to control and Spirulina groups, despite no significant difference in growth performance between those groups. This is similar to findings from Johannsdottir et al., 22 who reported a more developed foregut in Atlantic cod larvae fed hydrolysate-enriched live food, although this result was not correlated with higher body weight or longer fork length compared to larvae fed lipid-enriched live food.
As larval fish develop, the activity of specific digestive enzymes changes, reflecting the developmental status of their digestive system. 53 Before the full maturation of the digestive tract, food is mainly broken down by intracellular digestion (often expressed as leucine-alanine peptidase activity). As the larvae grow and the stomach and intestine mature and differentiate, intracellular digestive enzyme activity decreases, while extracellular digestion takes over (expressed as activity of trypsin, chymotrypsin, carboxypeptidase, and others53,54). Change in the digestive system can be also seen in the expression of alkaline phosphatase (alpi.1), a brush border enzyme used as a marker of enterocyte development and intestinal maturation. 55
Alkaline phosphatase activity has been shown to be positively correlated with the proliferation of enterocytes in the fish intestinal lining. 55 This increase in alkaline phosphatase is often correlated with a decrease in cytosolic enzymes, such as leucine-alanine peptidase, reflecting a change from intracellular to more mature extracellular digestion.3,8,56 Several species showed significant increase in alkaline phosphatase expression after 250 degree days of growth. 57 Martinez et al. 58 found a gradual decrease in alkaline phosphatase starting from the development of exocrine pancreas in Senegalese sole. However, this trend reversed at the end of larval metamorphosis, and alkaline phosphatase rose sharply once a fully functioning stomach was present.
Chymotrypsin and trypsin
The expression of proteases, such as trypsin and chymotrypsin, can indicate increased ability of fish to perform extracellular digestion of dietary protein. Our study found that fish given SBM-enriched live food achieved a longer total body length at 23 dph compared to those given unenriched live food, or food enriched with Spirulina, FM, or hydrolysate. Larval fish have high protein requirements early in life; hence, efficient delivery of amino acids is needed for tissue protein accretion during the first days of feeding.9,10,59 The improved total body length observed in zebrafish was likely associated with higher protein content of rotifers enriched with SBM compared to the other groups. Consequently, these fish likely achieved higher extracellular digestion activity (reflected by increased ctrl expression) and continued to show good growth ahead of majority of the remaining groups until the juvenile stage.
Although the increase of ctrl expression in SBM group was not significant compared to control, it was trending very close to significance (p = 0.0514) and was over threefold greater than control. In addition, a significantly longer body length in SBM group compared to FM was found, which correlated with a significant increase in ctrl in the SBM group compared to FM. We therefore believe this result to be worthy of consideration. As the digestive tract of the fish matures, a higher expression of extracellular digestive enzymes is observed, 8 regulated by genes such as chymotrypsin (ctrl), responsible for breakdown of proteins. This increased ctrl expression in the SBM group could suggest more reliance of the larvae gut on extracellular digestion compared to the other groups, possibly indicating a more developed state of the zebrafish intestine.
Based on this evidence, SBM could be a promising protein enrichment to provide zebrafish with a head start on growth and development. Higher protein at the earliest stages (during rotifer feeding) likely increases protease (ctrl) activity, which in turn increases protein utilization when growth rates are highest. Furthermore, a more mature status of the digestive tract development would likely translate to more efficient utilization of formulated diets during and after weaning process, resulting in a better growth performance compared to fish from other groups.
This study found no difference in the expression of trypsin (prss1). Guerrera et al. 60 suggested that trypsin is the main proteolytic enzyme in early development of zebrafish; however, the rate of trypsin expression in early stages may not be easily affected by dietary history. Trypsin-specific activity can also increase at significantly higher rates compared to chymotrypsin in zebrafish after 22 dph. 60 Perera and Yúfera 61 found that SPC- and SBM-based microdiets given to zebrafish during the first three days of feeding (before switching to FM-based diets) did not affect trypsin expression up to 30 dpf.
Elastase
The ela2l gene codes for a peptidase precursor, an inactive zymogen. 62 In its activated form, the elastase 2 enzyme is a serine peptidase that aids in extracellular digestion, hydrolyzing elastin and proteins in connective tissue.63,64 Overall, the elastin results seem somehow counterintuitive. Groups with significantly higher ela2l expression (control, Spirulina, hydrolysate, and SPC) may have had improved digestion of dietary protein than the FM group, which presented the lowest expression compared to all the remaining groups apart from SBM. FM is naturally high in connective tissue; however, it does not seem to have led to a significant increase in ela21, nor does the higher dietary protein content of FM-enriched Artemia seem to positively correlate with ela2l expression.
Inflammatory cytokine expression
The utilization of dietary soy products, particularly those of low quality, in formulated feeds is usually limited due to the presence of antinutritional factors that can induce intestinal inflammation at high dietary inclusion levels.65–67 To evaluate the effect of SBM and SPC enrichments on fish gut inflammation, the expression of tumor necrosis factor alpha (tnfa), a gene that codes for proinflammatory cytokine produced as part of the fish immune response,68–70 was analyzed. The expression of tnfa in zebrafish fed SPC-enriched live food was significantly higher compared to the FM-enriched group. This is surprising, given that SPC is considered to be a high-quality product, which has been processed to remove various anti-nutritional factors that cause intestinal inflammation.
The low-quality SBM treatment did not show a significant increase in tnfa expression compared to control or other high-quality products, FM and hydrolysate. Similar results have been seen with the use of formulated diets. Zebrafish raised on FM diets, and then given either SPC- or SBM-based microdiets from 30 dpf to 50 dpf, showed significant increase and no change in tnfa expression, respectively, compared to zebrafish fed FM during the same period. 61 The hydrolysate, Spirulina, and FM enrichments notably do not have the antinutritional factors found in some PP.
Impaired growth performance has been seen in fish given formulated diets with high levels of PP,71–74 which contain antinutritional factors. 37 A difference in tnfa expression was, however, not correlated with any change in total body length and body weight between the SPC and FM groups. It is difficult to evaluate the full extent of intestinal inflammation based on a single gene, and future evaluations should use histological analyses. However, we do believe that PP provided through enriched live food likely does not deliver antinutritional factors at a level high enough to cause reduced growth in larval to postlarval zebrafish. The potential ways in which live food affects these antinutritional factors should be investigated further.
A final note should be made regarding the genetic line of zebrafish used. It is understood that various behavioral and growth effects can occur as a result of strain choice,75–77 in this case, a pet store-purchased strain was used. However, replication with other strains may lead to different results and therefore, any extrapolation of results from this study should be done with caution.
Conclusion
The results from this study indicate that live food can be enriched to increase its protein content through plant-based products. SBM specifically is a promising enrichment to be used toward improved total length of larval zebrafish during the first feeding. While dietary PP has been shown to reduce growth at high dietary inclusion levels, this has not been found in zebrafish larvae when SBM was used as a live food enrichment. The more developed digestive tract reflected by increased expression of ctrl in the SBM group likely contributed to the positive growth performance in that group. Similar growth to SBM group was seen in the SPC group. The use of SBM as an enrichment, however, is benefitted by its relatively low cost and wide availability.
The effect of live food on the antinutritional properties of PP enrichments and their inflammatory effects requires further investigation. Both Spirulina and fish hydrolysate did not seem to be suitable live food enrichments for zebrafish. Finally, rotifers and Artemia nauplii are effective vehicles for delivering enrichment protein to zebrafish larvae, and further investigation of how different PP sources are processed and affected by live food organisms is warranted.
Footnotes
Authors' Contribution
Conceptualization: K.K. and M.W.; data curation: K.K. and C.S.; formal analysis: K.K., C.S., and G.S.M.; funding acquisition: K.K.; investigation: K.K., M.W., C.S., and G.S.M.; methodology: K.K. and M.W.; project administration: K.K. and M.W.; resources: K.K.; software: K.K.; supervision: K.K.; validation: K.K. and M.W.; visualization: K.K.; writing–original draft: C.S.; and writing–review & editing: K.K., M.W., C.S., and G.S.M.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.