
Abstract
Severely skewed sex ratios in zebrafish stocks can pose significant hurdles for line propagation and sperm cryopreservation. To overcome female-biased sex ratios in stocks derived from imported sperm samples, the Zebrafish International Resource Center has implemented routine supplementation of larval food with 17α-methyltestosterone to skew gonadal sex differentiation toward masculinization. Resulting stocks averaged 80% males.
The Zebrafish International Resource Center (ZIRC) serves biomedical research as a central repository of wild-type, mutant, and transgenic zebrafish (Danio rerio) lines. Most alleles (>90%) in ZIRC's genetic inventory have been imported as sperm samples cryopreserved from multiallelic lines, each of which carry up to 30 distinct mutations.1,2 Because ZIRC imported only a few sperm samples per line, these must be routinely amplified by cryopreserving additional samples from stocks rederived at ZIRC. To facilitate timely distribution, the first generation of a multiallelic stock is cryopreserved as pooled sperm samples from 20 males.
This sperm is then used to fertilize eggs for line shipment. A challenge has been a significant female bias in rederived stocks, especially for lines imported from other facilities in which various genetic backgrounds, husbandry protocols, and genetic selection criteria are used. To overcome this limitation, ZIRC implemented the dietary supplementation of a synthetic androgen, 17α-methyltestosterone (17α-MT), in larval food to skew gonadal sex differentiation toward males.
Zebrafish sex determination is influenced by genetic and environmental factors.3–5 The administration of exogenous hormones to control gender has been described for many species including zebrafish,4,6 typically resulting in fertile animals. A literature review of 17α-MT treatment methods showed that immersion and dietary administration have been used in zebrafish and revealed significant variability in applied concentration, exposure onset age, and duration, leading to variable outcomes for gonad differentiation (Supplementary Table S1).7–13
Dietary administration of 17α-MT is predominately utilized in commercial aquaculture. 14 For dietary supplementation, the hormone is dissolved in ethanol before homogeneous mixing with the diet, followed by ethanol evaporation. The ZIRC food preparation procedure is outlined in Figure 1A and detailed in Supplementary Materials.
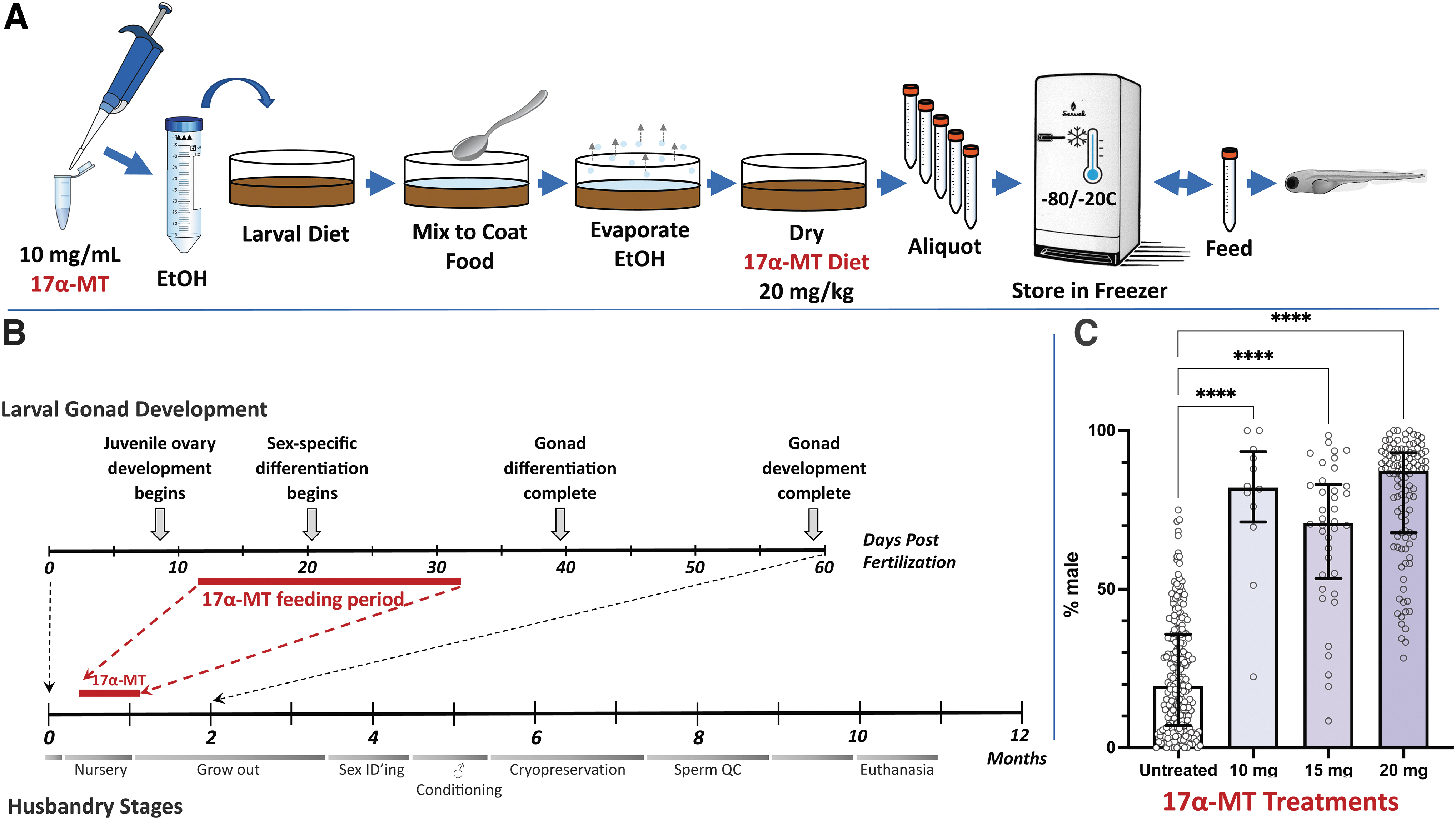
Skewing stocks toward male.
In general, fish are more sensitive to steroid treatment when gonads are still undifferentiated.6,14 Thus, the period of zebrafish gonad differentiation and hormone treatment needs to be correlated during juvenile rearing. In the ZIRC nursery, larval fish are fed paramecia between 5 and 10 days postfertilization (dpf). Subsequently, they are fed powdered larval diet (Zeigler AP mix; see Supplementary Protocol—Food Preparation) twice per day and artemia twice per day up to 32 dpf. Coordinating the dietary supplementation of 17α-MT with the powdered diet in the nursery (10–30 dpf) corresponds best with the bipotential period of zebrafish gonad differentiation (Fig. 1B).
To this end, we placed up to 50 rederived AB/TL larvae (AB, ZL1,
Using visual inspection of sexually dimorphic characteristics, we observed that food dosing concentrations from 10 to 20 mg/kg produced predominately male stocks (Fig. 1C; Supplementary Data S1). The median of untreated stocks (n = 228 populations; average population size: 127 ± 44 fish) was 19.5% males. In contrast, the median of 17α-MT-treated test groups was 82.1% males for 10 mg/kg (n = 12 populations, average population size 47 ± 3 fish), 71% males for 15 mg/kg (n = 38 populations, average population size: 79 ± 31 fish), and 87.4% males for 20 mg/kg 17α-MT (n = 108 populations, average population size: 78 ± 32 fish). Animal survival rates (94.7% ± 7% survival; see Supplementary Data S1) were not significantly affected by 17α-MT treatment.
Larval fish were exposed to 17α-MT on a recirculating mechanical and biological water filtration system with UV sterilization, 17 isolated from untreated fish to protect these from unintended masculinization. The 17α-MT added to the treated water system varied daily with the number of treated stocks (typically 5–20 tanks) and ranged from 4 to 16 μg/day. Diluted in system water, this amounted to ∼0.3–1.3 ng/L 17α-MT. Testosterone in water is transformed by biodegradation and photodegradation.17–19
Importantly, the sex ratios of wild-type control stocks on the treated recirculating system remained normal. The daily water exchange rate at ZIRC varies from 12% to 20%, and used water is discharged to the municipal sewer system. The amount of 17α-MT utilized and discharged at ZIRC did not require monitoring by the University of Oregon Environmental Health & Safety or local municipality.
The half-life of 17α-MT in fish food declines linearly over time and at higher temperatures. 20 Consistently, we observed that lower 17α-MT concentrations (i.e., 10–15 mg/kg) and increased food storage times produced inconsistent male conversion rates. Therefore, we recommend that 17α-MT-treated food be aliquoted in small volumes (8–10 mL), stored frozen (−80℃ to −20℃), protected from light, and used within 6 months.
Footnotes
Acknowledgments
We thank ZIRC staff for animal care and feeding.
Authors' Contributions
J.L.M. conceived and supervised the project and prepared original draft. J.M. performed experiments and data collection. A.N. managed data curation and validation. Z.V. performed statistical analysis and data visualization. All authors contributed to review and editing of article.
Ethical Treatment of Animals
Research and animal care were approved by the University of Oregon Institutional Animal Care and Use Committee, AUP# 21-15.
Disclosure Statement
No competing financial interests exist.
Funding Information
Zebrafish International Resource Center (ZIRC) is supported by an NIH grant (P40 OD011021) from the Office of Research Infrastructure Programs (ORIP) in collaboration with the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.