
Abstract
Loricariidae (Siluriformes) comprises ∼1026 species of neotropical fish, being considered the most diverse among the Siluriformes. Studies on repetitive DNA sequences have provided important data on the evolution of the genomes of members of this family, especially of the Hypostominae subfamily. In this study, the chromosomal mapping of the histone multigene family and U2 snRNA was performed in two species belonging to the Hypancistrus genus, Hypancistrus sp. “pão” (2n = 52, 22m + 18sm +12st) and Hypancistrus zebra (2n = 52, 16m + 20sm +16st). The presence of dispersed signals of histones H2A, H2B, H3, and H4 in the karyotype of both species, with each sequence displaying a varied level of accumulation and dispersion of these sequences between them was observed; in addition, U2 snDNA probe only showed positive results in H. zebra, which present this multigene in the terminal region of three chromosomal pairs. The obtained results resemble data already analyzed in the literature, in which the action of transposable elements interfere in the organization of these multigene families, in addition to other evolutionary processes that shape the evolution of the genome, such as circular or ectopic recombination. This study also shows that the dispersion of the multigene histone family is quite complex, and from this, these data serve as a point of discussion for the evolutionary processes that occur in the Hypancistrus karyotype.
Introduction
Repetitive DNA sequences constitute a large part of the genome of eukaryotic organisms, in which they can be located at multiple loci randomly (such as transposable elements) or form in tandem arrays in specific regions of the chromosomes (satellite DNAs and multigene families). 1 Histones constitute a complex multigene family responsible for coding five protein classes (histones H1, H2A, H2B, H3, and H4) that act in compaction of nucleosomes and chromatin, as well as in the regulation of gene expression. 2 They present variable number of copies that can be organized in sites located in one or more chromosomal regions, or disseminated throughout the genome, forming incomplete clusters or isolated genes.3–5 Also noteworthy is the family of U snRNA genes, subdivided into five gene classes (U1, U2, U4, U5, and U6), that act in the RNA splicing process.6–8
Although studies on the mapping of multigene families in the fish genome are still limited, interesting features have already been observed, such as the interaction with other multigene families, influence on karyotype diversification, different patterns of distribution in chromosomes, and interference of transposable elements.9–23
Loricariidae is a family of neotropical fish recognized for its high diversity, with 1026 valid species. 24 At present, Loricariidae is divided into six subfamilies, of which Hypostominae has the highest species richness. 25 Hypancistrus is a neotropical genus, comprising eight species, 26 being its type species, Hypancistrus zebra Isbrücker and Nijssen, 1991 endemic from the Rio Xingu region. They are highly sought after and marketed as ornamental fish, because of their exuberant coloration patterns. Such color patterns are one of the few morphological characteristics that allow the identification and differentiation of this genus members.27,28
Cytogenetic studies in Hypancistrus are still scarce, at the moment, it has been observed that this group presents 2n = 52, indicating a highly conserved karyotype, a characteristic observed in other members of the tribe Ancistrini (except in the genus Hypostomus, which has a wide variation in diploid number 29 ), with different karyotypic formulas and absence of heteromorphic sex chromosome.30,31 In most Hypancistrus species, the mapping of repetitive sequences is restricted mainly to 5S and 18S rDNA and transposable element Rex-3.30,31 Considering the conserved nature of the chromosomal macrostructure of Hypancistrus, the physical mapping of repetitive sequences will provide useful information about the karyotype evolution of this genus.
In this context, this study's objective was to physically map different multigene families, including H2A, H2B, H3, and H4 histone sequences, as well as U2 snRNA in two species of Loricariidae, belonging to the Hypancistrus genus (H. zebra and Hypancistrus sp. “pão”), to observe the organization pattern of these repetitive sequences and comprehend its dynamics in the genome of this genus.
Materials and Methods
The samples used in this study were composed of six male specimens of H. zebra and six male specimens of Hypancistrus sp. “pão.” All individuals sampled came from the municipality of Altamira, Pará, Brazil (Fig. 1). Taxonomic identification was performed according to current literature. 28 The specimens were collected with authorization from the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio), SISBIO no21078.

Map indicating sample collection locations of Hypancistrus analyzed cytogenetically in this study.
Chromosomal material was obtained from fish anterior kidney cells, according to the protocol described in previous studies. 32 Chromosomes were stained with Giemsa 5%, and analyzed in an optical microscope. Five metaphases from each individual were used to calculate the chromosomal measurements using the DRAWID software. 33 To assemble the karyotype, we followed the morphological classification of the chromosomes. 34
The C banding was performed as described by Sumner, 35 with some alterations. Slides were put in 0.2 HCl for 10 min at room temperature and then washed in distilled water. Next, they were put in 2% barium hydroxide at 60°C for 14 s, and immediately washed in 0.1 HCl at 60°C for 10 s. After that the slides were incubated in 2 × SSC at 60°C for 20 min. Finally, the slides were stained with Giemsa, and the results were observed on the microscope.
The genomic DNA of adult Hypancistrus sp. “pão” were extracted using the GenElute Mammalian Genomic DNA Miniprep kit (Sigma-Aldrich, St Louis, MO). The sequences used in the production of probes for fluorescent in situ hybridization (FISH) analysis were amplified by PCR with the following set of primers: H2A (H2AF 5′-AAG AAA AAC CRG CGG AAA RG-3′ and H2AR 5′-CTT GAC GGB CTT CTC GGY C-3′), H2B (H2BF 5′-AAG AAR GCC GTG ACC AAR AC-3′ and H2BR 5′-ACT TGG AGC TDG TGT ACT TGG-3′), H3 (H3F 5′-ATG GCT CGT ACC AAG CAG AC(ACG) GC-3′ and H3R 5′-ATA TCC TT(AG) GGC AT(AG) AT(AG) GTG AC-3′), H4 (H4F 5′-GGT AAG GGA CTC GGY AAA GG-3′ and H4R 5′-GCC GAA GCC RTA CAG RG-3′), and U2 snRNA (5′-TCT CGG CCT (AT) (AT) T GGC TAA-3′ and 5′-G(AC)G GTA (GC) TG CAA TAC CGG-3′).
Sets of primers for the amplification of histones H2A, H2B, and H4 sequences were produced in Primer3 Plus software, 36 based on sequences from other fish species deposited in NCBI, whereas primers described by Colgan et al 37 were used in the isolation of histone H3 and U2 snDNA sequences. The PCRs of the sequences were performed as follows: 1 cycle of 95°C for 5 min, followed by 35 cycles of 95°C for 1 min, 50°C–55°C for 50 s, 72°C for 2 min, a cycle of 72°C for 10 min and then a hold at 4°C. All PCR products were purified before sequencing using the illustra™ ExoStar™ 1-Step Kit. Sequencing was performed by the Sanger method, on the AB3500 platform, and the sequence identity was analyzed by the BLAST tool at the NCBI. The CENSOR tool 38 was used to identify possible insertions of transposable elements in the obtained sequences.
The probes of histones H2A, H2B, H3, and H4 were marked with nick-translation with biotin-11-dATP and detected with avidin-CY3, and the probes of histones H2B and H4 were marked with digoxigenin-14-deoxyuridine triphosphate (dUTP) (Dig-Nick Kit; Roche) and detected with anti-digoxigenin-fluorescein isothiocyanate (FITC). U2 snDNA probe was produced by PCR labeling using digoxigenin-11-dUTP and detected with anti-digoxigenin-FITC. FISH was performed following the protocol of Pinkel et al 39 with the following alterations: chromosomes were treated with Pepsin solution, PBS and dehydrated in alcohol battery (70%, 90%, and 100%). For chromosomal denaturation, the slides were immersed in a 70% formamide solution at 65°C, and the probes were denatured at 70°C for 15 min on thermocycler.
Slides were kept overnight at 37°C for hybridization. Subsequently, slides were washed with 50% formamide, 2 × SSC, and 4 × SSC-Tween to remove nonspecific hybridizations. Chromosomes were counterstained with 4-6-diamidino-2-phenylindol (DAPI) in VECTASHIELD antifading solution (Vector). Images were captured using an AxioCam MRm CCD camera (Nikon, Tokyo, Japan) attached to a Nikon H550S epifluorescence microscope (Nis-Elements AR program [Nikon]) and the Nis-elements software; all images were photographed using 100 × magnification objective lenses.
Results
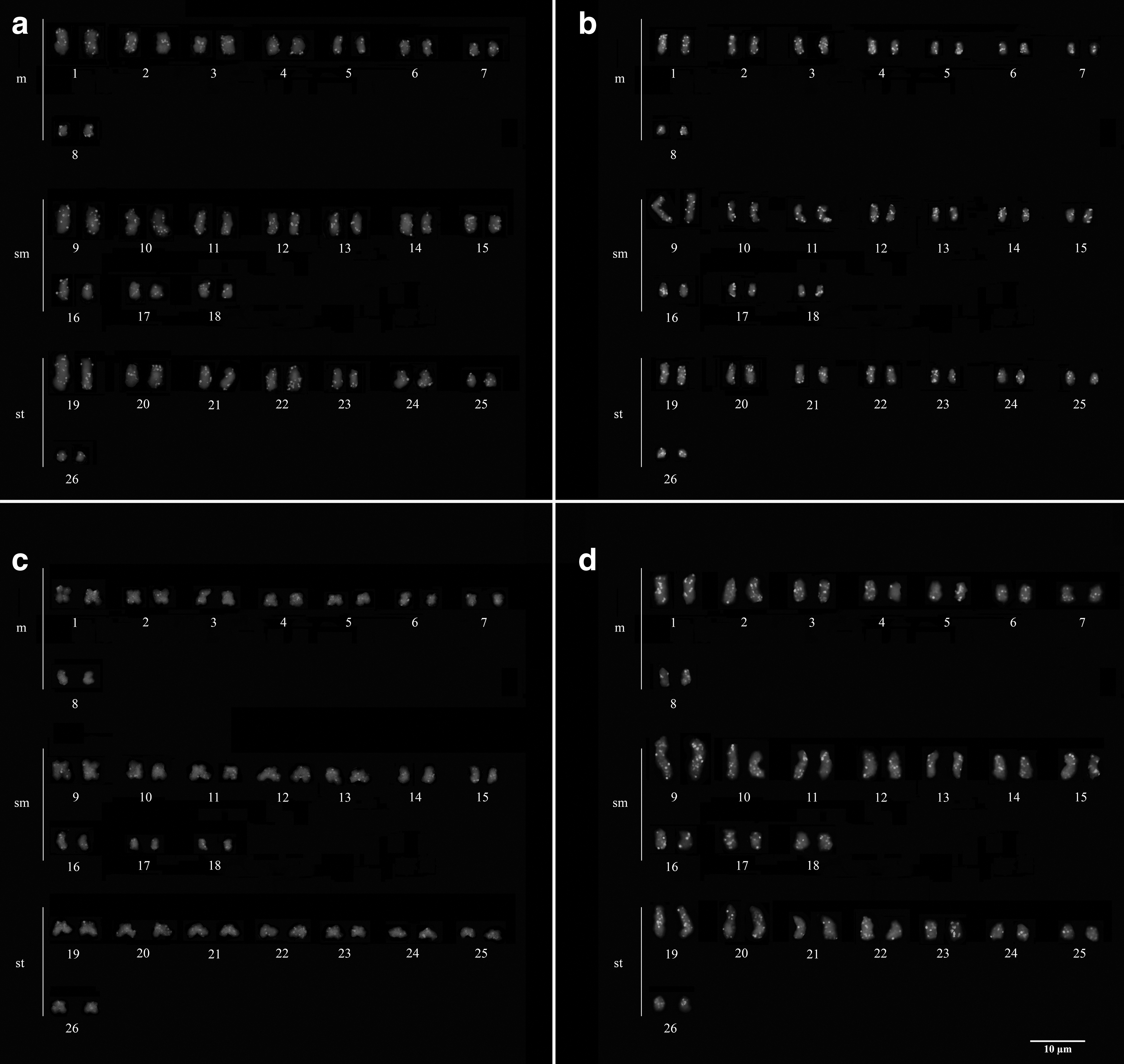
Cytogenetic analysis and chromosomal mapping of H2A, H2B, H3 and H4 histone sequences were performed in the karyotype of two species of Hypancistrus. Hypancistrus sp. “pão” presented 2n = 52, FN = 104, and karyotypic formula of 22m+18sm +12st (Fig. 2a) and H. zebra presented 2n = 52, FN = 104, and karyotypic formula of 16m+20sm +16st (Fig. 2b). C-banding analysis revealed heterochromatin blocks in both species. Hypancistrus sp. “pão” displayed heterochromatin blocks in terminal regions of short arms of pair 3 and in the long arms of pair 18, as well as in the long arms of homologous chromosomes of pairs 2 and 12 (Fig. 2c). Meanwhile, H. zebra displayed heterochromatin blocks in the long arms of pairs 10 and 12 (Fig. 2d).

Karyotype Giemsa stained
The mapping of histone sequences showed a similar distribution pattern between both species analyzed. H. zebra showed H2A signals highly dispersed by its karyotype, with no concentration of this multigene in specific regions (Fig. 3a), whereas Hypancistrus sp. “pão” showed signs of H2A along the chromatids, with small levels of concentration of this sequence being observed in terminal and pericentromeric regions of the chromosomes (Fig. 4a). H2B also showed scattered signals with a small heteromorphic cluster identified on a chromosome in both species (Figs. 3b and 4b).

FISH in H. zebra with histone probes
FISH in Hypancistrus sp. “pão” with histone probes
H. zebra also showed scattered signals of histone H3, with a small cluster located in the terminal region of two acrocentric chromosomes (Fig. 3c), whereas Hypancistrus sp. “pão” showed histone H3 signals dispersed along the chromosomes (Fig. 4c). As for the mapping of H4, a subtle degree of accumulation in the terminal regions of the chromosomes of H. zebra (Fig. 3d) was observed, and in Hypancistrus sp. “pão” the presence of small signals of this multigene, both in euchromatin and in pericentromeric heterochromatin was observed; however, a certain amount of signals from these genes were more concentrated in terminal regions of the chromosomes (Fig. 4d).
Owing to its dispersed chromosomal distribution, we chose to analyze the sequence of histone genes obtained in this study. Histone amplicons for H2A, H2B, H3, and H4 genes showed, respectively, 288, 235, 260, and 200 bp in size. After BLAST analysis, the histone H2A sequence produced in this study showed 90.28% identity with histone H2A from Tachysurus fulvidraco (XM_027177019.1), among other fish species; gistone H2B sequence showed 94.47% identity with histone H2B from Pangasianodon hypophthalmus (XM_027177019), among other fish species; histone H3 sequence showed 99.62% identity with histone H3 from Hypostomus ancistroides (JX680316.1), among other fish species; and histone H4 sequence showed 91.5% identity with histone H4 from Pygocentrus nattereri (XM_037530990.1), among other fish species. CENSOR analysis of these sequences revealed no insertion of transposable elements.
U2 snDNA probe only showed positive results in H. zebra. H. zebra has clusters of this multigene in the terminal regions of pair 1 (metacentric), in the short arm of pair 10 (submetacentric) and in the long arm of pair 19 (subtelocentric) (Fig. 5).

FISH with snRNA U2 probes in H. zebra.
Discussion
H. zebra and Hypancistrus sp. “pão” share the same diploid number 2n = 52, indicating that this is a conserved characteristic for this group of fish.30,31 Despite this, the two species have different karyotypic formulas, indicating that although they have a conserved karyotypic macrostructure, small changes in their chromosomal morphology are present and come from chromosomal rearrangements such as inversions.30,31
In situ localization of U2 snDNA in H. zebra revealed clusters in three chromosome pairs. Data on the mapping of U2 snRNA gene demonstrate that this multigene is commonly organized in a cluster located on a chromosome pair in most of the fish analyzed14,19–21; despite this, studies have shown the presence of U2 snRNA gene signals in several chromosomal pairs, as observed in Gymnotus pantanal, 19 in five species of Astyanax, 40 and in M. merluccius 41 ; in members of the Batrachoididae family, this multigene showed a widely dispersed distribution along the chromatids. 10 More than one site for U1 snRNA gene have been reported in Peckoltia. 42 The diversification of the number of chromosomal sites of U2 snRNA genes is mainly related to transposition events because these multigenes can generate pseudogenes by retrotransposition of their RNAs, as suggested for U4 snRNA in Ancistrus. 43 Thus, it is possible that part of the multiple U2 snRNA sites in H. zebra originated from a similar event.
In some fish, U2 snRNA gene may present syntenic localization with ribosomal genes. 20 However, in H. zebra this phenomenon does not occur because these sequences are located on distinct chromosomal pairs as given in Figure 6, which compiles the distribution of repetitive DNAs in the karyotype of H. zebra described in the current literature.
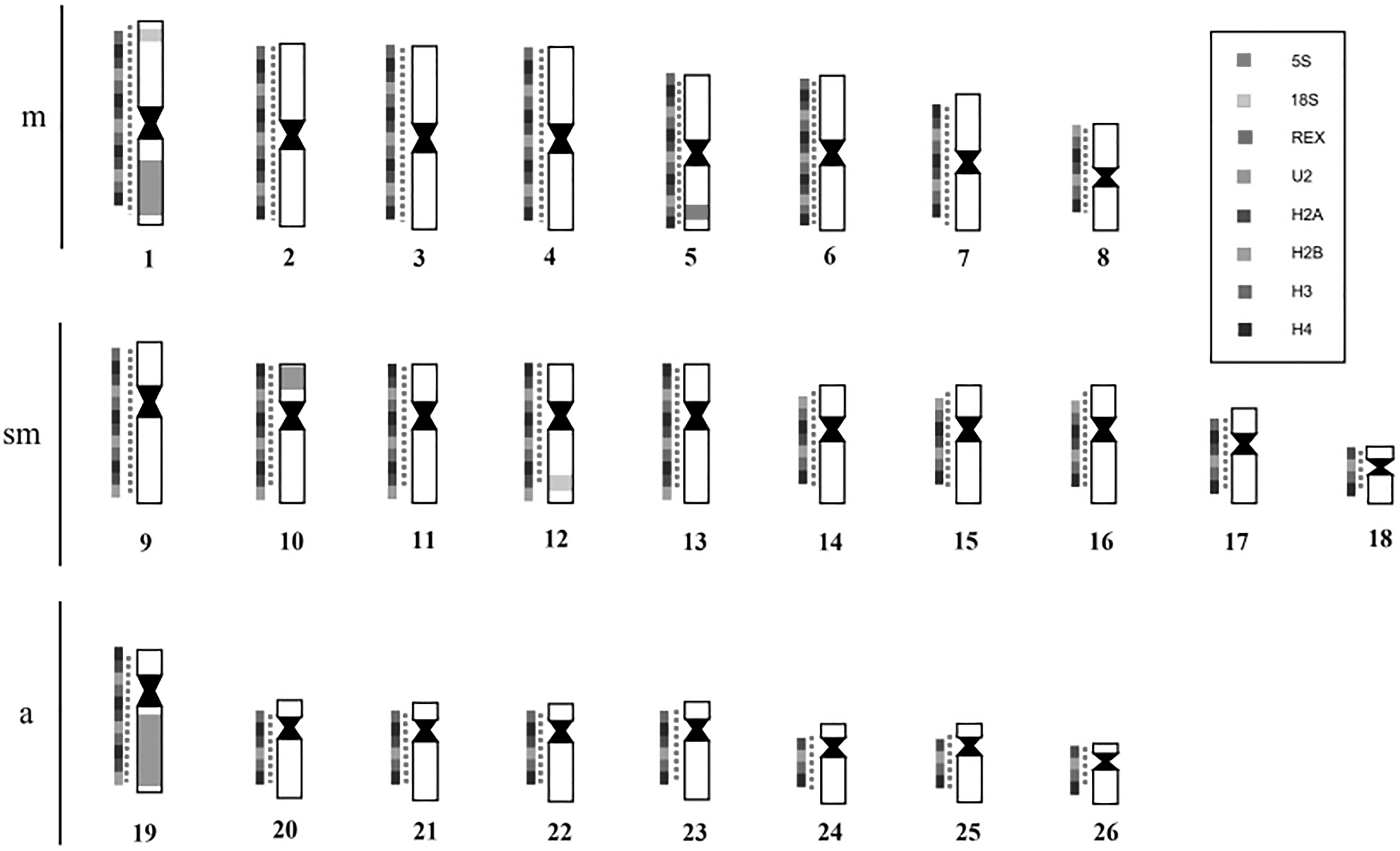
Idiogram showing the mapping of different sequences studied in H. zebra.
The multigene family of histones are considered excellent cytogenetic markers; however, histone gene mapping has been performed only in few organisms. In fish, most works were directed to the order Characiformes, with few studies regarding the organization of these sequences in Loricariidae.11,16,17,23 In most bony fish groups, histone genes are clustered in the karyotype.9,11–13,15–17 Studies show that the location, organization, and number of copies of histone clusters vary between species.4,5 Pendás et al, 9 for example, observed a histone cluster located in only one lócus in karyotype of Atlantic salmon, rainbow trout, and the brown trout, meanwhile, Hashimoto et al 11 observed that in Astyanax species, histone clusters were present in two chromosome pairs.
The histone sequences mapped in this study showed signals dispersed by the karyotype of the species, with variable degree of accumulation and location; H2A genes showed the least accumulation of signals, with small marks distributed along the chromosomes, whereas H2B, H3, and H4 showed small signals accumulated in certain chromosomal regions. These results resemble recent findings such as those observed in the karyotype of Synbranchus marmoratus 18 with H3 and H4 histone genes, as well as species from the genus Characidium and family Paradontidae related to H1 and H3 histone genes.22,23
Recent studies have shown that the action of transposable elements is a major factor in the dispersion of multigene families on the karyotype, and considering the insertion capacity of these elements, the dispersion of histone signals can be attributed as a consequence of the hitchhiking of genes histone, caused by transposition events of the mobile elements.22,23 Such influence was observed in the H3 histone sequence in Rhinoclemmys punctularia, which exhibited high similarity with the retrotransposon Gypsy, and they propose that from evolutionary processes (molecular cooptation), this retroelement has been inserted in the sequence of histone H3 and acquired new genomic functions, and explains that the scattered signals of H3 visualized in the karyotype of R. punctularia are also signals of Gypsy. 44 Although we have not identified the presence of transposable elements in the partial sequences of histone genes in Hypancistrus (analyzed in this study), it is not possible to completely exclude the possibility that the dispersion of histone signals in this genus may be owing to transposition events, because transposable elements may be located in other regions of these multigenes.
In addition to the interference of transposable elements, it is also important to consider other factors that influence the dispersion of repetitive DNA signals throughout the genome, such as the action of circular DNAs, a class of DNA composed of chromosomal sequences, including in tandem-repeated genes such as histones and ribosomal genes, which become abundant after exposure to carcinogens,45,46 or ectopic recombination, an atypical recombination event where the crossing-over process occurs in nonhomologous regions, triggering chromosomal rearrangements.47,48
As observed in this study and in previous studies, the multigene family of histones is quite complex, presenting varied patterns of dispersion in different groups of fish; the presence of clusters in certain chromosomal pairs or small signals dispersed by the chromosomes associated with interference of transposable elements being common. The results obtained in this study show that in the karyotype of Hypancistrus, the histone sequences are dispersed, even so, further studies should be carried out to obtain information about the dynamics of this family of genes in cytogenetically conversed groups such as Hypancistrus.
Footnotes
Acknowledgments
This study is part of Carlos Eduardo's Masters Dissertation, who was a recipient of a CAPES Scholarship in Aquatic Ecology and Fishing (PPGEAP), UFPA. The authors thank Cesar Martins (UNESP-Botucatu) by sequencing analysis, and ICMBio (Chico Mendes Institute for Biodiversity Conservation) for the collection permit (ICMBio Registration: 21078).
Authors' Contributions
C.E.V.S. and R.C.R.N.: Conceptualization; C.E.V.S., B.R.R.A, L.F.S.F., A.L.C., L.M.S., and R.C.R.N.: Methodology, Data curation, Validation, Formal analysis, Investigation; R.C.R.N., C.Y.N., and J.C.P: Funding acquisition, Resources; R.C.R.N.: Project administration, Supervision; C.E.V.S. and R.C.R.N.: Visualization, Writing—original draft; C.E.V.S, R.C.R.N., L.M.S., B.R.R.A, L.F.S.F., A.L.C.,C.Y.N., and J.C.P.: Writing—review and editing. All authors approved the submitted article.
Ethics Statement
The specimens were collected with authorization from the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio), SISBIO no21078. This study was conducted in strict compliance with the ethical recommendations for the use and handling of animals in research, under a protocol approved by the Ethics Committee on Research with Experimental Animals (No. 68–2015). The Cytogenetics Laboratory from UFPA has permit number 19/2003 from the Ministry of Environment for sample transport and permit 52/2003 to use the samples for research.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was financed by the Coordenação de Aperfeiçoamento de Pessoa de Nível Superior (CAPES) to C.Y.N (047/2012) and Banco Nacional do Desenvolvimento Econômico e Social to J.C.P (2,318,697,0001). C.Y.N (307170/2021-7) and J.C.P (307154/2021-1) thank CNPq for productivity grants.