
Abstract
Population aging is a global problem worldwide, and the discovery of antiaging drugs and knowledge of their potential molecular mechanisms are research hotspots in biomedical field. Tetrahydroxystilbene glucoside (TSG) is a natural component isolated from Heshouwu (Polygonum multiflorum Thunb.). It has been widely used to treat various chronic diseases for its remarkable biological activities. In this study, we successfully established aging larval zebrafish by exposing larvae to 2 mM hydrogen peroxide (H2O2). Using this aging model, we assessed the antiaging effect of TSG with different concentrations (25–100 μg/mL). After being treated with H2O2, zebrafish showed the obvious aging-associated phenotypes characterized by higher senescence-associated β-galactosidase activity, significantly downregulated expression of sirtuin 1 (sirt1) and telomerase reverse transcriptase (tert), and upregulated serpine1 mRNA level compared to the control group. TSG pretreatment delayed the aging process of oxidative stress-induced zebrafish, indicative of the reduced positive rate of senescence-associated β-galactosidase, improved swimming velocity, and stimulus-response capacity. Further studies proved that TSG could suppress reactive oxygen species production and enhance the activity of antioxidant enzymes superoxide dismutase and catalase. TSG also inhibited the H2O2-induced expressions of inflammation-related genes il-1β, il-6, cxcl-c1c, and il-8 in aging zebrafish, but it did not affect apoptosis-related genes (bcl-2, bax, and caspase-3) of aging zebrafish. In conclusion, TSG can protect against aging by regulating the antioxidative genes and enzyme activity, as well as inflammation in larval zebrafish, providing insight into the application of TSG for clinical treatment of aging or aging-related diseases.
Introduction
With the aging population growing at an unprecedented rate worldwide, aging is the 21st century's dominant demographic phenomenon. 1 Aging is characterized by a progressive loss of physiological integrity, leading to decreased bodily functions and an increased death rate. Meanwhile, aging is a predominant risk factor for most aging-related diseases, such as cancers, cardiovascular disorders, and neurodegenerative diseases. 2 In recent years, aging research has experienced unprecedented advances that promote an understanding of aging mechanisms.
Studies show that the aging process can be slowed down to a certain extent by intervening in cellular senescence, 3 internal environment, 4 gut microbiota,5,6 and caloric restriction, 7 which will alleviate the functional decline of aging-related diseases. Although evidence has revealed the potential molecular mechanisms that trigger aging, there is still no fully effective drug for preventing aging due to its complex process. Hence, how to delay aging and the screening of antiaging medicines have been a research hotspot in the biomedical field.
Zebrafish shares high homology with the human genome and has emerged as a model system for vertebrate aging research. 8 The telomere length of zebrafish is similar to that of human beings, and they have the same aging process as human beings, but their life span is relatively short.9,10 Like humans, zebrafish are subject to gradual aging, which displays similar senescence-associated symptoms, including high β-galactosidase activity, increased oxidized protein levels, severe spinal curvatures, impaired locomotor activity, and declined cognitive functions.11–13 Therefore, zebrafish is an ideal aging model for large-scale, high-throughput pharmacological and genetic screening.
Polygonum multiflorum Thunb., called Heshouwu in China, is a traditional Chinese herbal medicine and exhibits curative activities in treating various diseases,14,15 such as cancer, 16 dyslipidemia, 17 and aging-related diseases.18,19 Stilbene glycoside (2, 3, 5, 4′-tetrahydroxystilbence-2-O-β-D-glucoside, TSG) is a natural component extracted from Polygonum multiflorum Thunb. TSG is the main quality control standard of Polygonum multiflorum Thunb in the Chinese pharmacopoeia. 20 Modern pharmacological researches show that TSG has various biological activities, including anti-inflammation, antioxidation, neuroprotection, and antiaging.21,22
Based on previous studies, accumulation of reactive oxygen species (ROS) can lead to oxidative stress and consequent damage to cellular membranes and functions, thus leading to cellular senescence.23,24 Hence, TSG may be conducive to delaying the oxidative stress-induced aging process. However, there are limited investigations to clarify the underlying pharmacological mechanisms of TSG against aging. This study established an oxidative stress-induced aging zebrafish model by treating larvae with hydrogen peroxide (H2O2). Using this model, we explored the antiaging effect and potential mechanism of TSG.
Materials and Methods
Materials
2,3,5,4′-Tetrahydroxystilbence-2-O-β-D-glucopyranoside (TSG) (purity >98%) was purchased from Aladdin reagent, Inc. (Shanghai, China). A H2O2 (30 wt.%) solution was purchased from Sinopharm Chemical Reagent Co., Ltd. (Beijing, China). Senescence-associated β-galactosidase (SA-β-Gal) Staining Kit was purchased from Cell Signaling Technology, Inc. (Beverly, MA). ROS Assay Kit, Total Superoxide Dismutase (SOD) Assay Kit, Catalase (CAT) Assay Kit, and Lipid Peroxidation malondialdehyde (MDA) Assay Kit were obtained from Beyotime Biotechnology (Shanghai, China). Trizol reagent, cDNA synthesis kit, and SYBR QPCR Mixture were purchased from ALLMEEK Technology Co., Ltd (Beijing, China).
Zebrafish feeding and TSG treatment
The wild-type AB strain zebrafish used in this study were raised in recirculation systems at 28.5°C under an alternative 14-h light/10-h dark cycle. H2O2 and TSG reagents were directly dissolved into egg water. Larval zebrafish were randomly divided into five groups (n = 100 per group) at 72 h postfertilization (hpf) and cultured in the 90 mm cell culture dish. The specific operations of each group are as follows: control group, treated with egg water; H2O2 group, treated with H2O2 (2 mM) in embryo water; TSG-L group, treated with H2O2 plus low concentration of TSG (25 μg/mL); TSG-M group, treated with H2O2 and middle concentration of TSG (50 μg/mL); and TSG-H group, treated with H2O2 and high concentration of TSG (100 μg/mL).
Varying concentrations of TSG (25–100 μg/mL) was dissolved into egg water and pretreated larvae for 24 h. Then, 96 hpf larvae were incubated with egg water containing 2 mM H2O2 for 4 h, as referred by a previous report. 25 During the experiment, a complete water change was carried out every 24 h to ensure water quality. The flow diagram of the experimental protocol is shown in Figure 1. All animal experiments were approved by the animal ethics committee of Hubei University of Chinese Medicine and conducted following the Guiding Principles for the Care and Use of Laboratory Animals.

Outline of experimental design used in this study. hpf, hours postfertilization.
SA-β-gal staining
According to the manufacturer's instructions (Cell Signaling Technology, Inc., MA), SA-β-gal activity was detected in larval zebrafish. Briefly, larval zebrafish were fixed in a fixative solution at 4°C overnight and then rinsed three times for 15 min in phosphate-buffered saline (PBS) (pH = 7.4), followed by washing in PBS (pH 6.0) for another 1 h at 4°C. After that, 1 mL of staining solution was added, and the samples were incubated overnight at 37°C in a dry incubator (without CO2). Images were taken using a dissecting microscope with an MSX1 Microscope camera (Mshot, China). Qualitative analysis was done by a selection tool in Adobe Photoshop for the area positive of SA-β-gal staining according to reference. 26
Behavior analysis
The larval behavioral testing was performed using the DanioVision system (Noldus, Wageningen, Netherlands). Zebrafish larvae from each experimental group (n = 18) were placed individually in 96-well plates with 150 μL of embryo water and allowed to acclimatize to the environment for 10 min. During the locomotor analysis, free-swimming speed was recorded for a 10-min light (visible light) period, followed by a 10-min dark (infrared light) period. The stress responses of individuals to light stimulus were tested by altering light on/off for flash, as well as the swimming behavior in response to vibration stimulus. Behavioral parameters were recorded and analyzed using the EthoVision software (Noldus, Wageningen, Netherland).
Measurement of ROS
ROS production in zebrafish embryos was analyzed using an oxidation-sensitive fluorescent probe dye, that is, 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) (Beyotime Biotechnology, Shanghai, China). Briefly, zebrafish were pretreated with different concentrations of TSG for 24 h and then exposed to 2 mM H2O2 for 4 h. After treatment, zebrafish embryos were further incubated with 10 μM DCFH-DA at 28.5°C in the dark for 30 min. After three rinses in embryo media, the stained embryos were viewed and photographed using a fluorescent microscope (Leica, Wetzlar, Germany). The fluorescence intensity was quantified using a fluorescence microplate reader (Molecular Devices Corporation, CA) for the ROS production among experimental groups.
Analysis of oxidative stress-related enzymes and peroxide
To evaluate the antioxidative activity of TSG in oxidative stress-induced aging zebrafish, we measured the activities of SOD and CAT, and the level of MDA. The experiment was performed according to the manufacturer's instructions (Beyotime Biotechnology, Shanghai, China).
Real-time quantitative polymerase chain reaction analysis
The total RNA of zebrafish embryos was extracted using Trizol reagent (Summerbio, Beijing, China) and reverse transcribed using the first-strand cDNA synthesis kit (ALLMEEK Technology Co., Ltd, Beijing, China). Real-time quantitative polymerase chain reaction (RT-qPCR) was performed with SYBR QPCR Mixture (ALLMEEK Technology Co., Ltd, Beijing, China) on a Bio-RAD CFX Connect Real-time System (BioRad, CA). The RT-qPCR protocol was as follows: 30 s at 95°C for 30 s, 40 cycles at 95°C for 10 s, 60°C for 30 s, and 72°C for 30 s. The specific sequences of primers used in this study are shown in Table 1. The 2−ΔΔCt method was used to calculate the relative expression levels of target genes by normalizing them to that of β-actin.
Specific Sequences of Primers Used in Real-Time Quantitative Polymerase Chain Reaction
Statistical analysis
The graphs were drawn using Prism 5 GraphPad Software. All data were presented as mean ± standard error of the mean. Differences among experimental groups were evaluated by a two-way analysis of variance (ANOVA) with the Kruskal–Wallis post hoc test. A p-value <0.05 was considered statistical significance. Results were confirmed in three independent experiments.
Results
Protective effect of TSG on oxidative stress-induced premature aging in larval zebrafish
After pretreatment with different concentrations of TSG for 24 h, larval zebrafish were induced aging with the H2O2 solution (2 mM) for 4 h. Then, we collected the zebrafish samples from each group to analyze the age-related indicators. The most widely used biomarker for aging is senescence-associated β-galactosidase (SA-β-gal), with a high galactosidase activity detectable at pH 6.0 in senescent cells or tissues. Larval zebrafish in the H2O2 group showed significantly higher SA-β-gal activity than the control group, especially in the head and abdomen (Fig. 2A, B), indicating that H2O2 treatment increased the number of senescent cells. Concomitantly, the mRNA levels of aging-related genes, like sirtuin 1 (sirt1) and telomerase reverse transcriptase (tert), were decreased in the H2O2 group compared to the control group (p < 0.05 or 0.01) (Fig. 2C, D).

TSG pretreatment protected H2O2-induced aging in larval zebrafish.
As an inducer of cell senescence, the mRNA expression of serpine1 was upregulated in the H2O2 group (p < 0.05 vs. control group) (Fig. 2E). In contrast, TSG pretreatment (25–100 μg/mL) reduced evidently the number of senescent cells in oxidative stress-induced larvae zebrafish (Fig. 2A, B). Meanwhile, TSG statistically increased the expression of sirt1, but decreased serpine1 mRNA level in oxidative stress-induced larval zebrafish (p < 0.05 or 0.01 vs. H2O2 group) (Fig. 2C, E). TSG pretreatment had no significant effect on tert mRNA level.
Restoration of locomotor behavior and stimulus-response capacity in oxidative stress-induced aging larval zebrafish pretreated with TSG
Given that aging usually accompanies impaired swimming performance and reaction capacity in zebrafish,11,27 we performed a light-dark locomotion test in which larval zebrafish were monitored for 20 min by light and vibration stimulus. Movement heatmaps showed that larval zebrafish had decreased swimming and motor impairments when exposed to H2O2 (Fig. 3A).
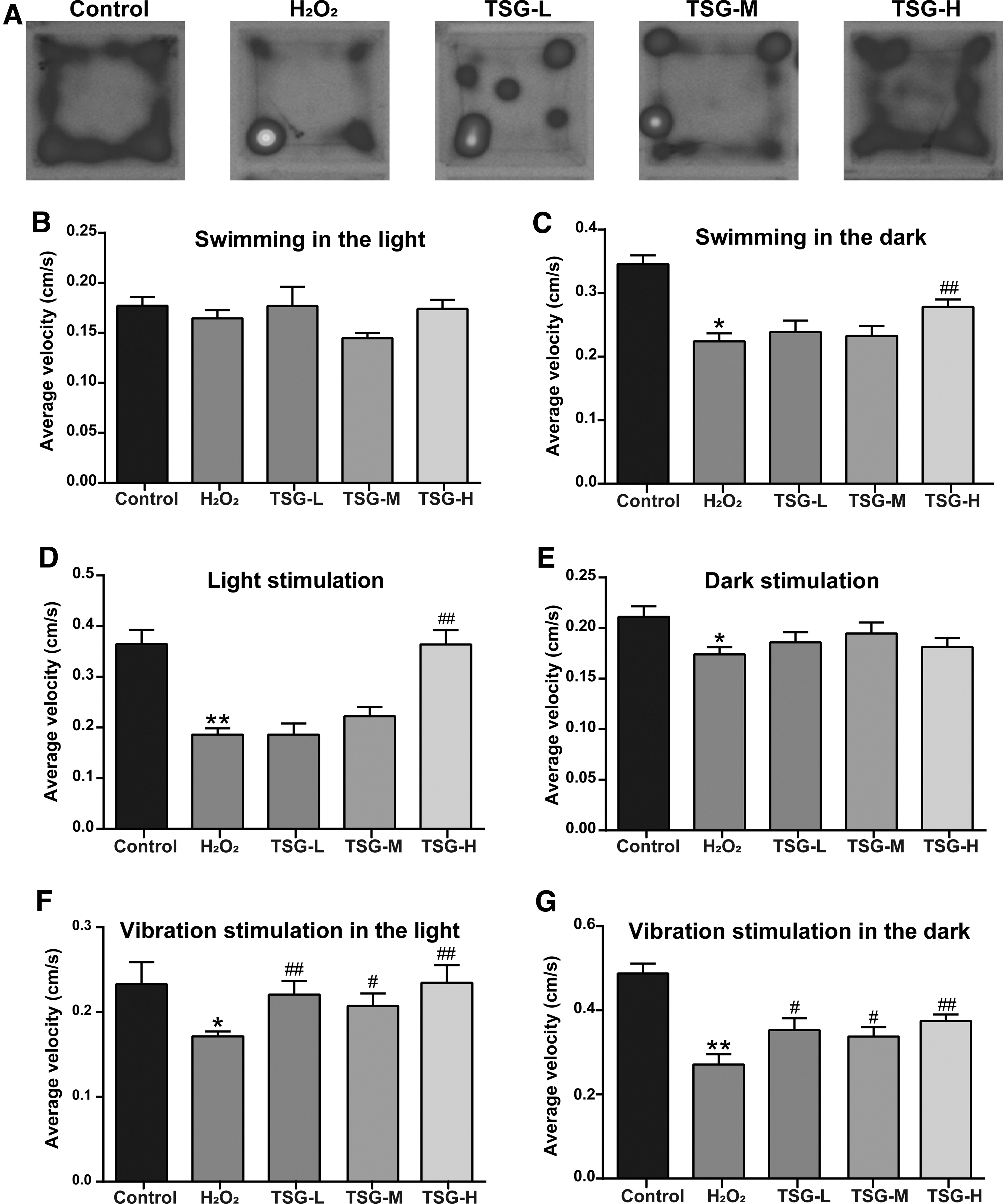
Effect of TSG on locomotor behavior of larval zebrafish induced by H2O2.
However, the larval zebrafish in each group exhibited normal activity as in the control group during the light period (Fig. 3B). In the dark period, the swimming velocity was significantly lower in the H2O2 group (0.22 cm/s) compared with the control group (0.35 cm/s) (p < 0.05 vs. control group), and it increased to 0.28 cm/s in the TSG-H group (p < 0.01 vs. H2O2 group) (Fig. 3C). Notably, when light-on or light-off for stimulus, H2O2 exposure statistically decreased the swimming speed by 50% and 22% in larval zebrafish, respectively (p < 0.05 or 0.01 vs. control group) (Fig. 3D, E).
When a vibration stimulus presented in the observation period, H2O2 exposure statistically decreased the swimming velocity by 27% in the light and 48% in the dark, respectively (Fig. 3F, G). TSG pretreatment recovered the above behavioral parameter changes in oxidative stress-induced larval zebrafish, especially in the TSG-H group. Compared with the H2O2 group, TSG pretreatment at 100 μg/mL increased significantly the moving speed by 49% upon light-on flash stimulus (p < 0.01 vs. H2O2 group) (Fig. 3D). When vibration stimulus presented in the TSG-H group, TSG pretreatment significantly increased the moving speed by 26% in the light and 32% in the dark (p < 0.01) (Fig. 3F, G), suggesting the ameliorative effect of TSG on locomotion defects in oxidative stress-induced aging zebrafish.
Inhibitory effect of TSG on oxidative stress-induced ROS generation in aging larval zebrafish
High levels of ROS are associated with the cellular damage and aging process of organisms. 24 Hence, we analyzed the ROS level of larval zebrafish in each group using an oxidation-sensitive fluorescent probe, DCFH-DA. Compared with the control group, the ROS level of larval zebrafish in the H2O2 group was markedly increased in liver and intestine tissues (Fig. 4A). However, TSG pretreatment (25–100 μg/mL) showed a significant decrease of ROS in oxidative stress-induced larval zebrafish (Fig. 4A). Quantitation in fluorescence imaging also showed that the level of ROS statistically elevated in the H2O2 group (p < 0.01 vs. control group), and TSG pretreatment dose-dependently reduced the intensity of ROS fluorescence in oxidative stress-induced zebrafish larvae (p < 0.01 vs. H2O2 group) (Fig. 4B).
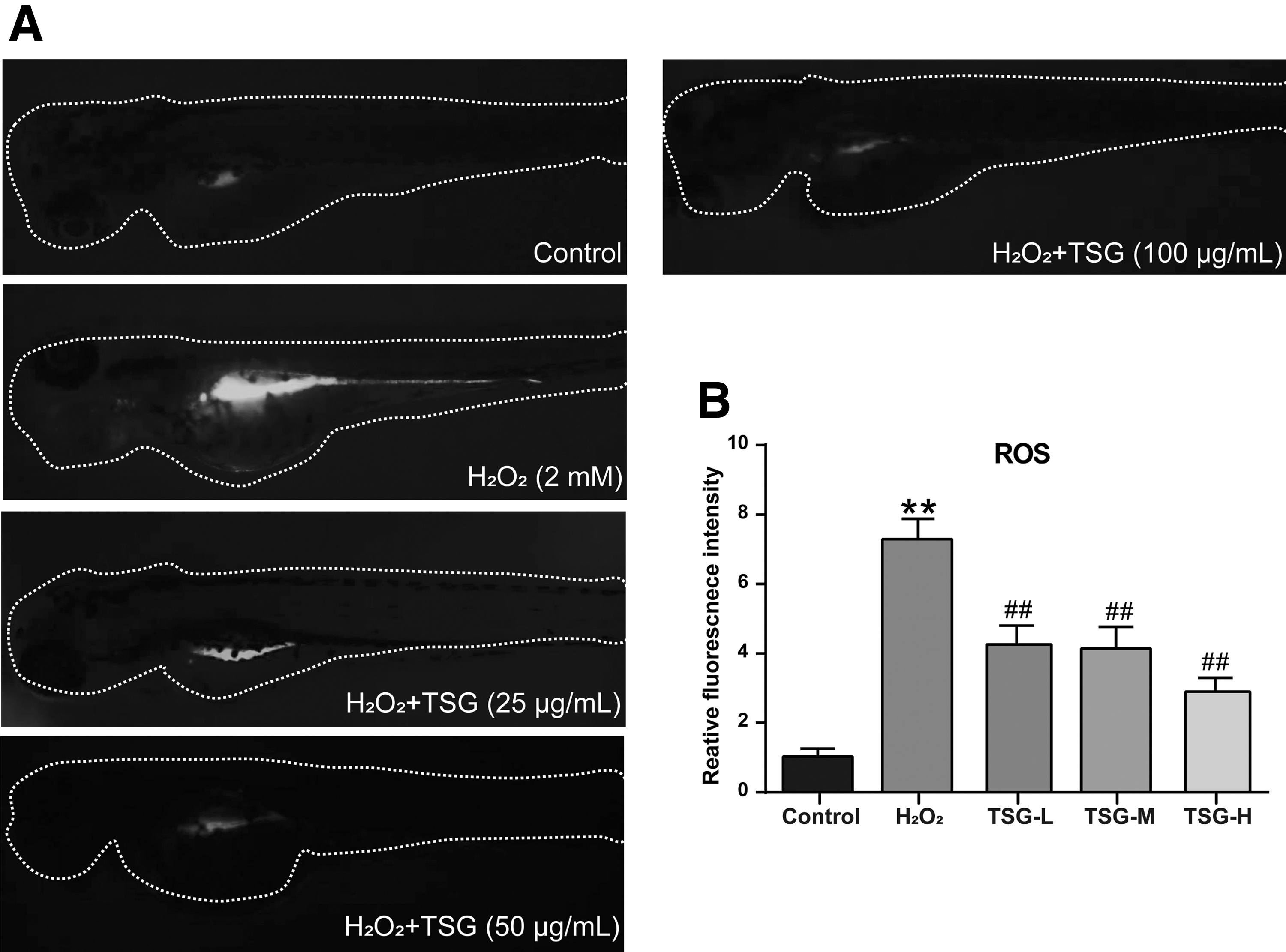
TSG pretreatment suppressed H2O2-induced ROS generation in larval zebrafish.
Antiaging effect of TSG by alleviating oxidative damage in oxidative stress-induced larval zebrafish
Aging is partly triggered by the imbalance between ROS production and antioxidative defense in organisms.23,28 SOD and CAT represent two main antioxidant enzymes. MDA is a lipid peroxidation product and a sensitive marker of oxidative stress. To confirm the antioxidative effect of TSG, we tested the activities of antioxidant enzymes in oxidative stress-induced aging zebrafish with or without TSG treatment.
As shown in Figure 5A and B, the remarkable decreases of SOD and CAT activities were detected in the H2O2 group (p < 0.05 or 0.01 vs. control group). Meanwhile, H2O2 induced a significant increment in MDA content (p < 0.05 vs. control group) (Fig. 5C). Although TSG treatment had no effect on the MDA level of the H2O2 group, it elevated the activities of both SOD and CAT in oxidative stress-induced larval zebrafish (p < 0.05 or 0.01) (Fig. 5A, B). Furthermore, the RT-qPCR results suggested that the expressions of sod1 and nqo1 were downregulated in the H2O2 group (p < 0.05 vs. control group), but were recovered after TSG pretreatment (p < 0.05 or 0.01 vs. H2O2 group).

TSG inhibited oxidative stress caused by H2O2 in aging larval zebrafish.
Inhibitory effect of TSG on inflammatory responses in oxidative stress-induced aging larval zebrafish
Aging usually accompanies a physical state of chronic inflammation. 29 To further investigate the underlying mechanism of TSG against aging, we examined the mRNA expressions of inflammation-related genes in aging larval zebrafish. H2O2 treatment significantly facilitated mRNA expressions of pro-inflammatory genes, including interleukin-1 beta (il-1β), interleukin 6 (il-6), chemokine (C-X-C motif) ligand C1c (cxcl-c1c), and interleukin 8 (il-8) (p < 0.01 vs. control group). Besides, H2O2 treatment also reduced mRNA level of transforming growth factor-beta (Tgf-β, an anti-inflammatory cytokine) (p < 0.05 vs. control group) (Fig. 6). Noticeably, TSG pretreatment reversed all the above changes in oxidative stress-induced aging larvae (Fig. 6).

TSG suppressed inflammatory responses in H2O2-induced aging larval zebrafish. The mRNA levels of inflammation-related genes in larval zebrafish were detected by RT-qPCR, including il-1β
Cell senescence triggered by oxidative stress usually leads to cellular damage and subsequent cell apoptosis.30,31 Hence, we also detected the mRNA expression of apoptosis-related markers (bcl-2, bax, and caspase-3). Results displayed that the mRNA levels of three genes had no dramatic alteration in the H2O2 group compared to the control group (Fig. 7A, B, D). Although the bcl-2/bax ratio was significantly decreased in oxidative stress-induced larval zebrafish (Fig. 7C) (p < 0.05 vs. control group), TSG did not affect apoptosis-related genes of aging zebrafish.

Effect of TSG on cell apoptosis in H2O2-induced aging larval zebrafish.
Discussion
With the aggravation of population aging, how to delay aging has become a research hotspot, and selecting appropriate animal models is vital for exploring the mechanism of aging and developing antiaging drug candidates. 32 As a powerful vertebrate model for developmental and genetic studies, zebrafish has many similar aging characteristics to mammals for aging research.25,33 Zhao et al. showed that depletion of Δ133p53 (a human p53 isoform) resulted in the long-term stress of ROS at a low level and finally led to accelerated brain aging in zebrafish. 34 Sirt1-knockout zebrafish attracted a significantly increased generation of ROS that was subsequently followed by apoptosis and aging. 35
In this study, we established oxidative damage-induced aging zebrafish by exposure of 96 hpf zebrafish larvae to H2O2. The aging zebrafish had more senescent cells, upregulated aging-promotive genes, and decreased antiaging genes (Fig. 2). Moreover, H2O2-treated larval zebrafish displayed a reduced swimming performance and slowed escape responses to external stimuli (Fig. 3), while TSG pretreatment effectively ameliorated these pathological symptoms. Thus, H2O2 exposure to larval zebrafish can establish the oxidative stress-induced aging model, while TSG pretreatment could alleviate the aging-related signals in aging zebrafish.
Cellular ROS are generated endogenously during the process of oxygen metabolism in cells, like superoxide anion (O2•−), hydroxyl radical (OH•), and H2O2. 24 ROS may derive from interactions with exogenous sources, and their double-edged role in organisms is complicated. Low levels of ROS are necessary for activating the signaling pathways to initiate biological processes. On the contrary, higher levels of ROS and their long-time exposure will incur damage to cellular biomacromolecules such as DNA, lipids, and proteins. The accumulation of oxidative damage contributes to irreversible cell injury that causes ultimate cell death and aging onset.23,36,37 In contrast, antioxidants can neutralize ROS and attenuate oxidative damage, thereby inhibiting aging process and preventing aging-related diseases.
Previously, the in vitro and in vivo results revealed that TSG had an antioxidant effect and hydroxyl radical-scavenging activity.22,38,39 For instance, TSG can protect human brain microvascular endothelial cells from oxidative stress-induced dysfunction by inhibiting oxidative stress and inflammatory responses. 38 Also, TSG prevented glial cells (PC12 cells) from 6-hydroxydopamine-induced apoptosis by regulating the ROS-NO pathway. 40 Furthermore, TSG could alleviate oxidative stress-induced apoptosis and autophagy in mouse cochlear UB/OC-2 cells through the Nrf2 antioxidant pathway. 41 In vivo experiments proved that TSG enhanced stress resistance to extend the life span of Caenorhabditis elegans. 42
Likewise, TSG acted as a potent antioxidant against D-galactose-induced aging in rats by increasing SOD activities and glutathione peroxidase. 43 These results imply the potential of TSG as a pharmaceutical antiaging drug. In this study, we further demonstrated that TSG exerted antioxidant activities in aging process by removing excess ROS and upregulating the mRNA expressions of antioxidant enzymes (SOD and CAT) and antioxidative genes (Sod1 and Nqo1) (Figs. 4 and 5). In summary, we confirmed that TSG had an antioxidative effect on aging zebrafish by strengthening oxidative resistance.
Chronic low-grade systemic inflammation is associated with the degenerative aging process and represents a significant risk factor for morbidity and mortality in the elderly. 44 Senescence-associated secretory phenotype (SASP) is a proinflammatory response linked to aging. 45 In aging mice and older adults, SASP factors are statistically increased, such as IL-1β, IL-6, IL-8, and tumor necrosis factor-α.46,47 In previous studies, TSG has been proven effective in proinflammatory models. For example, TSG could increase mRNA levels of proinflammatory cytokines and activate the expression of Sirt1 to ameliorate ligature-induced periodontitis in rats. 48 TSG dramatically attenuated acetic acid-induced colon lesions by upregulating PPAR-γ and inhibiting NF-κB inflammatory pathway in mice. 49
In addition, Fan et al. demonstrated that TSG reduced the release of proinflammatory cytokines and blocked the p38/NF-κB signaling pathway to relieve Freund's adjuvant-induced chronic inflammatory pain in mice. 50 In parallel, our studies found that the antiaging effect of TSG contributed to its anti-inflammatory activities by regulating inflammation-associated gene expressions in aging zebrafish, including il-1β, il-6, cxcl-c1c, il-8, and Tgf-β (Fig. 6). Based on the above, TSG has an antiaging effect by suppressing the inflammatory responses in larval zebrafish.
Remarkably, TSG plays a neuroprotective role in Parkinson's disease by regulating Bax/Bcl-2 ratio and caspase-3 activity to inhibit apoptosis in MPP+-exposed cells.51,52 However, we did not find the inhibitory effect of TSG on apoptosis in oxidative stress-induced aging zebrafish, partly due to the distinction between in vitro and in vivo studies or different disease models. It is also possible that the degree of apoptosis is relatively mild for oxidative stress-induced larval zebrafish since no significant difference in the caspase-3 activity occurred between the control and H2O2 groups (Fig. 7).
In conclusion, this study reveals that the aging model is established successfully by treating zebrafish with H2O2. Using this model, we demonstrated that TSG delayed aging process in oxidative stress-induced larval zebrafish. Furthermore, we explored the underlying mechanism of the TSG-initiated antiaging effect, indicative of the suppressed ROS production, enhanced expressions of antioxidant enzymes, and inhibited inflammatory responses in aging larval zebrafish. Summarily, our studies provide insight into the application of TSG for clinical treatment of aging or aging-related diseases.
Footnotes
Authors' Contributions
All authors contributed to the study conception and design. H.L. and X.Z. conceived and designed research. H.X. and X.C. conducted experiments and wrote the article. X.Y. and X.S. contributed new reagents or analytical tools. F.H. and M.C. analyzed data.
Ethical Approval
All animal experiments were approved by the animal ethics committee of Hubei University of Chinese Medicine and conducted following the Guiding Principles for the Care and Use of Laboratory Animals.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Hubei Provincial Department of Education (no. Q20192006).