
Abstract
Un-ionized ammonia (NH3) is a prevalent contaminant found in aquatic ecosystems, frequently associated with complex mixtures of other toxic contaminants. Early embryo-larval stages of zebrafish Danio rerio became an important model for water quality monitoring, and it is necessary to characterize its susceptibility to NH3 exposure. Fertilized eggs were exposed to NH3 concentrations ranging from 0.02 to 5.23 mg NH3 L−1 until 168 h postfertilization (hpf). The lethal concentration to 50% of exposed zebrafish during 96 h was 2.07 mg NH3 L−1, 25% above the median value reported values for early developmental stages of fishes. Sublethal toxicity endpoints indicated the lowest observed effect concentrations for slow blood circulation at 24 hpf, decreased heart ventricular contractions at 72 hpf, incomplete yolk sac absorption at 96 hpf, failure in swim bladder inflation at 96 hpf, developmental retardation at 96 hpf, decreased total length, decreased swimming speed, altered trajectories, and acetylcholinesterase inhibition at 168 hpf of 0.85, 0.06, 0.15, 0.06, 0.15, 0.61, 1.35, 0.35, and 0.85 mg NH3 L−1, respectively. Environmentally relevant NH3 concentrations can affect zebrafish's early development and larval viability, and our results help discriminate NH3 contribution to the toxicity of complex environmental mixtures when zebrafish is used in water quality monitoring.
Introduction
Ammonia is an important contaminant in aquatic ecosystems due to anthropogenic alteration of the nitrogen cycle. Inorganic nitrogen can enter aquatic ecosystems through point sources such as wastewaters from livestock and municipal sewage effluents or nonpoint sources such as runoff from agricultural fertilizers or urban runoff from unsewered urban areas. 1 Ammoniacal nitrogen is found in freshwaters as NH4+ ion or the more potent un-ionized NH3, which is ecotoxicologically relevant and harmful to fishes, which are among the most sensitive groups with regard to both chronic and acute effects of ammonia. 2 The species mean acute value for 44 fishes range from 0.21 to 1.11 mg NH3 L−1, 2 and the lethal NH3 concentration to 50% of individuals exposed during 48 to 96 h varies from 0.14 to 4.2 mg NH3 L−1 in Canadian freshwater fishes. 3
Sublethal consequences of ammonia retention in fishes are linked to neurotoxicity caused by elevated NH4+ displacing K+ and depolarizing neurons leading to cell death in the central nervous system, 4 reduced immune defenses, 5 imbalance of ionic regulation, impairment of swimming performance, reduced feeding and slower growth, 6 and reduced ability to escape from predators. 7 Juvenile and adult teleostean fishes are mostly ammoniotelic, excreting ammonia to the surrounding water without energy expenditure. 8 During early life stages fish embryos use yolk proteins as their major energy source, generating ammonia which is not easily excretable through the chorion before they hatch, and several species are ureotelic to prevent ammonia toxicity during this embryonic phase, becoming ammoniotelic only after hatch. 9 Hatched larvae are considered the most sensitive stages, and the lethal NH3 concentration to 50% of individuals exposed during 96 h varies from 0.39 mg NH3 L−1 in sensitive Sciaenops ocellatus 10 to 11.5 mg NH3 L−1 in more resistant larval stages of Labeo bata. 11
The use of parameters focused on embryo-larval development of the zebrafish Danio rerio to monitor the ecotoxicity of surface waters in rivers12–15 and lakes 16 has grown and it has been proposed as an important effect based tool for monitoring and assessment within the European Water Framework Directive. 17 Concentrations of NH3 verified in these aquatic ecosystems vary from less than 0.1 to more than 4 mg NH3 L−1 and are frequently present in association with a diversity of inorganic and organic contaminants in complex environmental mixtures. Although the toxicity of nitrate 18 and nitrite 19 to zebrafish embryo-larval stages has been studied, there is a lack of detailed studies focused on lethal and sublethal effects of NH3 within environmentally relevant concentrations.
Therefore, the objective of this work is to establish the sensitivity of the initial embryo-larval stages of Danio rerio to a range of environmentally relevant NH3 concentrations in terms of lethal and sublethal endpoints focused on detailed aspects of embryo-larval development, developmental abnormalities, and biochemical, behavioral, and morphometric effects. This information will also help in the discrimination of NH3 contribution to the toxicity of surface water mixtures containing ammonia when the abovementioned zebrafish endpoints are used in water quality monitoring.
Materials and Methods
Cultivation of Danio rerio
Wild-type short-finned zebrafish originally purchased from a local supplier was kept in 100 L recirculating aquariums with mechanical and biological filtration. Water quality parameters in breeding aquariums were analyzed with a YSI Professional Plus multiparameter probe, and pH was 7.8 ± 0.4 (mean ± standard deviation), dissolved oxygen was 6.0 ± 0.4 mg L−1, temperature 27°C ± 1°C. Breeding adult Danio rerio was fed Artemia nauplii and commercial fish food with 40% protein. Fertilized eggs were obtained from a group of 3 males and 6 females separated the previous afternoon in a 20 L aquarium until early morning spawning and fertilization. The selection of fertilized and viable eggs was made by direct observation in a Stemi 2000 C stereomicroscope (Zeiss Microscopy, Rio de Janeiro, Brazil) at 50 × magnification. The fertilization rate was above 90%, and animal handling and embryo exposures were performed by a protocol approved by the Ethics Committee for Animal Experiments from the Federal University of Pernambuco (UFPE).
Ammonia concentration analysis
Total ammonia concentrations (NH4+ + NH3) in water were quantified by the colorimetric indophenol method
20
adapted to 96-well microplates. A calibration curve was prepared with nine concentrations of total ammonia prepared by dilutions of an ammonium chloride (NH4Cl) 99.5% pure (Sigma-Aldrich, Sao Paulo, Brazil) stock solution at 0, 0.25, 0.5, 1.0, 2.0, 3.0, 4.0, 5.0, and 6.0 mg NH4 L−1. Absorbance was measured at 673 nm in 96-well microplates in a spectrophotometer SpectraMax M3 (Molecular Devices), and the following analytical curve was obtained relating total ammonia concentrations and absorbance:
Preparation of ammonia exposure solutions
Ammonium chloride (NH4Cl) 99.5% pure (Sigma-Aldrich) was dissolved in ultrapure water to prepare a stock solution at a concentration of 400,000 μg NH4 L−1. The stock solution was appropriately diluted with micropipettes in clean UV sterilized water, pH 8.41 ± 0.02 (mean ± standard deviation), dissolved oxygen 6 ± 0.5 mg L−1, temperature 27.5°C ± 0.5°C, alkalinity expressed as total carbonates 97 ± 5 mg L−1, to obtain the desired total ammonia nominal exposure concentrations.
NH3 concentration in exposure solutions was calculated based on total ammonia concentrations, temperature (27.5°C), pK, and pH (8.41), according to equations from
21
:
pK = 0.09018 + (2729.92/(273.2 + T)) fraction of NH3 = 1/1 + (10(pK-pH)) concentration of NH3 (mg L−1) = fraction of NH3 × total ammonia (mg L−1) where T is temperature in degrees Celsius
Experimental design
Zebrafish embryos at 3 h postfertilization (hpf) were individually exposed in 24-well plates with a water volume of 2.5 mL during 96 h to NH3 concentrations ranging from 0.02 to 5.2 mg NH3 L−1 plus controls, all at a pH of 8.41. Two plates were used as controls with a total of 24 embryos or 24 hatched larvae in each one. Exposure solutions were renewed at a rate of 80% every day in each well. The rate of decay of NH3 concentration in the different plates was measured every 24 h before changing the solutions and after the change (Supplementary Table S1). Mortality was checked according to OECD guidelines, 22 and mortality rates were calculated by the ratio between the total accumulated number of deaths after each considered exposure period and the total of exposed individuals for each sample. The number of embryos at the beginning of exposure for each treatment was 72, 20, 40, 20, 20, 20, 20, 20, 20, 40, 20, 20, and 20 for treatments 0 (control), 0.02, 0.06, 0.15, 0.33, 0.60, 0.85, 1.35, 1.58, 2.25, 3.42, 4.42, and 5.23 mg L−1 NH3, respectively.
General morphology score
General Morphology Score (GMS) integrates partial scores credited to each embryo as they develop hallmark structures during the first 96 h of development. Each embryo of each treatment was individually evaluated for the presence of these characteristics in an inverted DIGILAB® microscope at magnifications ranging from 40 × to 100 × . Both controls and treated embryos receive points for each developmental hallmark achieved, including detachment of the tail during early somite formation; eye development and pigmentation; presence of movement of the embryo or larva; presence of blood circulation; presence of heartbeat; pigmentation of the head and body; pigmentation of the tail; presence of pectoral fins; hatching; presence of a protruding mouth; and consumption of yolk sac reserves. At the end of 96 h of exposure, a perfect larva has a maximum GMS of 17, according to reference. 23 In this study the morphological milestone of swim bladder inflation was also evaluated at 96 hpf, and therefore, the maximum GMS for a perfect larva was equal to 18. Reductions in GMS indicate delays in embryo-larval development.
Developmental abnormalities
Developmental abnormalities were recorded daily for each embryo during testing, and frequencies were calculated dividing the total number of live larvae presenting the abnormalities by the total number of live larvae present in the sample. Evaluated abnormalities from 24 to 96 hpf were: slow or absent blood circulation (blood stasis) (24 hpf) on the posterior cardinal vein and dorsal aorta close to the yolk sac, abnormal spinal curvature (72 hpf), absence of pectoral fin (72 hpf), incomplete protrusible mouth development (72–96 hpf), pericardial edema (96 hpf), yolk sac not completely absorbed (96 hpf), and uninflated swim bladder (96 hpf).
Frequency of ventricular contractions
Heart rate was measured in embryos at 48 hpf and 72 hpf. The observations were made with an inverted microscope at 200 × . Ventricular contractions were counted at 48 and 72 hpf in 10 embryos per exposure concentration over a period of 30 s using a stopwatch.
Swimming endpoints
Surviving larvae at stage 168 hpf were transferred individually to a well from 12-well cell culture plates filled with 3 mL of water. Larvae from the different NH3 treatments were filmed for 300 s in top view using video cameras with 6–60 mm variable zoom lenses, and the videos were recorded on a computer. The mean spontaneous swimming speed (cm s−1) performed by the larvae during the total period of 300 s, maximal swimming speeds of larvae in 30 sequential intervals of 10 s along the monitored time, and swimming trajectory maps were drawn by the video tracking software Spontaneous Motor Activity Recording & Tracking (SMART)(Harvard Apparatus, Holliston). Trajectory maps were used to count the number of larvae that swam along the edges of the well and did not explore the center of the arena (thigmotaxis) and the frequency of larvae showing this behavior at each exposure treatment.
Morphometric measurement
After the behavioral tests larvae were photographed with a digital camera attached to a Stemi 2000 C stereomicroscope (Zeiss Microscopy, Rio de Janeiro, Brazil) at 20 × magnification. Total larval length was measured to an accuracy of 0.1 mm with AxioVision software (Zeiss).
Acetylcholinesterase
The quantification of acetylcholinesterase (AChE) activity was adapted for microplates using the method of Ref. 24 Four sample replicates were analyzed, each one formed by pools of 4 larvae at 96 hpf homogenized in 100 μL of potassium phosphate buffer (0.25 M KPI at pH 7.4) cooled and kept under ice, followed by centrifugation at 3000 g for 4 min in a refrigerated centrifuge (model 5415R; Eppendorf) at 4°C, and the supernatant was used for analysis. During the essay, formation of 5-thio-2-nitrobenzoic acid (TNB) was quantified by absorbance in a spectrophotometer (model SpectraMax M3; Molecular Devices) using a wavelength of 415 nm. A molar absorptivity coefficient Ɛ = 14.15 mM−1 cm−1 was used, and AChE was expressed in μmol TNB min−1 mg protein−1.
Glutathione-S-transferase
Glutathione-S-transferase (GST) activity was adapted for microplates using the method of Ref. 25 Replicates and homogenization of samples used the same procedures for AChE. GST activity was determined from the conjugation of glutathione (GSH) with the substrate 1-chloro-2,4-dinitrobenzene (CDNB), producing a compound that can be detected at 340 nm in a spectrophotometer SpectraMax M3. A molar absorptivity coefficient Ɛ = 9.6 mM−1 cm−1 was used, and GST was expressed in μmol CDNB min−1 mg protein−1.
Catalase
Catalase (CAT) activity was based on consumption of hydrogen peroxide (H2O2) following the method adapted for microplates by Ref. 26 Four sample replicates were analyzed, each one formed by pools of 20 larvae at 96 hpf which were homogenized in 100 μL of buffer and centrifuged at 12,000 g for 30 min at 4°C, and the supernatant was used for analysis. The reaction was quantified by reading the absorbance at 240 nm. A molar absorptivity coefficient Ɛ = 0.0436 m M−1 cm−1 was used. CAT was expressed in μmol H2O2 min−1 mg protein−1.
Protein quantification
Quantification of total proteins in samples used for enzyme analysis was based on Ref. 27 adapted to microplates, and the ratio between absorbances at 595 and 450 was applied to an analytical curve with six concentrations of bovine serum albumin ≥96% purity (Sigma-Aldrich).
Statistical analysis
The lethal and effect concentrations to 50% of the organisms exposed until 96 hpf (CL50–96h and EC50–96h) were calculated based on the logistic model using dose response curve package in software R version 4.0.2. 28
Biochemical biomarkers, GMS index, heart rate, total length, and swimming behavior endpoints were analyzed by ANOVA when data were normal and homoscedasticity or by Kruskal–Wallis when normality or homoscedasticity failed. Normality was checked by Kolmogorov–Smirnov test and homoscedasticity by Levene median test. When differences were detected, multiple comparison tests using Dunnett (ANOVA) or Dunn (Kruskal–Wallis) were used to evaluate differences between different treatments and controls. Frequencies of developmental abnormalities between fish exposed to different NH3 concentrations and controls were compared using two sample z-test for proportions. ANOVA, Kruskal–Wallis, and z-tests were based on SigmaPlot 12.0 software (Jandel Scientific, Erkrath, Germany).
Results
Mortality rate
Logistic modeling of the dose–response curves based on mortality rates of the embryo-larval stages of Danio rerio exposed during 72 h and 96 h to increasing concentrations of NH3 estimated lethal concentrations (in mg NH3 L−1) to 10% (LC10–72h = 2.19, LC10–96h = 1.54), 50% (LC50–72h = 2.61, LC50–96h = 2.07), and 90% (LC90–72h = 3.11, LC90–96h = 2.8) of exposed individuals (Fig. 1). Mortality rates of exposed embryo-larval did not increase at all tested concentrations until 48 hpf and varied from 0% to 10% (Fig. 1).

Mortality rates for the embryo-larval phase of Danio rerio exposed for 96 h to increasing concentrations of NH3.
General morphology score
A concentration dependent delay in embryo-larval development based on global morphology scores was detected. The lowest observed effect concentration (LOEC) for GMS decrease was 0.15 mg NH3 L−1, 7.2% of the LC50–96 h (Fig. 2A).
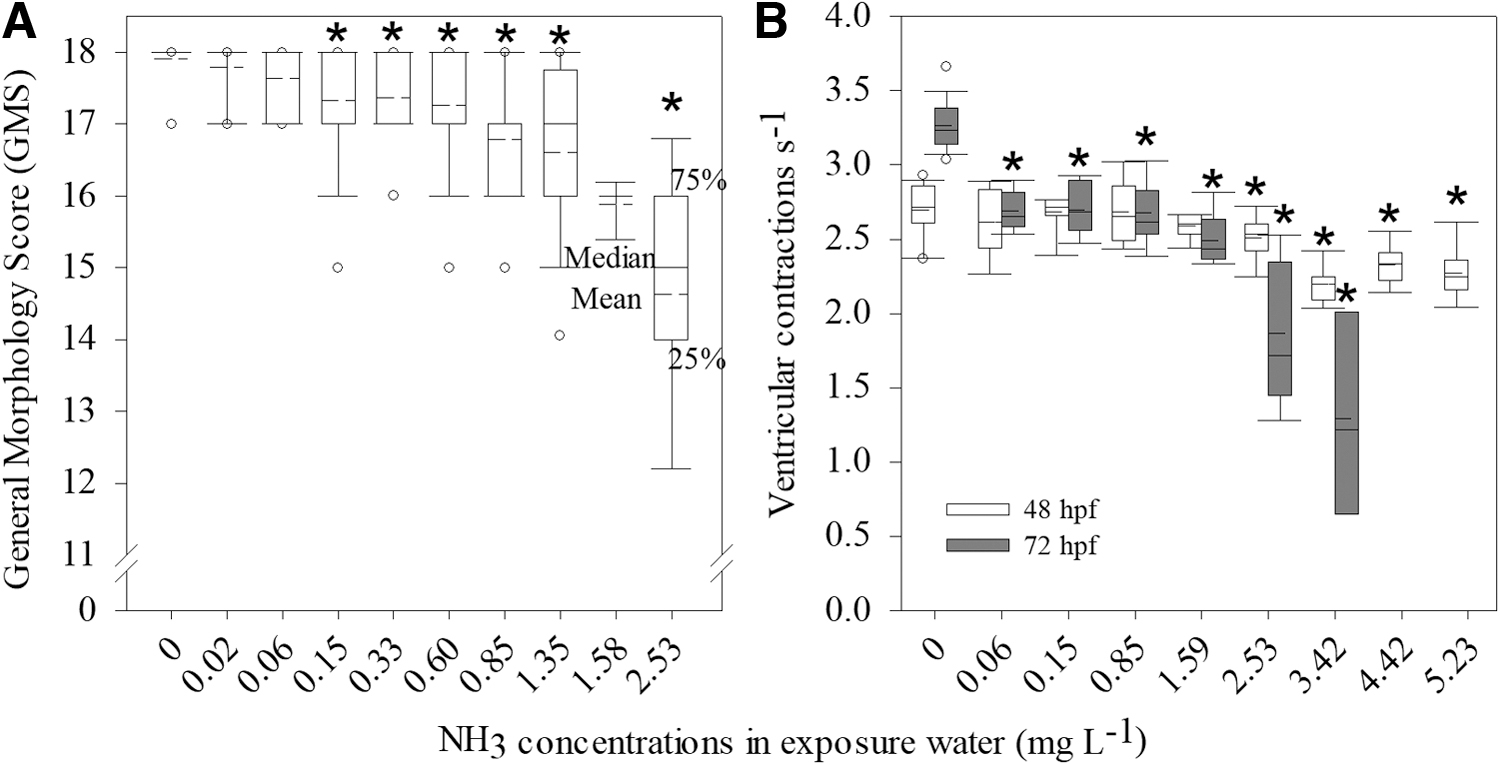
General Morphology Score at 96 hpf
Developmental abnormalities
Frequencies of developmental abnormalities indicated LOECs for blood stasis at 24 hpf, incomplete protrusible mouth at 72–96 hpf, absence of pectoral fin at 96 hpf, failure in swim bladder inflation at 96 hpf, and incomplete yolk sac absorption at 96 hpf of 0.85, 2.53, 3.41, 0.06, and 0.15 mg NH3 L−1, respectively (Table 1). The EC50 for failure in swim bladder inflation was equal to 0.18 mg NH3 L−1, 7.2% of the LC50–96h.
Frequency (Percentage) of Developmental Abnormalities in Danio rerio Larvae After 96 h of Exposure to Different Concentrations of NH3
Significant difference based on a z-test for the proportion of two samples (p < 0.05).
N = 44 exposed fish in control and 20 exposed fish at all NH3 concentrations.
Heart ventricular contractions
The LOEC for decreased heart ventricular contractions at 48 and 72 hpf was 2.5 and 0.06 mg NH3 L−1, equivalent to 120% and 2.8% of the LC50–96h, respectively (Fig. 2B). Heart ventricular contractions were reduced by 19% in embryos exposed to 3.4 mg NH3 L−1 at 48 hpf compared to controls and were more severely reduced at 72 hpf, ranging from a reduction of 18% in larvae exposed to 0.06% to 60% in larvae exposed to 3.4 mg NH3 L−1.
Spontaneous average and maximal swimming speeds and trajectories
The average and maximal swimming speeds decreased in larvae exposed to 1.35 NH3 L−1 (Fig. 3A, B). Swimming trajectory maps indicated a concentration dependent increase in frequency of exposed larvae swimming along the edge of the well (thigmotaxis) compared to control, and the LOEC was 0.33 mg NH3 L−1, 15.9% of the LC50–96h (Fig. 3C).

Maximum swimming speed
Total length at 168 hpf was reduced in larvae exposed to 0.607 (LOEC), 1.35, and 2.53 mg NH3 L−1 by 4.5%, 6.3%, and 6.5%, respectively (Fig. 3D).
Biochemical biomarkers
AChE activity was reduced in larvae exposed to 0.85 (LOEC), 1.58, and 2.53 NH3 L−1 by 25%, 35%, and 38% relative to control larvae mean activity of 0.567 μmol TNB min−1 mg protein−1 (Table 2). GST activity was induced in larvae exposed to 2.53 NH3 L−1 by 52% relative to control larvae mean activity of 0.36 μmol CDNB min−1 protein−1 (Table 2). CAT activity was induced in larvae exposed to 0.022 NH3 L−1 by 58% relative to control larvae mean activity of 6.35 μmol H2O2 min−1 protein−1 (Table 2).
Acetylcholinesterase, Catalase, Glutathione-S-Transferase, and Reactive Oxygen Species in Danio rerio Larvae After 96 h of Exposure to Different Concentrations of NH3
AChE: μmol TNB min−1 mg protein−1, CAT: μmol H2O2 min−1 mg protein−1, GST: μmol CDNB min−1 protein−1.
Statistically different from control (Anova followed by Dunnett, p < 0.05).
AChE, acetylcholinesterase; CAT, catalase; CDNB, 1-chloro-2,4-dinitrobenzene; GST, glutathione-S-transferase; TNB, 5-thio-2-nitrobenzoic acid.
Discussion
Un-ionized ammonia induced lethal and sublethal alterations in a concentration-dependent manner during the first 7days of Danio rerio embryo-larval development. Zebrafish embryos are predominantly ureotelic until hatching which prevents ammonia toxicity while inside the egg. 29 After hatch the ornithine-urea cycle is inactivated, and ammonia is excreted by diffusion mediated by the glycoprotein Rhesus (Rh) present in larval skin, yolk sac epithelium, and developing gill.9,29 In addition, before hatch the fish egg chorion protects the embryo from direct contact with high levels of environmental ammonia (HEA), 9 which makes the embryo more resistant to HEA exposure than hatched larvae both in marine and freshwater fishes. 30
These facts are coherent with our results showing that almost no mortality occurred until 48 hpf (before hatch), with a sharp increase in mortality rates from 72 to 96 hpf (Fig. 1). In addition, it has been shown that tissue ammonia concentrations more than doubled in zebrafish larvae exposed to HEA after 4 dpf, 31 also justifying this steep increase in mortality verified after hatch. A similar pattern of increased resistance to HEA exposure in embryos of Oncorhynchus mykiss before hatch has also been verified. 32
Blood circulation initiates at 24 hpf in zebrafish, 33 and is important for the development of organs such as the pancreas and liver during early embryonic stages. 34 In addition, vascularization and subsequent blood circulation plays a crucial role in zebrafish swim bladder development and inflation after 96 hpf, as it is responsible for the diffusion of gases into the swim bladder. 33 In our study the detected concentration dependent increase in the frequency of larvae with uninflated swim bladders at 96 hpf (Table 1) may have been a consequence of the circulatory problems verified. These include the increase in the frequency of exposed larvae with blood stasis at 24 hpf (Table 1), as well as the decrease in heart rates of 72 hpf exposed larvae (Fig. 2B). In addition, these circulatory issues might also be involved in the increased frequency of exposed larvae with improper yolk sac absorption (Table 1), essential to provide metabolic energy for organ development 35 and growth, causing the detected reduced length of exposed larvae at 168 hpf (Fig. 3D).
The LC50–96h of 2.07 mg NH3 L−1 estimated in this study for zebrafish exposed from 0 to 96 hpf is in the 75% quartile of the median LC50 of 1.0 mg NH3 L−1 for early life stages of fishes compiled in Table 3. Zebrafish embryos and early larvae have become an important ecotoxicological model to assess ecological risk and monitor the quality of aquatic freshwater ecosystems contaminated by complex chemical mixtures. 17 Although zebrafish early life stages are relatively resistant to lethal effects of NH3 compared to other fishes (Table 3), significant levels of NH3 have been found in aquatic ecosystems monitored with zebrafish embryo-larval tests. Mortality of zebrafish exposed during 96h to water samples from site S6 of the Capibaribe River in Brazil was 45%, contaminated by 1.56 mg NH3 L−1 (0.75 toxic units of NH3 considering our estimated LC50–96h), low concentrations of polycyclic aromatic hydrocarbons, and other unmeasured contaminants. 12 The expected mortality contribution from NH3 alone in this sample is relatively close to the 45% mortality verified, suggesting that un-ionized ammonia is a major contributor to detected lethal toxicity.
Lethal Concentration of Un-ionized Ammonia to Early Life Stages of Fishes
d, days; LC50, lethal concentrations to 50% of exposed individuals after respective exposure durations.
In another study, mortality of zebrafish embryo-larval stages exposed during 96 h to water samples from the middle course of the Matasnillo river in Panama was 47.5%, contaminated by 0.04 mg NH3 L−1 (0.02 toxic units of NH3), 58 measured contaminants, including pharmaceuticals, pesticides, and industrial chemicals. 14 The 47.5% detected mortality is significantly higher than the expected mortality contribution from NH3 alone and suggests that in this case other contaminants are contributing to the detected toxicity. Zebrafish exposed to water samples from site KT in May in Kenting National Park in Taiwan were 100%, contaminated by 0.38 mg NH3 L−1 (0.18 toxic units) pharmaceuticals, personal care products, and other unmeasured contaminants. 15 The 100% detected mortality is higher than the expected mortality contribution from NH3 alone and suggests that other contaminants are contributing to the detected toxicity.
USEPA water quality criteria for the protection of freshwater aquatic life establish maximum concentrations of ammonia for acute exposure (1 h) and chronic exposure (30 days) adjusted to pH 7.0 and temperature of 20°C equal to 0.0673 and 0.0075 mg NH3 L−1, respectively. 2 Our results indicate that even the less restrictive criteria for acute toxicity of 0.0673 mg NH3 L−1 would not protect Danio rerio from sublethal toxicity based on the LOEC of 0.06 mg NH3 L−1 for decreased heart rate and absence of swim bladder inflation, both equivalent to 2.8% of the LC50–96h.
The development and inflation of the swim bladder during early fish development confer neutral buoyancy that is essential for proper swimming and initial capture of exogenous food, growth, predator evasion, and recruitment (Czesny et al. 2005). Therefore, improper development and/or function of the swim bladder caused by exposure to contaminants is of high ecological relevance. Exposure of zebrafish embryo-larval stages to a wide range of contaminants can lead to failure in swim bladder inflation, and 50% effective concentrations (EC50) for this endpoint range from 0.06 mg L−1 for the pharmaceutical lovastatin to 39 mg L−1 for the aromatic p-toluidine, among 20 tested chemicals. 36 The EC50 for lack of swim bladder inflation verified in this study of 0.18 mg NH3 L−1 (8.6% of the LC50–96h) indicates that NH3 is among the most potent contaminants for this endpoint in zebrafish.
Exposure to neurotoxicants such as AChE inhibitors was hypothesized to potentially disrupt swim bladder function, 37 as inflation and deflation involve the autonomic nervous system and acetylcholine based synapses. 38 Our results indicated a LOEC for AChE inhibition in zebrafish of 0.85 mg NH3 L−1 (Table 2) in association with high frequencies of larvae without inflated swim bladders (Table 1). It has been demonstrated that exposure of the fish Cirrhinus mrigala to 0.34 mg NH3 L−1 caused brain AChE inhibition and behavioral alterations, including sluggish movement and loss of balance, 39 and it has been suggested that AChE inhibition might be a relevant mechanism for the neurotoxicity of ammonia. 40 Zebrafish larvae at around 96 hpf need to rise to the water surface and gulp air for initial swim bladder inflation, 41 and the detected decrease in swimming speed and increased thigmotaxis (Fig. 3) suggest that swimming behavioral alterations might have influenced the increase in frequencies of larvae without inflated swim bladders.
In addition, after initial inflation, further adjustments in volume are essential to reach neutral density, which involve cholinergic axons 38 that may have been affected. The detected thigmotaxis in NH3 exposed zebrafish larvae at 168 hpf also indicates a smaller capacity to explore the environment, which may also decrease prey detection rates at a phase where initial feeding is essential for survival. 42 Different sensorial systems are important at this critical transition, and the number and function of lateral-line hair cells decreased in zebrafish at 96 hpf exposed to 1.12 mg NH3 L−1. 43 The concentration dependent response and LOEC of 0.15 mg NH3 L−1 (7.2% of the LC50–96h) verified in this study for the GMS index indicate that un-ionized ammonia is a potent inhibitor of zebrafish embryo-larval development. Application of the GMS index to monitor surface water of 10 sites along the Capibaribe River in Northeast Brazil found a significant correlation between delayed development and increased NH3 concentrations, suggesting that environmentally relevant NH3 can affect zebrafish early development. 12
Conclusions
Our results indicate that zebrafish early developmental stages are sensitive to environmentally relevant NH3 concentrations based on lethal and sublethal biochemical, developmental, and behavioral endpoints, providing information about mechanisms of ammonia toxicity and its potential contribution to the toxicity of complex environmental mixtures when zebrafish is used in water quality monitoring.
Footnotes
Acknowledgments
Authors are very grateful to all students who contributed during experimental exposures and testing and to unknown reviewers who helped to increase the quality of this publication.
Authors' Contributions
C.F.M.Jr.: Conceptualization, data curation, methodology, investigation; writing, original draft, review, formal analysis; M.K.M.A.: Investigation; S.M.V.S.: Investigation; R.N.A.: Investigation; P.S.M.C.: Conceptualization, Methodology, Writing original draft, Writing—review & editing, Investigation, Formal analysis, Supervision, Project administration, Funding acquisition.
Data Access Statement
Data, associated metadata, and calculation tools are available from the corresponding author (paulo.smcarvalho@ufpe.br).
Ethics Statement
Animal management and embryo exposure were performed according to a protocol approved by the Animal Experimentation Ethics Committee of the Federal University of Pernambuco (UFPE).
Disclosure Statement
No competing financial interests exist.
Funding Information
Célio Freire Mariz Jr. was sponsored by a MSc fellowship from ‘‘Fundação de Amparo à Ciência e Tecnologia de Pernambuco - FACEPE’’. Paulo SM Carvalho (Grant 312308/2016-7) is a research fellow of CNPq.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.