
Abstract
The liver plays a very important role in physiological processes of the human body. Liver regeneration has developed into an important area of study in liver disease. The Mtz (metronidazole)/NTR (nitroreductase)-mediated cell ablation system has been widely used to study the processes and mechanisms of liver injury and regeneration. However, high concentrations and toxic side effects of Mtz severely limit the application of the Mtz/NTR system. Therefore, screening new analogs to replace Mtz has become an important means to optimize the NTR ablation system. In this study, we screened five Mtz analogs including furazolidone, ronidazole, ornidazole, nitromide, and tinidazole. We compared their toxicity on the transgenic fish line Tg(fabp10a: mCherry-NTR) and their specific ablation ability on liver cells. The results showed that Ronidazole at a lower concentration (2 mM) had the same ability to ablate liver cells comparable with that of Mtz (10 mM), almost without toxic side effects on juvenile fish. Further study found that zebrafish hepatocyte injury caused by the Ronidazole/NTR system achieved the same liver regenerative effect as the Mtz/NTR system. The above results show that Ronidazole can replace Mtz with NTR to achieve superior damage and ablation effects in zebrafish liver.
Introduction
The liver is a vital human organ crucially involved in digestion, metabolism, filtration, detoxification, protein synthesis, glycogen storage, and biliary excretion. Liver diseases have become some of the most important diseases affecting human health over the past few decades. About 2 million deaths per year, worldwide, occur from various liver diseases, and more than one fifth of people in China suffer from liver disease. 1 Early lesions of the liver can resolve by regeneration and self-repair with or without drug intervention. However, once massive fibrosis develops in the liver, it is likely to progress to liver cirrhosis and liver failure. The only effective treatment is liver transplantation.2,3 The sources of the donor liver and immune rejection of the transplant have become important factors determining the prognosis in advanced liver disease. Whether the remaining liver tissue cells can be re-differentiated and trans-differentiated into new liver cells and then generate new functional liver organs is the ultimate goal of all researchers.
Various animal models of liver injury and regeneration have been used to explore molecular mechanisms of liver regeneration, to advance treatments for end-stage liver disease.4–7 Zebrafish demonstrate rapid, effective regeneration and repair after injury, particularly compared with mammals such as mice. 8 Recently, a variety of methods have been applied to the investigation of zebrafish liver injury and regeneration.9,10
The Mtz (metronidazole)/NTR (nitroreductase) system is ablative regenerative research model that has been widely used to study organ tissue-specific injury and repair.11–14 The basic principle is that cell-specific expression of NTR interacts with an externally added nitro-analog substrate, Mtz, producing cytotoxicity in specific cells, and targeted tissue cell damage. However, the application of the Mtz/NTR system has shown that Mtz inevitably causes toxic side effects with increase in concentration and prolongation of treatment time.12–14 This toxicity greatly limits the application of Mtz/NTR in the field of regenerative medicine research. This has led many researchers to screen an alternative drug to Mtz with NTR-induced cell ablation system.
Studies have shown that a variety of Mtz analogs have the same, or similar, effects in inducing nerve cells ablation and regeneration. 15 However, it is unknown whether these Mtz analogs have similar effects on liver tissue regeneration and this was the objective of our study. Five Mtz analogs were evaluated for their effects on injury, death, and regeneration of transgenic zebrafish hepatocytes specifically expressing NTR, examining their efficacy and drug side effects.
Materials and Methods
Ethics statement
All experimental protocols were approved by the Chengdu Medical College (Chengdu, China), and the experiments were carried out in accordance with the approved guidelines. The zebrafish facility and study were approved by the Institutional Review Board of Chengdu Medical College (Chengdu, China). Zebrafish were maintained in accordance with the Guidelines for Experimental Animal Welfare from the Ministry of Science and Technology of the People's Republic of China (2006) and the Institutional Animal Care and Use Committee protocols of Chengdu Medical College.
Zebrafish embryo culture
Adult fish and embryos were raised and maintained under standard laboratory conditions according to Institutional Animal Care and Use Committee protocols. They were staged as previously described. 16 Fish embryos were treated with 0.003% PTU from 24 hpf to inhibit pigmentation.
Construction of the transgenic plasmid and line
To generate the Tg(fabp10a:mCherry-NTR) transgenic line, we separately polymerase chain reaction (PCR) amplified the mCherry and NTR encoding sequences from our previously plasmids. 17 ClonExpress Ultra One-Step Cloning kit (Vazyme, Nanjing, China) was used to insert mCherry and NTR fragments fusion into the fabp10a:HA-zc3h8-T2A-mCherry plasmid after the excision of HA-zc3h8-T2A-mCherry with Nhe I/Not I. The fabp10a:mCherry-NTR plasmid was co-injected with I-Sce I meganuclease into one-cell-stage ABGO line embryos as previously described. 18 Candidate founder embryos were selected based on specific expression of mCherry in the livers raised to adulthood. Germline transmission and screening were performed on these zebrafish, and the most representative transgenic lines were selected for the following experiments.
Whole-mount in situ hybridization and antibody staining
Whole-mount in situ hybridization were performed as previously described. 19 Probes for the vitamin D binding protein (gc) and hematopoietically expressed homeobox (hhex) transcripts were generated from PCR products using the Dig RNA labeling kit (Roche Applied Science, Penzberg, Germany). Embryos were imaged using Olympus microscope (SZX16). Antibody staining was performed as previously described using the following antibodies: anti-mCherry (1:1000; Abcam, Cambridge, MA) and anti-2F11 (1:1000; Abcam).
Quantitative real-time PCR
Quantitative real-time PCR were performed for hepatocyte markers ceruloplasmin (cp) and vitamin D binding protein (gc) and hepatoblast developmental markers hematopoietically expressed homeobox (hhex), forkhead box a3 (foxa3), and SRY (sex determining region Y)-box 9b (sox9b) using Fast SYBR Green qPCR Master Mix with UDG (Exongen, Chengdu, China) following the manufacture's protocols, and their levels were normalized to that of β-actin. The primer used for these analyses was as previously described. 20
Chemical treatment
Mtz (Cat. No.: A600633; Sangon Biotech, Shanghai, China), furazolidone (Cat. No.: HY-B1336; MedChemExpress, Shanghai, China), ronidazole (Cat. No.: HY-B0565; MedChemExpress), ornidazole (Cat. No.: HY-B0508; MedChemExpress), nitromide (Cat. No.: HY-B0945; MedChemExpress), and tinidazole (Cat. No.: HY-B0177; MedChemExpress) were used in this experiment.
Confocal imaging
Larvae were anesthetized by 0.01% Tricane and mounted in 1% low-melting agarose containing 0.01% Tricane. Nikon A1 confocal microscope was used for imaging of live and antibody-stained larvae.
Quantification and statistical analysis
The fluorescence intensity and the area of liver sizes were quantified with ImageJ 1.47 software (National Institute of Health). Student's t-test was used for statistical analysis by GraphPad Prism Vision 8.0 for Windows. p < 0.05 was considered statistically significant.
Results
Toxic effects of five Mtz analogs on Tg(fabp10a:mCherry-NTR) larval zebrafish
Toxicity and adverse effects are an inevitable consequence of drug. In this study, five Mtz analogs were selected as candidate drugs: ronidazole, ornidazole, tinidazole, furazolidone, and nitromide. The toxic effects of these drug candidates were examined on Tg(fabp10a:mCherry-NTR) transgenic zebrafish larvae. About 10 mM concentrations, as used in previous reports, 4 were used as starting concentrations for all drugs. The starting time and duration of drug treatment were also based on previous studies.20–22 Juveniles at 5 days postfertilization (dpf) were selected, and drug was removed after 24 h of exposure (up to 6 dpf), and deformity and survival of embryos after drug treatment in each group was recorded (Fig. 1A and Supplementary Fig. S1A).
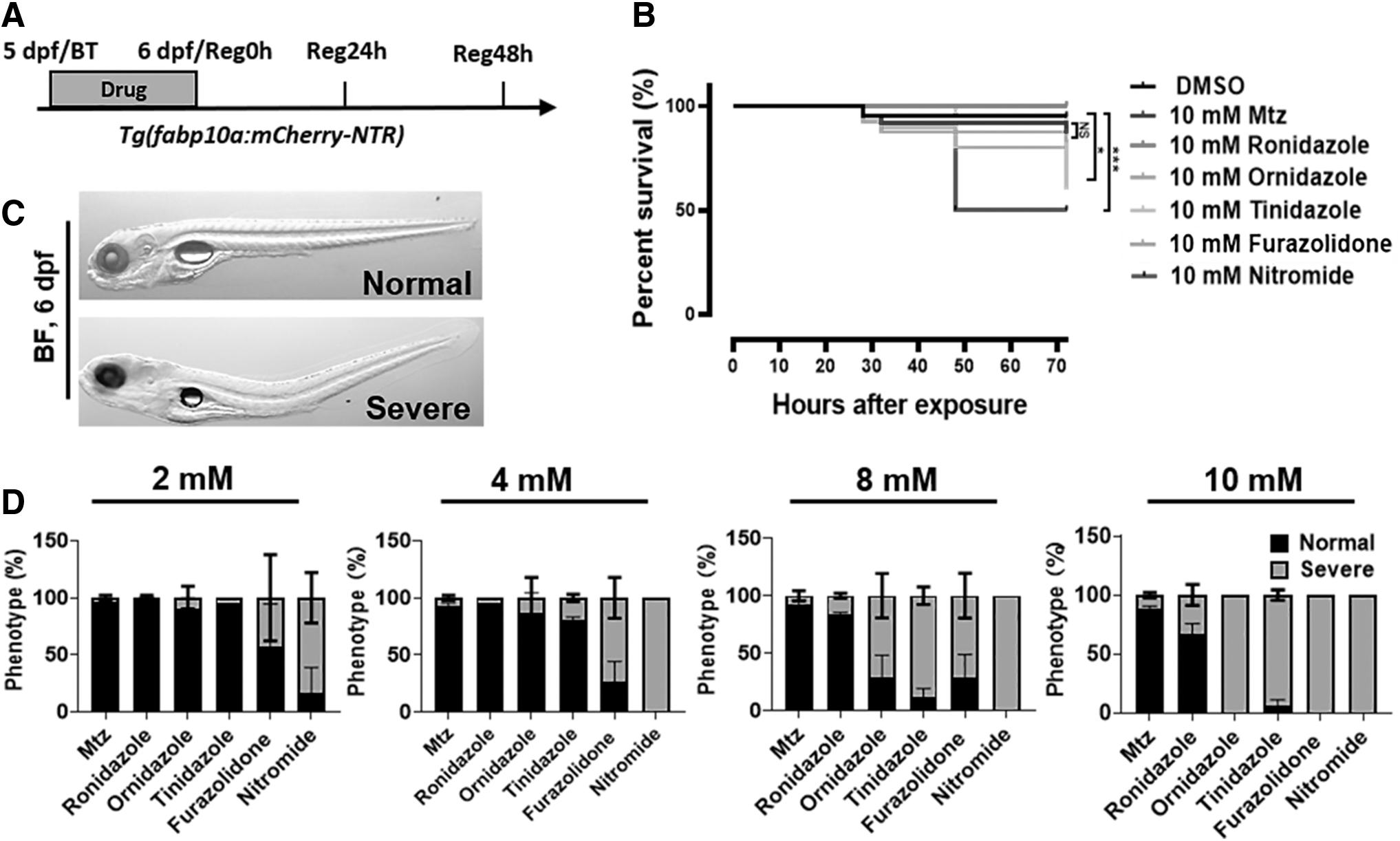
Zebrafish larvae toxicity test showed that ronidazole and furazolidone had the same low toxic side effects as Mtz.
Experimental results showed that the survival rate of juvenile fish in the ronidazole, ornidazole, and furazolidone groups within the range of Reg 48 h (up to 8 dpf) were not significantly different from the Mtz treatment group (all treatments at 10 mM). Survival rate of juveniles in tinidazole and nitromide groups was significantly lower than that in the Mtz treatment group within Reg 48 h (Fig. 1B and Supplementary Fig. S1B). At Reg 48 h, 10 mM tinidazole treatment reduced the survival rate of juveniles to ∼55%; nitromide decreased the survival rate to 50% at Reg 24 h. These results indicated that tinidazole and nitromide have definite lethal effects on juvenile fish at the concentration of 10 mM. Ronidazole, ornidazole, and furazolidone showed no significant difference from Mtz in overall lethality.
Phenotypic change after drug use is an additional important indicator of drug side effects; spinal curvature, swim bladder disappearance, and pericardial swelling may be evident in zebrafish embryos. Each of the five Mtz analogs were formulated at 2, 4, 8, and 10 mM, and juveniles were exposed to each of these four concentrations of drugs. The ratio of normal to affected phenotypes (Fig. 1C and Supplementary Fig. S1C) was used to judge toxic side effects on juvenile fish. Ornidazole and tinidazole at concentrations of 8 mM and above caused >50% of juveniles to be severely affected. Nitromide and furazolidone at a concentration of 2 mM severely affected >50% of the juvenile fish. However, neither ronidazole nor Mtz severely affected >50% of fish at concentrations of 10 mM and below (Fig. 1D and Supplementary Fig. S1D).
Ronidazole can achieve the same cell ablation efforts as Mtz at lower concentrations
To study the specific ablation ability of various drugs on liver cells, we first constructed and screened a liver-specific Tg (fabp10a: mCherry-NTR) transgenic zebrafish line that stably expressed the NTR fusion protein in the liver. Using the half concentration of each drug causing a 50% teratogenic rate of fish as the upper limit, a series of concentrations were examined on liver-specific ablation experiments (ensuring sufficient survival of zebrafish larvae) (Fig. 2A). In addition, Tg(fabp10a: mCherry-NTR) at 5 dpf were treated with various drug concentrations over 24 h to compare liver ablation of zebrafish larvae in each group. The results showed that tinidazole and nitromide had little effect on liver ablation at low concentrations (1 and 0.25 mM concentration), weak ablation at high concentrations (4 and 1 mM concentration), but considerably above the effect of Mtz at 6 mM concentrations.

Comparison of cell ablation efficiently between different Mtz analogs in Tg(fabp10a:mCherry-NTR) larvae.
Ornidazole had very weak/minimal ablative effect at 1 and 2 mM but cause ∼50% liver ablation at 4 mM. In contrast, furazolidone caused liver ablation at 0.25 mM, more pronounced at 0.5 and 1 mM, whereas Mtz had a complete liver ablation effect at 10 mM concentrations. Ronidazole, however, showed an ablative effect on the liver at 0.5 mM. The ablative effect of ronidazole at 2 mM concentration is comparable with that of Mtz at 10 mM concentrations (Fig. 2A, B). Based on the results, ronidazole at 2 mM has the most ideal liver cell ablation effect (closest to that caused by Mtz at 10 mM).
Zebrafish liver regenerates normally after ronidazole ablation, and the regeneration mechanism is similar to Mtz
Whether the liver can regenerate normally after liver ablation is an important prerequisite for determining whether the ablation drug can be used as a specific tool for the study of liver regeneration (Fig. 3A). The livers of Tg(fabp10:mCherry-NTR) transgenic zebrafish were completely ablated after treatment with ronidazole 2 mM, and regenerated following drug withdrawal (Fig. 3B). At the time points of Reg 48 h and Reg 72 h, the regeneration of the zebrafish liver in the ronidazole group was nearly the same as in the Mtz group; zebrafish liver recovery to pretreatment size at Reg 72 h. Previous studies in zebrafish have shown that single-tubular cells were trans-differentiated to liver progenitor cells to promote regeneration of the entire liver.4,22 We examined whether the regeneration process after ronidazole treatment was similar. Antibody staining results showed that ronidazole regeneration was also accompanied by a large number of bifunctional cells (Fig. 3C), indicating that hepatocytes regeneration was accompanied by corresponding trans-differentiation, which is the same as Mtz treatment group.

Ronidazole showed the similar process as Mtz for inducing liver injury and regeneration in zebrafish larvae.
Other studies 21 have shown that hepatocyte-like progenitor cell markers such as hhex, foxa3, and sox9b are highly expressed in the early stages of liver regeneration, with gradual increase in liver cells and almost recovery. Expressions of cp and gc is also upregulated in regenerated liver. In this study, we found that regenerated livers treated with Reg 0 h (up to 6 dpf) and ronidazole were similar to those treated with Mtz, and were accompanied by high levels of expression of hepatoid progenitor markers such as hhex, foxa3, and sox9b, which were restored to normal after regeneration (Fig. 4A, B). However, after ronidazole treatment, the damaged hepatocytes were ablated, and the expression of hepatocyte marker molecules cp and gc were significantly downregulated, with gradual restoration accompanying the gradual recovery of the liver during regeneration (Fig. 4A, C).

The mechanism of liver regeneration after ronidazole treatment is similar to that of Mtz.
Discussion
The Mtz/NTR system involves interaction of NTR specifically expressed in tissues or cells by addition of a substrate such as Mtz at a specific time, producing cytotoxicity to specific tissues or cells to achieve killing at a specific time point. Because of its ability to achieve specific localized cell damage, it is widely used in research into ablation and regeneration of various tissues and organs.11,14 The key factors affecting the function of the system include: the expression of NTR enzyme and the availability of substrate. NTR expression is directly related to the promoter efficiency in the transgenic zebrafish construct; the cell type also determines whether NTR interacts with Mtz, and so on. There are also reports that partial replacement of the amino acid sequence of NTR can also increase enzymatic efficiency to a certain extent. 23
Efforts to optimize NTR's ability to achieve the most ablated cells by re-engineering new NTR variants are often time-consuming and labor-intensive. Screening for suitable NTR substrates is relatively easy to achieve. Combining all our research results, nitromide, had the highest cell lethality, but required concentrations of 2 mM and above, which caused deformity exceeding 50% of juvenile fish (Fig. 1D). However, nitromide has very limited killing effect on hepatocytes at boundary concentrations (Fig. 2A, B).
The cytotoxicity of ornidazole, tinidazole, and furazolidone, at the same concentration (2 mM concentration), was less than that of nitromide, although greater than that of ronidazole. Although furazolidone can effectively kill the liver at a lower concentration (0.25 mM), and its ablation effect is even better than that of ronidazole at this concentration overall, in terms of embryotoxicity and lethality to liver tissue together, the effect of ronidazole is the best among the five Mtz analogs. Furthermore, comparing ronidazole and Mtz under the conditions of use, the toxicity to embryos and the effectiveness of specific liver damage ronidazole is undoubtedly more suitable than Mtz as a substrate for NTR to produce liver ablation.
Footnotes
Authors' Contributions
Q.Z., X.D., and P.W. designed the experimental strategy, analyzed data, and wrote the article. T.C. and H.L. performed liver injury and antibody staining experiments. Y.C. and P.L. performed all the other experiments.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Youth Program of National Natural Science Foundation of China (No. 32000567), Foundation of Chengdu Medical College (No. 19Z039), Graduate Innovation Fund of Chengdu Medical College (No. YCX2021-27), and Disciplinary Construction Innovation Team Foundation of Chengdu Medical College (No. CMC-XK-2101).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.