
Abstract
Zebrafish is a popular animal model in regeneration studies due to their ability to regenerate the heart. Primary cardiomyocytes could be an alternative tool for studying the intrinsic mechanisms of cardiovascular disease in vitro. Thus, our objective is to develop an efficient protocol to isolate primary cardiomyocytes from zebrafish hearts. Low concentration of digestive enzyme (0.5 mg/mL collagenase type II) was utilized in our protocol to obtain single-cell suspension. The ventricles were fragmented, mechanically pipetted, and constantly shaken to ensure adequate contact between the tissues and the enzyme. Preplating the cell suspension onto culture plates for 2 h helped remove cardiac fibroblasts. The purity of isolated cells was validated by flow cytometry analysis of transgenic zebrafish with cardiomyocyte-specific expression of enhanced green fluorescent protein (EGFP) or endothelial cell-specific expression of mCherry. Quantitative real-time PCR analysis revealed a high level of the purity, with cardiac fibroblasts, endothelial cells, and epicardial cell markers scarcely detected in the purified cells. Altogether, this study established a reproducible protocol for isolating primary cardiomyocytes with high purity and activity from adult zebrafish hearts that can be cultured in vitro for up to 4 weeks. This protocol provides a valuable tool for studying the intrinsic mechanisms of cardiovascular disease in vitro using primary cardiomyocytes.
Introduction
The zebrafish (Danio rerio) is a remarkable model organism for investigating myocardial regeneration and repair, due to its unique ability to completely regenerate an adult heart following injury. However, recent studies have identified certain limitations in conducting in vivo experiments on adult zebrafish, such as variability in phenotypes resulting from drug administration or surgical procedures.1–3 To overcome these limitations, in vitro studies based on the isolation and cultivation of adult zebrafish cardiomyocytes have emerged as a promising alternative approach to investigate the underlying mechanisms of cardiovascular disease. 4
One unique advantage of primary adult cardiomyocytes is their similarity in both morphological and functional characteristics to cardiomyocytes found in living organisms. This differentiates them from immortalized myocardial cell lines, such as HL-1 (mouse atrial myocyte line) and H9c2 (embryonic rat myoblast line), which lack the contractile properties of primary cardiomyocytes.5–7 While human-induced pluripotent stem cell-derived cardiomyocytes (hiPSC-CMs) have emerged as an alternative model system for in vitro disease pathophysiology,8–10 they still exhibit a marked immature phenotype even at the terminal differentiation stage.11,12 Isolated primary cardiomyocytes offer a potential solution to these issues, as well as a precise visualization of cellular structures and localization of intracellular molecule. 13
Powell and Twist 14 first described a collagenase perfusion technique for isolating cells from adult rat myocardium in 1976, which has since served as the foundation for numerous studies on cardiomyocyte isolation and culture in rodents.13–17 However, this method relies on perfusion and multiple enzyme digestion systems, making it unsuitable for the tiny zebrafish, which lack easily accessible coronary vessels. Alternatively, Vidyasekar et al. 18 demonstrated a simpler two-step approach for isolating and purifying neonatal cardiomyocytes from ventricular tissue using enzymatic digestion and mechanical agitation. This method holds potential to be adapted to zebrafish cardiomyocyte isolation and may offer a more feasible option.
Currently, there are only a limited number of reported methodologies for isolating adult zebrafish cardiomyocytes, and a universal method that produces high-quality and viable cardiomyocytes has yet to be established.19–21 Ten years ago, Kim et al. 22 described a protocol for isolating epicardial cells from adult zebrafish hearts, and Sander et al. 20 subsequently provided a detailed methodology for isolating and culturing zebrafish cardiomyocytes in vitro, although the entire process can take up to 4–5 h. Zeng et al. 21 attempted a three-dimensional (3D) approach using fibrous polyglycolic acid scaffolds to culture zebrafish cardiomyocytes but encountered premature disintegration of the scaffolds after 15 days, resulting in cardiomyocyte detachment and an inability to maintain long-term culture.
To address these challenges, we have developed an efficient, reproducible, and cost-effective protocol for rapidly isolating cardiomyocytes from adult zebrafish hearts. Our method is based on enzymatic digestion and mechanical isolation, incorporating a simplified digestion step and a short isolation time. This protocol offers a new option for researchers who wish to utilize adult zebrafish cardiomyocytes for in vitro studies but find current methods overly complex. Our novel method could broaden the potential applications of zebrafish models in cardiovascular research.
Materials and Methods
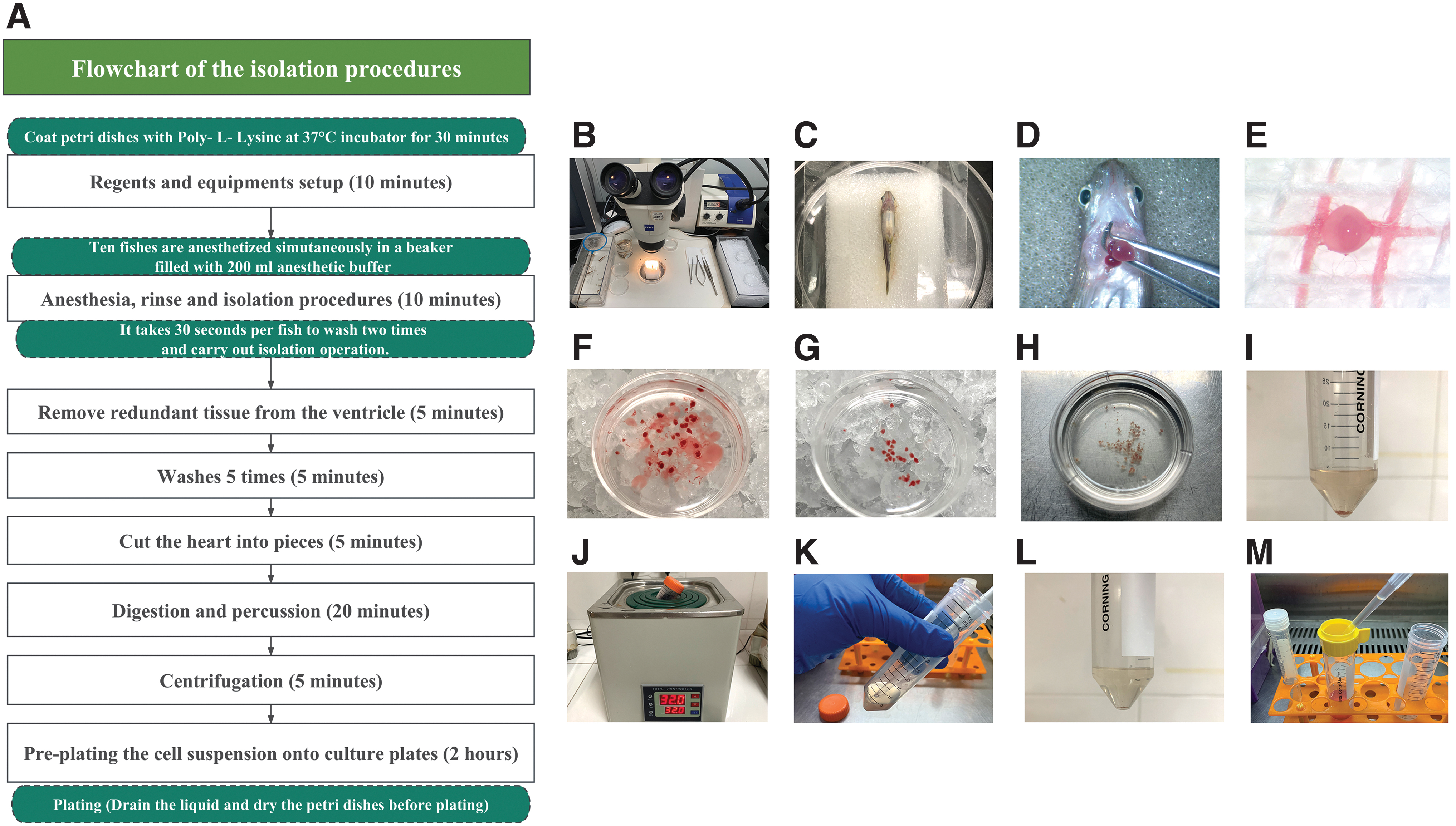
We depict the isolation procedures by flowchart (Fig. 1A).
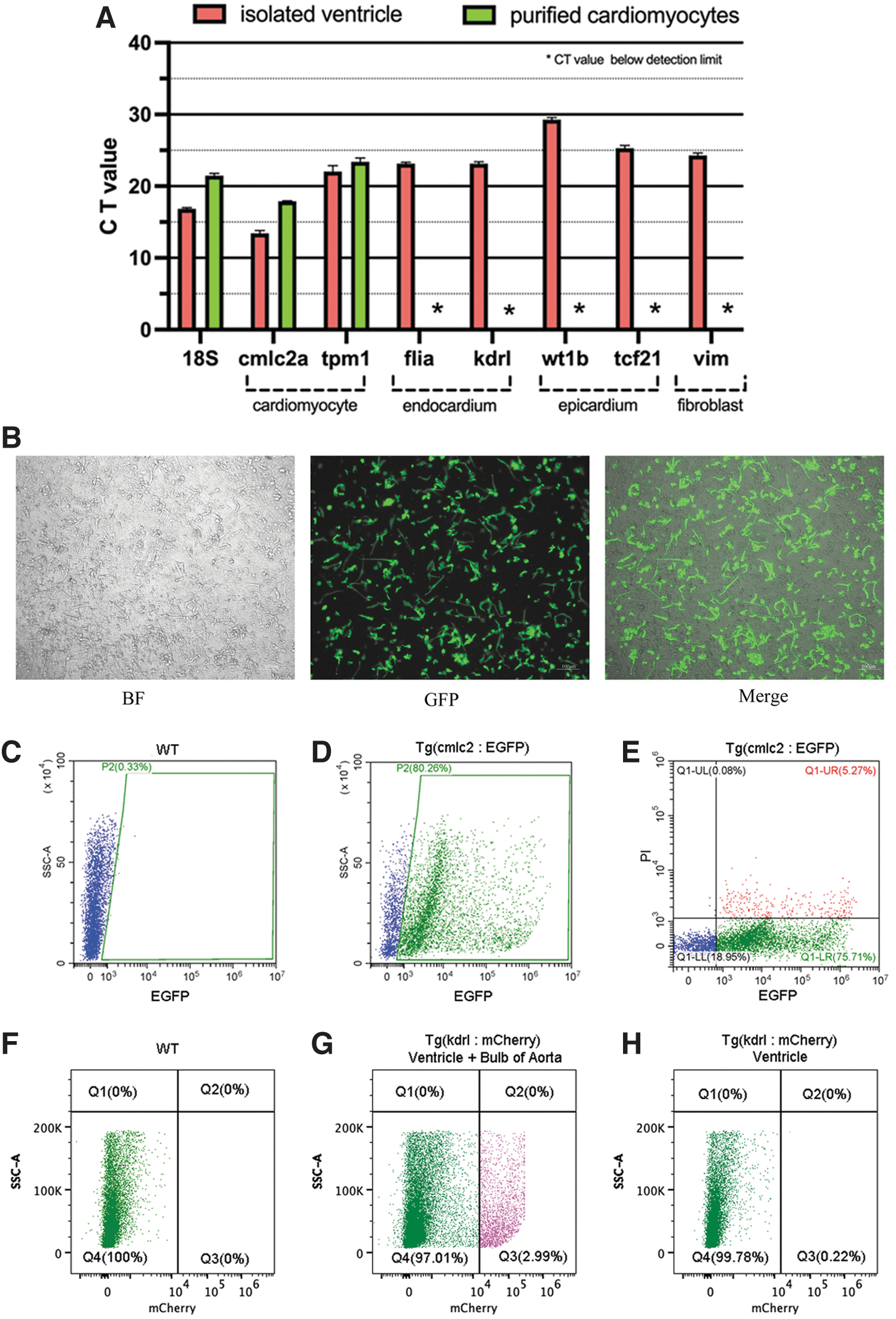
Flowchart and critical procedures in the isolation of cardiomyocytes from zebrafish hearts.
Reagents
Zebrafish (Danio rerio), 6–12 months old, wild-type (WT), transgenic line: Tg(cmlc2: enhanced green fluorescent protein [EGFP]) whose cardiomyocytes are labeled with green fluorescence, and Tg(kdrl:mCherry), whose endothelial cells are labeled with red fluorescence. All zebrafish are maintained at 28.5°C on a 14-h light and 10-h dark cycle.
Digestive buffer: 5 mg collagenase II (Cat. No. C6885-1G; SIGMA) and 10 mL Hanks' balanced salt solution (HBSS) (cold and sterile; Gibco).
Culture medium (90% DMEM/F-12 [LOT 2085358; Gibco], 10% fetal bovine serum [FBS]; Cat. No. C0234; Gibco, Australia), 1% ITS Liquid Media supplement (100 × ; Cat. No. 13146; SIGMA) and 1% penicillin-streptomycin (P/S, 100 × ; LOT 2019321; Gibco).
Poly-
4′,6-diamidino-2-phenylindole (DAPI) (Cat. No. P0131; Beyotime).
Tricaine (Working solution 1.3 mM; A5040-25G; Fluka).
α-sarcomeric actin mouse monoclonal antibody, 1:500 (Cat. No. A2172; Sigma).
Tropomyosin rabbit polyclonal antibody, 1:500 (Cat. No. AF8217; Beyotime).
Goat anti-Rabbit IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor™ Plus 555 (Cat. No. A32732; Invitrogen).
Goat anti-Mouse IgG (H+L) Cross-Adsorbed Secondary Antibody, Alexa Fluor™ 488 (Cat. No. A-11008; Invitrogen).
Propidium iodide (PI; Cat. No. 556463; BD Pharmingen).
Binding Buffer (Cat. No. 556454; BD Pharmingen).
TRIzol reagent (Cat. No. 15596026; ThermoFisher).
Fasting gDNA Dispelling RT SuperMix FastKing (Cat. No. KR118-02; TIANGEN).
Talent qPCR PreMix (SYBR Green) (Cat. No. FP209-02; TIANGEN).
Equipment
Three sets of sterile operating instruments (2 micro forceps, 2 micro scissors, and 1 tissue forceps for isolation operation; 1 micro forceps and 1 micro scissors for disassembling tissues into pieces in vertical flow clean bench).
Cell culture dishes (Corning, 35 and 60 mm) and cell culture plates (48- and 24-well).
Twenty milliliters Syringe, Sterilization filter (LOT R1CB00325; Millex), 100 μm Cell Strainers (LOT 21139830; Biosharp).
Centrifuge tubes (Corning, 15 and 50 mL).
Sterile micropipettes for Eppendorf pipettes, 1000 μL.
Dissecting stereomicroscope (Zeiss, Stemi 2000-CS).
Wet operation sponge (self-made).
Water bath set to 32°C.
Absorbent cotton ball and Nonwoven Gauzes.
Flow Cytometer (Beckman Coulter-CytoFLEX).
Fluorescence ration PCR instrument (ABI 7500).
Ultrasonic Cell Disruptor (Sonica Q700).
3D images using Laser Scanning Confocal Microscope (Nikon A1R HD25).
Inverted fluorescence microscope (Zeiss-Axio Vert.A1).
Procedures of isolation
Step 1: Reagent setup (Fig. 1B)
Add 5 mg collagenase II into 10 mL HBSS solution and run the solution through a sterilized filter before use.
Prepare two 60 mm dishes filled with 4 mL 75% ethyl alcohol, two 60 mm dishes filled with 4 mL HBSS, and two 35 mm cell culture dishes filled with 2 mL cold HBSS and place them on ice.
1.3 mM tricaine (67.94 mg ethyl 3-aminobenzoate methane sulfonate salt +200 mL ddH2O)
Step 2: Heart excision
Anesthetize 20 fish simultaneously in a beaker filled with 200 mL anesthetic buffer for ∼3 min until vital signs are lost.
Individually, rinse fish over HBSS and 75% ethyl alcohol twice, and place euthanized zebrafish ventral side up on a moist surgical sponge under a dissecting stereomicroscope (Fig. 1C).
Remove skin from the chest with scissors, squeeze the hypogastrium to expose the heart, and quickly excise the heart from the chest (Fig. 1D; Supplementary Video S1).
Collect hearts into a 35 mm cell culture dish filled with cold HBSS buffer on ice immediately after being separated from the chest (Fig. 1F).
Remove bulb of aorta and redundant tissue from the ventricle and transfer the pruned ventricles to a new 35 mm dish filled with HBSS buffer (Fig. 1G).
Step 3: Enzymatic digestion and mechanical separation
Transfer ventricles to the vertical super clean bench, rinse them with cold HBSS three times, then transfer ventricles to a 60 mm sterilized dish and wash them with HBSS two times.
Add 6 mL digestive buffer into the 60 mm dish containing ventricles and cut them into pieces (Fig. 1H).
Transfer the mixture of heart pieces and digestive buffer into a 50 mL centrifuge tube. The heart fragments appear a deep red at this step (Fig. 1I).
Bathe the mixture at 32°C for 10 min and resuspend the pieces every 2 min by oscillating. After 10 min digestion, pieces turn pale (Fig. 1J).
Gently pipet the heart pieces with 1 mL tips to force mechanical separation in the sterilized bench (Fig. 1K).
Transfer supernatant to cold stopping buffer (10 mL culture medium).
Add the remaining 4 mL digestive buffer to the sediment and bathe the mixture at 32°C for 5 min (Fig. 1L). At this point, undigested pieces should be significantly less than that in Figure 1I.
Filter all gathered supernatants with 100 μm cell strainers, then centrifuge them at 1000 rpm for 5 min (Fig. 1M).
Resuspend cardiomyocyte sedimentation with fresh medium, then place it at 28°C, and 5% CO2 incubator to precipitate fibroblasts. After 2 h, transfer culture supernatant containing cardiomyocytes to a new dish or proper plates.
Method of culture
Immediate use: No coating required. The number of fish depends on the types of downstream experiments. For example, cardiomyocytes isolated from five fish hearts are enough for flow cytometry. Twenty fish can provide sufficient cells to plate four wells of a 24-well plate in high density for RNA and protein extraction (2.5 × 105 cells/well). Researchers can adjust regents and materials used according to the number of fish.
Long-term culture: Coat the culture plates or dishes with poly-
Formula of culture medium: There are no standardized recipes for the nutrient and growth factor medium that help cardiomyocytes grow into beating cells. We propose a method that worked for us, which consisted of 90% DMEM/F-12 (Ham), 10% FBS, 1% ITS, and 1% P/S.
Selection of coating agent: poly-
Plating density: Studies23,24 in adult and neonatal mouse cells suggest 1.5 × 104 cells per 35 mm dish is optimal. Higher plating density is not recommended, as it may lead to cell death due to cell-to-cell contact. 25 Densities could be adjusted based on the intended purpose. Low density (104 cells/mL) is better for cell immunofluorescence assay, while medium to high density (105–106 cells/mL) is better for extended culture and extraction of RNA and protein.
Culture condition: Zebrafish cardiomyocytes should be cultured at 28°C in a 5% CO2 incubator. It is recommended to change the medium two to three times a week and only half of the medium are replaced at a time to avoid cell detachment.
Quantitative real-time PCR
Total RNA was extracted using TRIzol reagent (Cat. No. 15596026; ThermoFisher) and reversed to cDNA using Fasting gDNA Dispelling RT SuperMix FastKing (Cat. No. KR118-02; TIANGEN) according to the manufacturer's protocol. Talent qPCR PreMix (SYBR Green) (Cat. No. FP209-02; TIANGEN) was used for real-time PCR on a Fluorescence ration PCR instrument (ABI 7500). 18S RNA was used as a housekeeping gene. Primer sequences are provided in Supplementary Table S1.
Immunocytochemistry
Cell monolayers were fixed in 4% paraformaldehyde for 30 min and permeabilized with 0.05% Triton-X 100 for 20 min. Antibodies were diluted according to the manufacturer's instructions. Nuclei were stained with DAPI (Cat. No. P0131; Beyotime). Primary antibodies tropomyosin and α-sarcomeric actin were incubated for 2 h at room temperature. Cells were visualized with an inverted fluorescence microscope (Zeiss—Axio Vert.A1) and 3D images using Laser Scanning Confocal Microscope (Nikon A1R HD25).
Flow cytometry
WT and transgenic zebrafish lines (Tg(cmlc2: EGFP), Tg(kdrl:mCherry)) were used to harvest cardiomyocytes. All procedures were performed according to the manufacturer's instructions. Single-cell suspension from zebrafish (Tg(cmlc2: EGFP) hearts was stained with PI (50 μg/mL) at 4°C. Events were acquired on a CytoFLEX flow cytometer (Beckman Coulter) and results were analyzed using FlowJo 10.4 software. Viable cardiomyocytes were identified as EGFP+/PI− cells, endothelial cells as mcherry+ cells.
Results
The yield and purity of isolated cardiomyocytes
In this study, isolated cardiomyocytes were obtained from 20 adult zebrafish, resulting in a density of ∼5 × 105 cell/mL, totaling 1 × 106 of viable cardiomyocytes.
The gene expression levels of different tissue-specific markers were measured by quantitative real-time PCR (qPCR) (Fig. 2A). Cmlc2a and tpm1 were used as molecular markers for cardiomyocytes, flia and kdrl for endocardium, wt1b and tcf21 for epicardium, and vim for cardiac fibroblasts. We found that cardiomyocyte-specific markers, cmlc2a and tpm1, can be detected in the cell suspension before and after purification, while other markers, including flia, kdrl, wt1b, tcf21, and vim, are almost undetectable in the purified cardiomyocytes.
Purified cells highly expressed cardiomyocyte-specific markers and flow cytometry analysis confirmed the purity of cardiomyocytes.
Cells were obtained from transgenic zebrafish Tg(cmlc2:EGFP) ventricles (Fig. 2B) and were used for flow cytometry analysis. Purified cells isolated from WT zebrafish hearts were used as the negative control (Fig. 2C). Flow cytometry results indicated that the isolated cells were mainly viable cardiomyocytes (EGFP+/PI− cells) (Fig. 2D–F). Furthermore, purified cells from transgenic zebrafish Tg(kdrl:mCherry), whose endothelial cells were labeled with red fluorescence, were used to verify if there was contamination of endothelial cells. Isolated cells from the aortic bulb and myocardium served as positive control for endothelial cells (Fig. 2G). As shown in Figure 2H, mCherry+ endothelial cells were barely detected in the purified cells, indicating a low level of contamination of endothelial cells.
The activity of isolated cardiomyocytes
3D morphology of cardiomyocytes
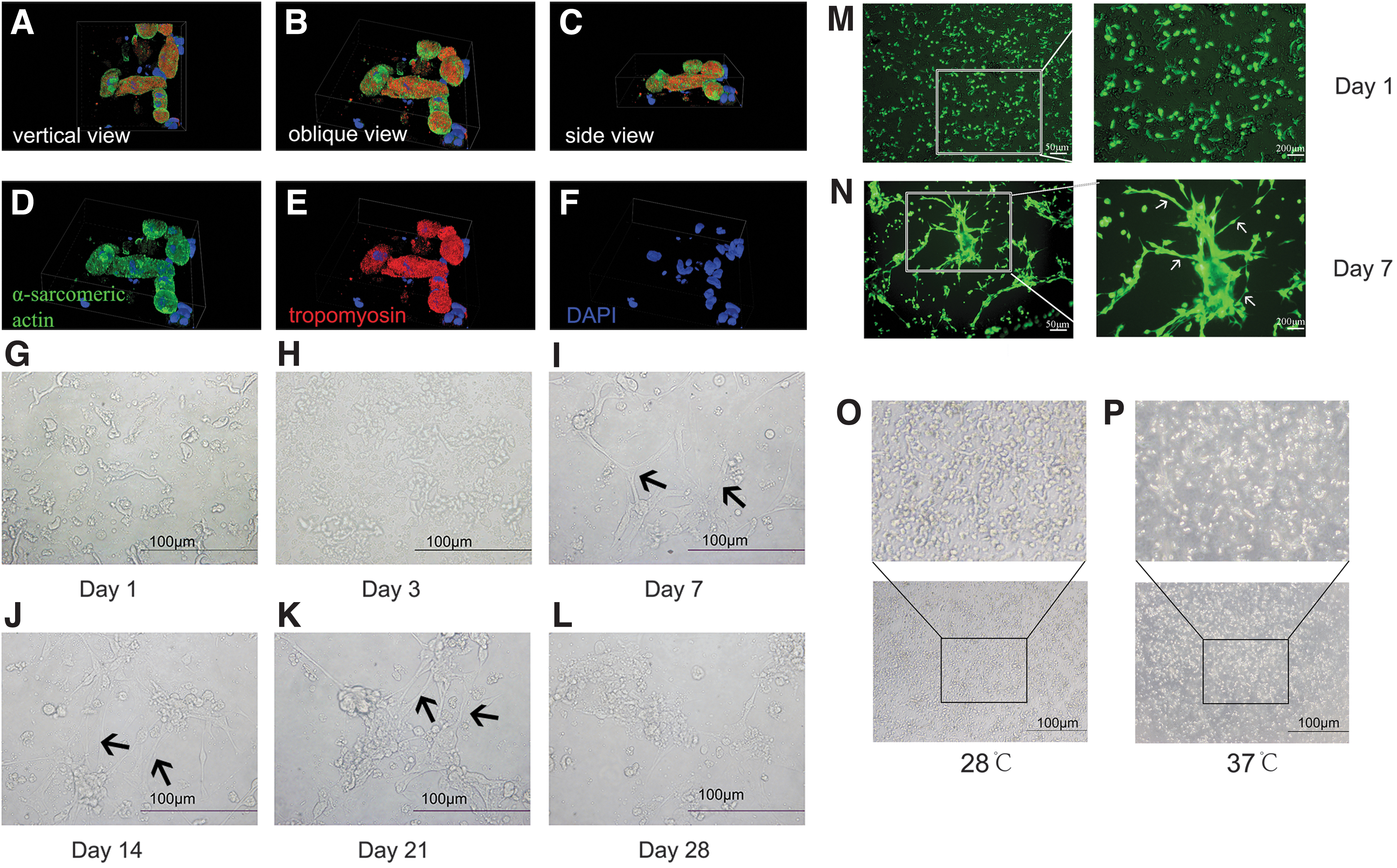
We labeled the isolated cardiomyocytes after plating for 24 h with antibodies targeting α-sarcomeric actin (green), tropomyosin (red), and DAPI (blue), and high-resolution 3D images were taken using a Laser Scanning Confocal Microscope and the morphology was depicted using deconvolution (Fig. 3; Supplementary Video S2-part1). The 3D morphology of cardiomyocytes at the initial culture stage was observed from vertical view (Fig. 3A), oblique view (Fig. 3B), and side view (Fig. 3C).
Morphology of in vitro cultured cardiomyocytes.
Dynamic changes in morphological characteristics of cardiomyocytes
From day 1 to 21 of in vitro culture, cardiomyocytes gradually transitioned to a firmly adherent state, with increased cytoplasmic volume and apparent intercellular connections, and could contract autonomously (Fig. 3G–K). On day 1 after plating, cardiomyocytes grew individually without intercellular connections (Fig. 3M), and a few of cells began to beat prematurely (Supplementary Video S2-part 2). Most cells started to contract on day 3 (Supplementary Video S2-part 3). After 7 days of plating, intercellular junctions appeared (Fig. 3N), and strong contractions were demonstrated in the spindle or rod-shaped cells (Supplementary Video S2-part 4 and part 5).
After 4 weeks, ∼30%–40% of cells were lost (Fig. 3L), but the remaining cardiomyocytes were able to form cell clusters, and contract strongly in sync (Supplementary Video 2-part 6).
We also investigated if zebrafish cardiomyocytes could grow at the conventional cell culture temperature of 37°C. Therefore, after cardiomyocytes adhered to the plastic Petri dishes, we transferred the dishes to a 5% CO2 incubator at 37°C for 24 h. As shown in Figure 3P, 37°C was not suitable for zebrafish cardiomyocyte growth.
Discussion
In the physiological anatomy of zebrafish, the heart is situated in the anterior part of the body cavity and ventral to the esophagus. 26 The adult zebrafish heart measures ∼1.5 mm in length 11 and weighs around 0.5 mg. Over 95% of adult zebrafish cardiomyocytes are diploid mononuclear cells,27,28 endowing them with a high regenerative capacity.29,30
Three hearts at a time are recommended in Veronika Sander's method, 20 while our method can handle the isolation of 20 or more hearts simultaneously. Grunow et al. 31 managed to isolate ∼60 μg of proteins from 20 larval zebrafish hearts. Kim et al. 22 suggested that around 5 × 104 viable cells per heart could be collected after tissue mincing and enzymatic digestion. Consistent with previous studies, our protocol was able to obtain 180–200 μg protein from 20 adult zebrafish hearts, equivalent to a yield of 1 × 106 cells, which should suffice for a variety of experiments (such as western blot, RT-qPCR, and flow cytometry).
A subset of cardiomyocytes isolated in our study began beating on day 1, earlier than in previous methods, 23 most started to contract on day 3–5, and achieved synchronized beating by day 8 postplating. The 3D images captured by Laser Scanning Confocal Microscope depict the initial morphology of the isolated cardiomyocytes, consistent with those reported in previous studies.
For the cardiomyocytes and endothelial cells account for the largest proportion in zebrafish heart, transgenic zebrafish Tg(cmlc2: EGFP) and Tg(kdrl:mCherry) are used to validate the purity of isolated cells through flow cytometry analysis, indicating that the majority of the isolated cells are cardiomyocytes. qPCR results also show that cardiomyocyte markers are highly expressed in the purified cells, while markers for cardiac fibroblasts, endocardial, and epicardial cells were almost undetectable.
Isolation
We have described the operational points in the section of Procedures of isolation. We tried to prepare a heart quickly within 10 s (Supplementary Video S1) to ensure the survival and good behavior of cardiomyocytes.
Digestion
Our protocol is primarily based on the enzymatic methodology of cardiomyocytes isolation from neonatal mice, 32 which is much simpler than that from adult rodents due to the omission of aorta cannulation and perfusion. Various enzyme digestion solutions (usually a mixture of collagenase and protease) are widely used for primary cell isolation. Enzyme selection is one of the key factors in the success of the isolation. Compared to the approach using high-concentration collagenase,16,20 we found that a lower concentration of a single enzyme (collagenase II, 0.5 mg/mL) is sufficient for digesting and separating zebrafish heart tissue.
Type II collagenase is recommended, for it is highly specific for insoluble collagen and gelatin and highly reactive against acid-soluble collagen and Pz-PLGPR.33,34 We only used 0.5 mg/mL type II collagenase, which is sufficient to digest the heart pieces, given its higher clostripain activity than other products 35 and its wide preference for primary cell isolation. 36
It is desirable to use a minimal amount of digestive enzymes as it minimizes damage to the cells. Our two-step enzyme digestion (0.5 mg/mL collagenase type II, 10 min for step 1 and 5 min for step 2) process is a simple, economical, and reproducible isolation technique that allows isolated adult cardiomyocytes to form stable cell colonies on cell culture dishes.
Stopping buffer
In contrast to Veronika Sander's method, our study did not use heparin buffer or different gradients of CaCl2 stopping buffer. Instead, we chose commercial Hank's Balanced Salt Solution to wash isolated hearts and serve as a solvent to dissolve collagenase, resulting in more viable cells at the correct osmotic pressure. Rather than using a variety of digestive enzymes in high concentrations to promote cell isolation from heart tissues, we increased the contact area of tissues with the digestive buffer by cutting the heart into small pieces and constant shaking in the 32°C water bath.
Purifying
Percoll gradients and centrifugation are widely used to purify cardiomyocytes isolated from rodents.37,38 This purification method is not applicable for zebrafish, whose cardiomyocytes are few and small, thus we opted for a differential velocity adherent approach. First, during the heart excision process (Procedures of isolation, Step 2), completely omitting the bulbus arteriosus and retaining only the ventricular tissue helps avoid contamination of nonmyocytes, especially endothelial cells. Two hours after plating, fibroblasts precipitated, and the supernatant containing cardiomyocytes was transferred to a new culture dish or plate for further culture. In this way, fibroblasts, which are the main confounding factor, could be efficiently removed.
Plating
Direct plating cells in a monolayer can lead to a lack of expansion and anchorage independence. Consequently, selecting the appropriate coating materials enhances the quality of cell culture. Morphological modifications (e.g., fibrous, or porous materials),
33
chemical modifications (e.g., glow discharge, grafting macromolecules), biological modifications (heparin, peptide sequence [arginine-glycine-aspartic acid], and growth factors and natural extracellular matrix proteins [collagen, laminin, fibronectin]) help avoid cardiomyocyte detachment. Moreover, commercially available synthetic polymers, including poly-
In our study, coating cell culture dishes or plates with 0.01% (w/v) Poly-
Some 3D culture methods and gene editing tools (e.g., modulation of Wnt signaling) have been used to promote hiPSC-CM maturation.13,15,30–43 The emergence of M-bands and cardiac-specific gene expressions in hiPSC-CMs has been reported by Kamakura et al. through 1-year cell culture. 44 Such extended culture periods limit its broad applicability for examining intracellular Ca2+ homeostasis, protein biochemistry, and gene manipulation (e.g., infection or transfection).13,15 In this study, we present an efficient and reproducible method for rapid isolation and in vitro culture of cardiomyocytes from adult zebrafish hearts, enabling the execution of experiments that mimic cardiomyocyte properties and functions in vivo. Furthermore, this method supports the application of zebrafish model organisms in cardiovascular disease research.
Conclusion
The culture of adult zebrafish cardiomyocytes as an important addition to the in vitro study models can provide new insights to elucidate the process of cardiomyocyte renewal and explore cellular and subcellular physiology. Our study isolated primary cardiomyocytes with high purity and activity using a relatively short time and low concentration of digestive enzymes, and was able to achieve long-term culture, adding an accessible tool to the field of zebrafish as a model organism for studying cardiac diseases.
Footnotes
Acknowledgments
The authors thank Professor Zu Yao from Shanghai Ocean University for the gift of transgenic zebrafish (Tg(cmlc2: EGFP) and Tg(kdrl:mCherry)).
Authors' Contributions
Conceptualization, original draft preparation: C.Z.; Substantively revise the article: Y.S.; Funding acquisition: Z.C. and Y.S.
Declarations
All the zebrafish were studied according to protocols approved by the Institutional Animal Care Committee of Ruijin Hospital in accordance with the Guide to the Care and Use of Experimental Animals and the Animals for Research Act.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China to Z.C. (81370331) and Y.S. (82200311).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.