
Abstract
Small nuclear DNA (snDNA) are valuable cytogenetic markers for comparative studies in chromosome evolution because different distribution patterns were found among species. Parodontidae, a Neotropical fish family, is known to have female heterogametic sex chromosome systems in some species. The U2 and U4 snDNA sites have been found to be involved in Z and W chromosome differentiation in Apareiodon sp., Apareiodon affinis, and Parodon hilarii. However, few studies have evaluated snDNA sites as propulsors of chromosome diversification among closely related fish species. In this study, we investigated the distribution of U2 and U4 snDNA clusters in the chromosomes of 10 populations/species belonging to Apareiodon and Parodon, aiming to identify chromosomal homeologies or diversification. In situ localization data revealed a submetacentric pair carrying the U2 snDNA site among the populations/species analyzed. Furthermore, all studied species demonstrated homeology in the location of U4 snDNA cluster in the proximal region of metacentric pair 1, besides an additional signal showing up with a divergence in Apareiodon. Comparative chromosomal mapping of U4 snDNA also helped to reinforce the proposal of the ZZ/ZW1W2 sex chromosome system origin in an A. affinis population. According to cytogenetic data, the study corroborates the diversification in Parodontidae paired species with uncertain taxonomy.
Introduction
Repetitive DNA constitutes the major fraction of the DNA content of many species.1,2 It is represented by the tandem repeats (satellites, minisatellites, and microsatellites), gene families (e.g., ribosomal DNAs, histones, and small nuclear RNAs [snRNAs]), and transposable elements.3,4 These repetitive sequences have been identified as a significant contributors to the observed karyotype variation among species.2,5
Gene families often originate through duplications from a common ancestral copy, resulting in similar DNA sequences and related functions. 6 Among these families, U snRNAs play a crucial role as components of the spliceosome, a complex machinery responsible for mRNA precursors splicing. 7 The major spliceosome, which is the dominant splicing machinery, is organized by multiple proteins and the U1, U2, U4, U5, and U6 snRNAs.8,9 These loci are considered valuables markers for evolutionary studies among closely related species, aiding in inferring homeology between chromosomes from different lineages, and tracking the origin and evolution of specific genomic regions, such as B and sex chromosomes. 10 However, comparative cytogenetic studies applying U small nuclear DNA (snDNA) probes are still scarce in teleosteans. 11
In general, studies have primarily focused on the in situ locations of U1 and U2 snDNAs, with the single-locus condition being present in most fish karyotypes.12–17 However, multiple chromosome sites18–21 or scattered signals across the chromosomes were also observed.22,23 Apart from U1 and U2 snDNAs, cytogenetic studies on U4, U5, and U6 snDNAs are scarce.21,24,25
Parodontidae is a Neotropical fish family with 32 valid species, 26 grouped into three genera: Parodon Valenciennes, 1849, Saccodon Kner, 1863, and Apareiodon Eigenmann, 1916. 27 The species presents a wide geographic distribution through South America and Panama, except in some coastal Atlantic basins and Patagonia. 28 Their representatives show a controversial taxonomy because most lack reliable diagnostic morphological traits to support accurate phylogenetic analysis. 28 Hence, controversies between the morphological and molecular species delimitation approach for some closely related species lead to taxonomy uncertainties or the nondescription of valid taxa in the scientific literature.29–34
Parodontidae species have a shared diploid chromosome number (2n) of 54. 35 Despite the 2n conservation, variations in karyotype formulae and number and location of repetitive DNAs were described in the Parodontidae karyotype evolution.21,32–39 The remarkable karyotypes feature in the family is the occurrence of species with no evidence of heteromorphic sex chromosomes, species presenting a proto-sex chromosome pair, and others with ZZ/ZW and ZZ/ZW1W2 sex chromosome systems.31–33,36,38–41
Recently, in addition to molecular characterization of the U snRNA genes, the U1, U2, U4, U5, and U6 snDNAs were mapped in the chromosomes of five Parodontidae species. 21 The U2 and U4 snDNAs sites have been localized in the Z and W chromosomes and proposed to act in sex chromosome differentiation events in some lineage-specific in Parodontidae. 21 Thus, this study aimed to investigate the karyotype distribution of the U2 and U4 snDNAs in different populations/species of closely related groups in Parodontidae.
Materials and Methods
Individuals from different populations/species of Apareiodon and Parodon sampled in different River basins in Brazil were analyzed (Table 1). Specimens were collected with the authorization of the Chico Mendes Institute for Biodiversity Conservation (ICMBIO), System of Authorization and Information about Biodiversity (SISBIO-License No. 15117-3), and National System of Genetic Resource Management and Associated Traditional Knowledge (SISGEN No. AE12D3D). The procedures were approved by the Ethics Committee of Animal Usage of the Universidade Estadual de Ponta Grossa (Process CEUA No. 6/2019) and Biosafety Certification according to Comissão Técnica Nacional de Biossegurança—CTNBio (CQB No. 0063/98).
Details of Parodontidae Species Analyzed in This Study: Location, Number of Males (♂) and Females (♀), and Identification
GPS, Global Positioning System; MNRJ, Museu Nacional do Rio de Janeiro; NUP, Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura.
Mitotic chromosomes were obtained from the anterior kidney by the air-drying method, according to Bertollo et al. 42 The U2 and U4 snDNA sequences from Apareiodon sp. (GenBank Ids MZ645212–MZ645213, respectively) were labeled by polymerase chain reaction using digoxigenin-11dUTP (Jena Bioscience, Jena, Germany), according to Azambuja et al. 21 Fluorescence in situ hybridization (FISH) was performed following Pinkel et al., 43 and it was conducted under stringency conditions as follows: 200 ng of each probe, 50% formamide, 10% dextran sulfate, and 2XSSC—saline-sodium citrate; 16 h of hybridization at 37°C. Signals were detected using anti-digoxigenin rhodamine Fab fragments (Roche Applied Science, Penzberg, Germany).
Chromosomes were counterstained with 0.2 μg/mL of DAPI in Vectashield mounting medium (Vector Laboratories, Burlingame), and chromosomal preparations were analyzed in an epifluorescence microscope (Leica DM 2000) coupled to a DFC3000 G CCD camera (Leica). Chromosomes were classified based on the arm ratio according to Levan et al. 44 and arranged in karyotypes as metacentric (m), submetacentric (sm), subtelocentric (st), and acrocentric (a).
Results
All populations/species analyzed presented 2n = 54, distinct karyotype formulas, and ZW sex chromosome system occurrence in Apareiodon ibitiensis and Apareiodon vladii (Table 2 and Fig. 1). FISH with the U2 snDNA probe demonstrated a single cluster located at the proximal region of short arm (p) of a submetacentric in all analyzed populations/species (Table 2 and Fig. 1). This submetacentric corresponds to the chromosome pair 4 in Apareiodon affinis populations (Table 2 and Fig. 1a–c), chromosome pair 3 in A. ibitiensis, Apareiodon vittatus, A. vladii, Parodon pongoensis, and Parodon sp. (Table 2 and Fig. 1d–g, i, j), and chromosome pair 2 in Parodon buckleyi (Table 2 and Fig. 1h).

Karyotypes of Apareiodon affinis populations (
Apareiodon and Parodon Cytogenetic Data and Location of U2 and U4 Small nuclear DNAs sites
2n, diploid chromosome number; a, acrocentric; m, metacentric; p, short arm; PS, present study; q, long arm; sm, submetacentric; snDNA, small nuclear DNA; st, subtelocentric; ♂, male; ♀, female.
U4 snDNA in situ localization showed two chromosomal sites on the metacentric 1, at proximal and terminal regions, in all Apareiodon species (Table 2 and Fig. 1a–g; boxes). In Parodon species, that is, P. buckleyi, P. pongoensis, and Parodon sp., a single locus at the proximal region of the metacentric 1 was observed (Table 2 and Fig. 1h–j; boxes).
Discussion
Parodontidae species present a conserved 2n and structural chromosome rearrangements as the main propulsors of karyotype evolution.21,31,34–36 Gene families mapped in the chromosomes of Parodontidae species, such as rDNAs, histones, and snDNAs, have shown few chromosomal remodeling events, most triggering specific events of differentiation in the lineage.21,35,36,45 In this study, the in situ comparative localization of U2 and U4 snDNA probes in more representatives showed a better diversification scenario between closely related species in Parodontidae.
Comparative studies using U2 snDNA probes in fish showed that it is usually located on a probable homeologous chromosome pair.10,13,17,46–50 However, multiple loci or scattered signals along all the chromosomes can also be observed.18,19,23 In this study, the data demonstrated that a single U2 snDNA locus in a probably homeologous chromosome site is the predominant condition in Apareiodon and Parodon species, also described in Parodon nasus. 21 Additional U2 snDNA sites were described in A. affinis (Upper Paraná River population), Apareiodon piracicabae, Apareiodon sp. (Verde river), and P. hilarii. 21 However, these additional sites acted in specific differentiation events in each lineage.
Studies with the U4 snDNA probes in fish are restricted to Hollandichthys multifasciatus, three Ancistrus species, and five species of Parodontidae.21,24,25 In situ localization of U4 snDNA probes showed a shared labeled site in the long arm (q) proximal region of chromosome 1 in all analyzed Parodontidae species. In addition, all Apareiodon species also showed a q terminal U4 snDNA site in chromosome 1. For now, the q terminal U4 snDNA site in the metacentric pair 1, specifically to Apareiodon, showed up as a consistent karyotype divergence compared with Parodon species.
The karyotypes in Parodontidae species are predominantly composed of bi-armed chromosomes.33,35,38,39,41 The exception occurs in A. affinis, which was considered a cryptic species group. 31 In A. affinis, despite 2n conservation, the Upper Paraná River population presented a multiple sex chromosome system (ZZ/ZW1W2), and others from Lower Paraná River showed a variation of 4–16 acrocentrics and no heteromorphic sex chromosomes.31,51–53 Distinct A. affinis representatives from Lower Paraná River populations showed homeologous chromosomes bearing U2 and U4 snDNA sites, despite the karyotype variation of 4, 10, and 16 acrocentrics to the Uruguay, Cuiabá, and Paraguay rivers populations, respectively. The U2 and U4 snDNA sites were also mapped in the karyotypes of A. affinis with a ZZ/ZW1W2 sex chromosome system, that is, from the Upper Paraná River population. 21 Comparing the ZZ/ZW1W2 karyotype with those without heteromorphic sex chromosomes, an additional U2 snDNA site can be found on st pair 27.
Furthermore, A. affinis from the Upper Paraná River showed U4 snDNA signals on Z, W1, and W2 chromosomes, whereas for those without heteromorphic sex chromosomes, this cluster is restricted to m pair 1. These data add one more chromosome difference to support the proposal of the cryptic species group for A. affinis among Upper Paraná, Cuiabá, Paraguay, and Uruguay rivers populations 31 although the morphological analyses have not demonstrated consistent differences among their representatives. 27
Based on in situ localization of the repetitive DNAs, a mechanism for the origin of the ZZ/ZW1W2 sex chromosome system in A. affinis has been suggested. 36 According to Schemberger et al., 36 in the first stage, a reciprocal translocation occurred between an autosomal pair and a proto-sex chromosome pair (Fig. 2). After that, the W chromosome generated by reciprocal translocation undergoes centric fission followed by pericentric inversions, originating the W1 and W2 sex chromosomes.
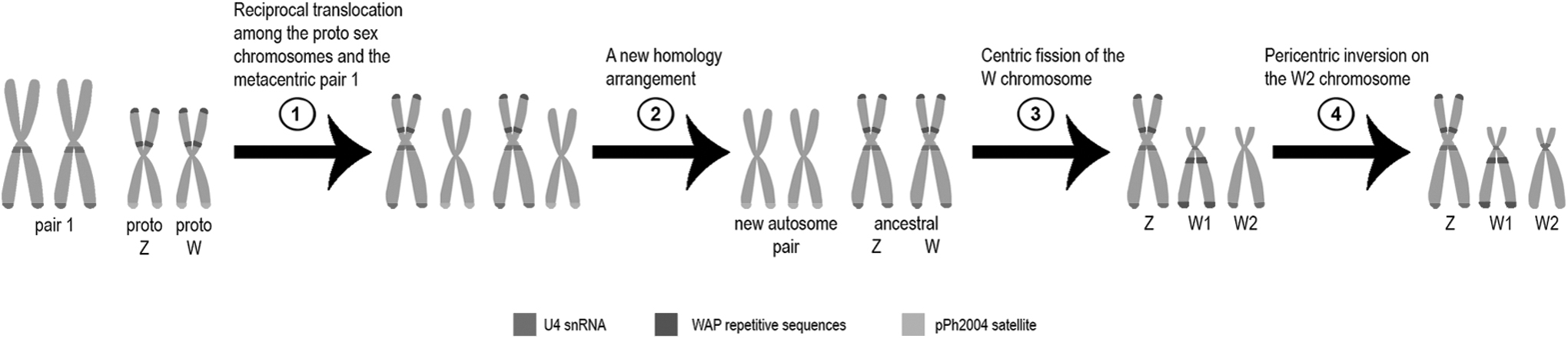
Updated proposal of ZZ/ZW1W2 sex chromosome system origin in A. affinis from the Upper Paraná River population, by Schemberger et al. 36 The image presents the location of U4 snDNA sites, and based on chromosome homeologies, we suggested the metacentric pair 1 involved in the reciprocal translocation with a proto-sex chromosome in the first stage of sex chromosome system differentiation.
In this study, the data demonstrated that the U4 snDNA sites are shared in the metacentric 1q from the Uruguay, Cuiabá, and Paraguay rivers A. affinis populations with the Z chromosome from the Upper Paraná River. In addition, according to Azambuja et al., 21 the pericentromeric U4 snDNA sites in the W1 and W2 chromosomes are owing to repositioning generated by pericentric inversions (Fig. 2). Thus, the homeology of the 1q in all Apareiodon species reinforces its involvement in the origin of the multiple sex chromosome system in populations of A. affinis from Upper Paraná River. In this proposal, the 1p of the Z chromosome in A. affinis corresponds to a large portion of proto-sex chromosome involved in reciprocal translocation.
Morphological and cytogenetic analyses demonstrated that A. ibitiensis, A. vladii, and Apareiodon sp. (Verde River) arrange a closely related group in Apareiodon, all of them possessing a ZZ/ZW sex chromosome system, and some doubts regarding Apareiodon sp. as a valid species.29,30,54 Despite the shared U2 and U4 snDNA sites in the karyotypes, Apareiodon sp. has an additional U2 snDNA site in ZW sex chromosomes. Taken together with other chromosome differences shown in Apareiodon sp.,36,40 the snDNAs sites localization corroborates its diversification from the A. ibitiensis lineage. Apareiodon piracicabae and A. vittatus also arrange a paired species group in Apareiodon, 27 neither of both showing evidence of heteromorphic sex chromosomes, 36 and low rates of genetic divergence.32,55 However, an extra U2 and U4 snDNA sites in A. piracicabae 21 and other chromosomal differences 36 reinforce the karyotype diversification between A. piracicabae and A. vittatus.
Among the Parodon analyzed species, the U2 and U4 snDNA sites conserved localization into karyotypes is more accentuated than the Apareiodon group. Parodon buckleyi, P. pongoensis, and Parodon sp. (Peixe river) shared the U2 and U4 snDNA chromosomal sites with P. nasus, analyzed by Azambuja et al. 21 In common, species from “P. nasus group” possess a proto-sex chromosome pair and few chromosome divergences. 36 The exceptions are described in P. hilarii, which has a differentiated ZW sex chromosome system and extra U2 and U4 snDNA sites in its karyotype. 21 In addition to Parodontidae species, snDNA signals on sex chromosomes have been described in Gymnotus pantanal and Triportheus albus.10,20 In all lineages, the snDNA sequences participated in specific events of sex chromosome differentiation.
Previous studies in Parodontidae have demonstrated the role of repetitive DNAs in chromosomal diversification and specific events of sex chromosome differentiation. 40 Here, the in situ U2 and U4 snDNA comparative localization showed that these gene families are located predominantly in homeologous chromosomes in Parodontidae species, with few karyotypes remodeling events. Still, the cytogenetic comparison detected a karyotype divergence for U4 snDNA distribution between Apareiodon and Parodon and some species-specific events of differentiation involving snDNA sequences, mainly to heteromorphic sex chromosomes.
Footnotes
Acknowledgments
The authors thank ICMBio (Instituto Chico Mendes de Conservação da Biodiversidade) for authorizing the collection of samples. The authors also thank Liano Centofante, Vladimir P. Margarido, Ezequiel A. de Oliveira, Francisco M. C. Sassi, Luis Henrique da Silva, and Antônio Donizete da Silva for collecting some specimens. This study is part of doctoral thesis of Matheus Azambuja in Genetics at the Universidade Federal do Paraná (CAPES scholarship—Finance code 001).
Authors' Contributions
M.A., V.N., O.M.-F., and M.R.V. conceived the project ideas, performed the experiments, analyzed data, and wrote the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Fundação Araucária (Fundação Araucária de Apoio ao Desenvolvimento Científico e Tecnológico do Estado do Paraná, Grant No.: 9/2017), FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo, Grant No.: 2015/16661-1), CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Finance Code 001) and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico, Grant No.: 305142/2019-4).