
Abstract
The zebrafish is a powerful model organism for studying development and regeneration. However, there is a lack of a standardized reference diet for developmental and regeneration experiments. Most studies evaluate the rate of growth, survival, and fecundity. In this study, we compare three diets and their effects on growth and regeneration after a spinal cord injury (SCI). Fish were fed daily for 1 week with daily measurements of overall length and width of spinal injury. Fish fed a live rotifer diet grew 32%, whereas a commercially available diet only led to a 4% increase in body length. Similarly, differences in rate of regeneration were observed with over 80% of rotifer-fed larvae forming a glial bridge after injury compared to <10% of zebrafish fed with the commercial diet. Our data highlight the need for establishing a standardized diet for regeneration studies to improve research reproducibility.
Introduction
Zebrafish are a powerful model organism for studying development and regeneration. One of the limitations with the use of zebrafish as a model is the lack of husbandry standardization, particularly for larvae, which are extensively used for developmental and regenerative studies. Diet and nutrition are variables that have potentially profound effects in multiple animal models,1–4 highlighting the need to control for diet within experiments.
Diet standardization in zebrafish laboratories is often an amalgam of research-based guidelines and anecdotal observations. Many different diets are available for use in zebrafish, including live feeds such as paramecia, rotifers, and artemia,5–7 or commercially formulated diets like powders and flakes.8,9 Although formulated diets offer some level of control over the nutritional content, the nutritional content of live feeds is dependent on what the live feed is fed, ultimately affecting what is available to larvae.10,11 In addition, the age of live feed can also affect the nutritional content available to larvae. 12 As such, laboratories are working toward the development of diets containing consistent ingredients and nutrients, referred to as standardized reference diets.13,14
Diet impacts many aspects of zebrafish physiology, including adult size and weight, 15 cardiac structures,16,17 and behavior, 18 as well as reproductive performance.19–21 Diet has been found to play a significant role in growth up to 15 weeks of age, with an early rapid growth period.22,23 Recent studies quantifying skeletal abnormalities up to 20 days post-fertilization (dpf) found dry feed increased caudal-peduncle scoliosis and gill-cover wrinkling. 11 In addition, providing high amounts of food lead to a faster time to maturation at the cost of a shortened window for embryo production. 24 However, larval growth at younger ages is not as well quantified.
Despite the numerous options for rearing larval zebrafish, there is a dearth of studies to determine whether different feeds may affect experiments, particularly when using larval zebrafish. We sought to determine whether diet affected early larval development and regeneration after a spinal cord injury (SCI). Larval zebrafish are particularly well suited to both neurodevelopment and neural injury models. 25 The spinal circuitry of larval zebrafish is well characterized and the larvae's transparency permits live imaging in real time of the recovery process. We tested two diets and a nutritive media on the growth and regeneration of larvae (Fig. 1). Total body length and free swim behavior were used to determine whether these conditions affected larval development. The locomotor activity and presence of a glial bridge were used to study the effect on regeneration after an SCI.

Schematic of experimental design. For developmental studies, larvae were raised until 5 dpf, and then given one of three diets. Larvae were recorded daily for length and swim behavior. For transection experiments, larvae were either transected at 5 dpf and recovered in different diets or were fed one diet from 5 to 6 dpf, were transected at 7 dpf, and resumed their assigned diet at 8 dpf. All larvae were recorded daily for length, glial bridge formation, and swim behavior. dpf, days postfertilization.
Materials and Methods
Zebrafish husbandry
Adult zebrafish [wild-type AB strain and Tg(GFAP:EGFP)] 3–18 months of age were obtained from the Zebrafish International Resource Center (ZIRC, Eugene, OR). Adults were maintained at 28.5°C in a recirculating system (Iwaki Aquatic, Holliston, MA) on a 14-h light/10-h dark cycle and fed in the morning with Ziegler's adult zebrafish diet (Pentair Aquatic Eco-Systems, Apopka, FL) and in the afternoon with brine shrimp (E-Z Egg; Brine Shrimp Direct, Ogden, UT). Water quality of system water is reported in Supplementary Tables S1–S2. Embryos were obtained through natural outcross matings. Embryos were cultured at 28°C–30°C in E3 medium containing 5 mM NaCl, 0.17 mM KCl, 0.33 mM CaCl2, and 0.33 mM MgSO4. All chemicals were purchased from Sigma Aldrich (St. Louis, MO). 26 Embryos were staged as described previously. 27 All animal procedures were approved by the IACUC committee at the University of Wyoming.
For all zebrafish experiments, at least two breeding tanks, each containing 2 to 4 males and 2 to 4 females from separate stocks, were set up to generate embryos. Embryos from each tank were randomly distributed across tested conditions. Unfertilized and developmentally abnormal embryos were removed before experiments. Zebrafish larvae began receiving food at 5 dpf, unless otherwise noted.
Larvae diet manipulations
Rotifers (Brachionus plicatilis; L-type rotifer) were maintained as a continuous culture at a density of 50–200 rotifers/mL in 15 ppt marine salt (Reed Mariculture, Campbell, CA). Rotifers were fed morning and afternoon with RGComplete (Reed Mariculture). RGComplete contains a minimum of 20% microalgae with a guaranteed analysis, including 2.7% crude protein (min), 0.9% crude fat (min), 0.2% crude fiber (max), 89% moisture (max), and total high unsaturated fatty acids 64.0 mg/g d.w. (min). Rotifers were harvested fresh for each day of the experiment and the salt was diluted to 5 ppt. Two milliliters of the diluted culture was then added to the buffer such that each dish had ∼100 rotifers/mL of buffer.
Ziegler's AP100 larval diet (Pentair Aquatic Eco-Systems) was added directly to the buffer to a final concentration of 0.1 mg/1 mL. Guaranteed analysis of the AP100 larval diet includes 50% crude protein (min), 12% crude fat (min), 2.5% crude fiber (max), 10% moisture (max), 15% ash (max), and 1.3% phosphorous. Buffer was replaced daily and supplemented with fresh feed. Given the high level of growth in media, the buffer was supplemented with 0.00003% methylene blue (Sigma #M9140). 26
Nutritive media developed for mouthless Xenopus tadpoles were used as an alternative to commercial and live feeds.28,29 The final working buffer was 9.5% Ham's Nutrient Mixture F12 (Sigma #51651C), 0.5% calf serum (Sigma #C8056) in buffer supplemented with methylene blue.
Water quality
For all diets, zebrafish were fed for 5 days. After experiments were completed, the solutions were filtered and sent to the Analytical Services Laboratory at the Wyoming Department of Agriculture. System water for the adult zebrafish housing system was tested at the same time. Results are reported in Supplementary Tables S1 and S2.
Zebrafish spinal transection
Larvae were transferred to E2 buffer 30 supplemented with penicillin/streptomycin (Pen/Strep; Gibco #15140–122, Grand Island, NY) on the day of transection. Larvae were mounted on flat slides in 2% low melting point (LMP) agarose (Goldbio #A-204-100, St. Louis, MO). The spinal cord was transected with a microblade (WPI #501731, Holliston, MA) at the level of the cloaca. Larvae were allowed to recover for ∼2 h in Petri dishes filled with E2 buffer supplemented with Pen/Strep before mounting and imaging. E2 buffer was replaced daily. Larvae were not fed on the day of transection.
Imaging, larval measurement, and confirmation of a glial bridge
Live larvae were mounted flat on a glass slide using 1.5% LMP agarose, and then anaesthetized using Syncaine/Tricane-S (MS-222; Syndel, Ferndale, WA). Larvae were imaged on an Olympus SZX16 stereoscope equipped with an Olympus DP80 camera and SDF PLANO 1XPF objective.
The preinstalled CellSens software (Olympus Life Science, Tokyo, Japan) was used to capture images and saved as TIFFs. Survival was noted on a daily basis. TIFFs were opened in FIJI open source software. 31 To measure body length, the segmented line tool was used. The start of the line was at the furthest rostral point of the larvae, the segment was placed just above the cloaca, and the terminal end of the line was placed on the end of the pigmentation at the caudal side of the larvae. To determine whether zebrafish recovered from SCI, images were opened in FIJI and the presence of a glial bridge was confirmed visually by two individual researchers. Zebrafish with continuous GFP signal across the injury site were considered fully bridged.
Locomotion behavior
Larvae were transected as outlined above and their behavior was recorded for five days. Behavior experiments were executed between 11:00 and 13:00 local time every day to ensure reproducibility. A clear 24-well plate (Celltreat #229123, Pepperell, MA) with 1 mL fresh E2 supplemented with Pen/Strep was used for the experiments, with one larvae per well. Larvae were fed respective diets, as outlined above. Locomotion was assessed using ZebraBox and its ViewPoint LS Tracking Software (v5.15.0.230; ViewPoint Life Sciences, Lyon, France). No stimulus was applied, and behavior was video recorded and tracked every 10 min for 60 min. Total swim distance was found by adding the tracked distances together (twitch: 0–2 mm/s, small: 2–4 mm/s, and large: >4 mm/s). Following the last day of behavior experiments, larvae were imaged as outlined above to record the absence or presence of a glial bridge.
Statistics
All larvae were included in experiments until the day of death. Graphs were plotted using GraphPad Prism (v9.3.0; Dotmatics, Boston, MA). Values for individual fish are plotted, and data are presented as the mean ± standard deviation (SD). For statistical testing, each distribution was assessed using the Shapiro–Wilk test to determine normality. Significant differences were determined using either an unpaired Mann–Whitney t-test or a Kruskal–Wallis ANOVA test with a post hoc Dunn's test. p-values of <0.05 were considered statistically significant.
Results
Rotifer diet promotes larval growth
To determine the effects a rotifer diet had on early larval development, we harvested rotifers and fed larvae from five dpf. Representative images of larval development from 5 to 9 dpf show larvae grew in length and width. (Fig. 2A). Larvae fed with rotifers for 5 days demonstrated a 32% increase in length. (Fig. 2B) To determine whether rotifers affected larval growth and regeneration after SCI, we next transected the transgenic line Tg(gfap:EGFP). This line labels radial glial, which form a proregenerative glial bridge after SCI.32–37

Rotifer-fed larvae have continued growth and regeneration after SCI.
After transection, zebrafish exhibited limited growth (Fig. 2C, D) with only a 16% increase in body length during recovery, suggesting that resources are prioritized toward regeneration over growth after an injury in larval zebrafish. Consistent with previous studies,32,37,38 larval zebrafish that survived the length of the experiment began the formation of a glial bridge 2–3 days postinjury (dpi), with ∼82% of larvae forming a glial bridge at 5 dpi (Table 1). As growth after transection was less than that seen during normal development, we next tracked survival (Fig. 2E). Ninety-seven percent of developing larvae survived on a rotifer diet, with 84% of larvae surviving after spinal cord transection. In summary, rotifers sustained high levels of growth and regeneration after SCI.
Percentage of Larvae Forming a Glial Bridge After Spinal Cord Transection at 5 Days Postfertilization
By 5 dpf, the nutrition from the yolk sac of zebrafish larvae is mostly exhausted, and larvae begin to rely on exogenous feed sources. 38 We wondered whether larvae injured after yolk sac depletion would be similarly affected by diet. For these experiments, we fed larvae 5–6 dpf, transected fish at 7 dpf, and resumed feeding at 8 dpf. Larvae grew in length until the day of SCI; however, after transaction, zebrafish larvae showed a small, but statistically significant decrease in length, with no additional growth (Supplementary Fig. S1A, B).
Our data suggest that after an injury, resources are dedicated to zebrafish regeneration and not developmental growth. We also noticed a significant decline in survival after SCI in larvae injured at 7 dpf and fed a rotifer diet (Supplementary Fig. S1C). Of the larvae that did survive, 55% of them were able to form a glial bridge (Table 2). Given the increase in death among injured larvae (<50% survival), we hypothesized that injured fish may struggle to catch the rotifers and sought to increase survival using non-live feeds.
Percentage of Larvae Forming a Glial Bridge After Spinal Cord Transection at 7 Days Postfertilization
NA, not applicable.
Powder diet does not promote regeneration from SCI
We first tested Ziegler's AP100 larval diet as an alternative to live feed. Figure 3A displays representative images of uninjured larvae from 5 to 9 dpf. In contrast to rotifer-fed larvae, zebrafish given the powder diet only grew 4% over the course of the experiment (Fig. 3B). We next transected larvae and found that the powder diet did not sustain continued growth during regeneration, with an overall 7% decrease in length (Fig. 3C, D). Moreover, few larvae were able to form a glial bridge (Table 1). Although larvae without injury were found to have a 100% survival over the course of the experiment, injured larvae only had a 60% survival (Fig. 3E).

Larvae fed with Zeigler AP100 diet only show continued growth without an SCI.
We also tested the effects of Ziegler's AP100 on the older larvae transection model (Supplementary Fig. S2A). Similar to rotifer-fed larvae, powder-fed zebrafish grew until the day of injury. However, after SCI, powder-fed larvae demonstrated an overall decrease in length by 1.6% (Supplementary Fig. S2B). Injured larvae given this diet had reduced survival compared to both uninjured and rotifer-fed SCI larvae. (Supplementary Fig. S2C). Similar to younger larvae, only 13% of larvae were able to form a glial bridge. As this diet could support development, these data suggest that the resources required for regeneration are different from those required for development.
Nutritive media does not encourage growth, but permit regeneration after SCI
We last tried nutritive media, as injured fish would not have to seek for or chase food. Representative images of larvae given this diet from 5 to 9 dpf are shown (Fig. 4A). Uninjured larvae did not grow and saw a decrease in length by 0.5% (Fig. 4B). We next transected larvae and found that the nutritive media did not sustain continued growth (Fig. 4C, D).

Larvae given nutritive media do not exhibit continued growth.
We observed a decrease in survival for both uninjured and transected larvae (Fig. 4E); however, of the transected larvae that survived, 82% of them were able to form a glial bridge by 5 dpi (Table 1), again suggesting that the nutrients required for larval development, survival, and regeneration from SCI are different. This was particularly evident when we transected older larvae (Supplementary Fig. S3). Larvae transected at 7 dpf and fed nutritive media decreased in length by 9.3% (Supplementary Fig. S3B). Nutritive media were unable to support larvae survival as a 100% mortality rate was observed by 5 dpi in both control and injured fish (Supplementary Fig. S3C). Surprisingly, 39% of larvae were able to form a glial bridge before death (Table 2).
Diet affects locomotion in developing and injured larvae
Zebrafish larvae begin freely swimming around 4 dpf. Recent studies have reported changes in swim behavior based on diet39,40; therefore, we chose to look at swimming behavior in uninjured and transected zebrafish larvae. We quantified total swim distances for larvae, given each of the diets (Fig. 5).
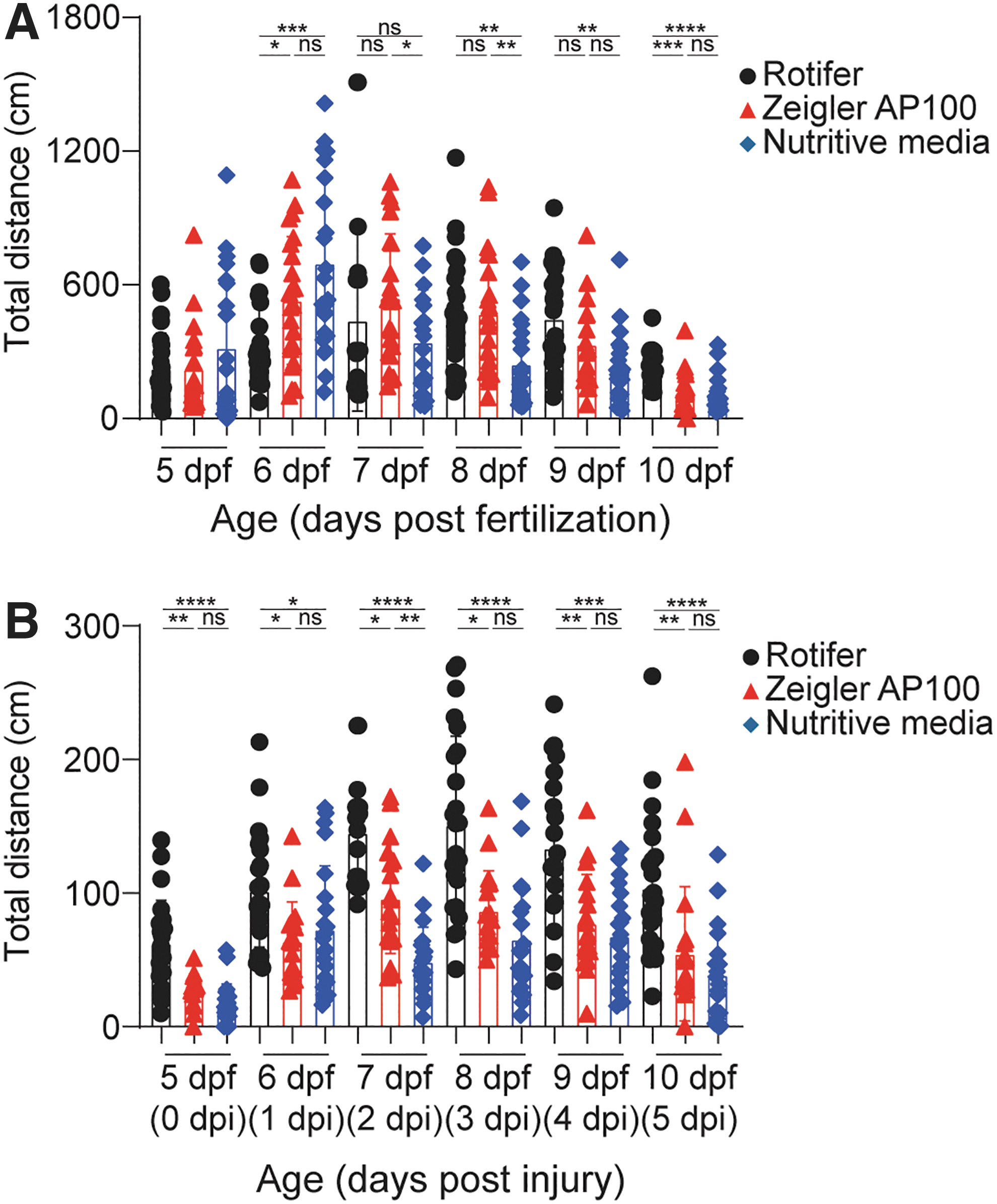
Larvae show differences in total swim distance based on diet.
In uninjured larvae, overall swim distances decreased as the zebrafish aged with all diets (Fig. 5A). After spinal cord transection, we found significant differences in swim distances based on diet. Surprisingly, there was a significant difference in total swim distances as early as the day of injury (Fig. 5B). On the day of injury (0 dpi), larvae given the rotifer diet had 143% more swim distance than those given the powder diet, and 257% more than those given the liquid diet. This trend remained throughout the trial, and by 5 dpi, the larvae fed a rotifer diet had total swim distances that were 118% more than those given the powder diet, and 207% more than those given the liquid diet (Fig. 5B).
Discussion
Diet plays an important role in zebrafish health and reproduction. Despite this, our understanding of how diet affects larval zebrafish experiments remains elusive. To that end, we systematically looked at the role different diets play in development and regeneration after spinal cord transection. Our results demonstrate that different feeds affect both growth and the ability of larval zebrafish to regenerate from an SCI.
We routinely feed our larvae rotifers to raise lines to adulthood due to the increased survival rate reported.5,41 In addition, rotifers are an ideal prey for developing zebrafish larvae because they are smaller than Artemia, move slowly through the water column, and promote development of prey capture skills. Because we culture rotifers in nutrient-rich microalgae, supplemented with highly unsaturated fatty acids, it is perhaps unsurprising that larvae fed a rotifer diet grow well.
One unexpected finding was that rotifer-fed larvae and larvae bathed in nutritive media regenerated well after SCI; however, larvae fed the Zeigler larval diet poorly regenerated. A major difference between the rotifer and AP100 diet is the minimum guaranteed crude protein content (2.7% vs. 50%, respectively). Previous work in zebrafish has reported dietary protein requirements for maximum weight gain (38%) and protein retention (45%). 42 Work in other fish species demonstrates that when too much protein is available, the excess is used for energy.43,44
Given the high percentage of protein in the Zeigler larval diet, it is possible that larvae metabolize the protein instead of using it toward tissue regeneration. Alternatively, studies in mammals show that the quantity and source of dietary protein impact microbiota function.45,46 It is possible that similar to mammals,47–49 bacterial imbalances can exacerbate spinal cord damage and/or impair recovery in zebrafish. The zebrafish community has multiple experimental paradigms to tackle this important question such as generating genetic mutants in germ-free zebrafish, transplanting exogenous bacteria, and environmental manipulation. 50
Experimental studies looking at swim behavior in zebrafish have used a range of feeds, including paramecia, 51 Artemia, 52 and Zeigler AP100 diet, 53 demonstrating the wide variety of feeding protocols used during behavioral experiments. Our study helps bridge the gap in knowledge by studying multiple diets in an experimental context. We found that after an SCI, rotifer-fed larvae had increased swim distances, even with complete spinal cord transection.
One explanation for the observed differences could be explained by an increase in available energy provided from the rotifers. Comparing free swimming feeds could provide insight into these differences. Another explanation is that the swimming behavior of rotifers provides an additional stimulus or alternative circuitry to promote free swim behavior. For example, ablation of brainstem projection neurons, which integrate visual inputs and project throughout the spinal cord to control turning movements, leads to an increase in forward swimming. 54 Both imaging and optogenetic technologies on injured animals can determine whether a visual stimulus can increase free swim through alternative circuits.
In conclusion, this study highlights that the choice of feed for zebrafish larvae can affect experimental results. Overall, these results have implications for the reproducibility of results across laboratories and highlight the need for publications to provide information on the feed used on experimental animals.
Footnotes
Acknowledgments
We thank Thomas Rynes for excellent fish care. We gratefully acknowledge support from the Wyoming Research Scholars Program (E.J.P.) and NIH P20GM121310 (K.M.).
Authors' Contributions
E.J.P. joined the Mruk laboratory in Fall 2020 and received her Bachelors of Science from the University of Wyoming in May 2023. At UW, she joined the Wyoming Research Scholars Program, which offered her a grant to pursue undergraduate research within the Mruk laboratory, studying spinal cord injury in zebrafish. E.J.P. completed the literature review, performed all experiments, and wrote the majority of this article. E.J.P. will be continuing her education at Colorado State University, where she has been accepted into the Doctor of Veterinary Medicine program.
K.M. is currently an Assistant Professor. The goal of the Mruk laboratory is to understand mechanisms by which zebrafish recover their behavior after a spinal cord injury. K.M. helped Emily design and analyze her experiments and provided edits to the figures and text of the article.
Disclosure Statement
The authors declare no competing financial interests.
Funding Information
This study was supported by NIGMS P20GM121310–05 to K.M.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.