
Abstract
Various methods have been used in rodents to evaluate learning and memory. Although much less frequently used, the zebrafish emerges as an alternative model organism in this context. For example, it allows assessing potential behavioral deficits because of neurodevelopmental disorders or environmental neurotoxins. A variety of learning tasks have been employed in previous studies that required extensive habituation and training sessions. Here, we introduce a simpler and faster method to evaluate learning and memory of zebrafish with minimum habituation. A new apparatus, a transparent L-shaped tube, was developed in which we trained each zebrafish to swim through a long arm and measured the time to swim through this arm. We demonstrate that in this task, zebrafish could acquire both short-term (1 h) and long-term memory (4 days). We also studied learning and memory of a gene knockout (KO) zebrafish that showed social impairments related to autism. We found KO mutant zebrafish to show a quantitative impairment in habituation, learning, and memory performance compared with wild-type control fish. In conclusion, we established a novel learning apparatus and sensitive paradigm that allowed us to evaluate learning and memory of adult zebrafish that required only a brief habituation period and minimal training.
Introduction
In the past two decades, zebrafish have been widely used to model a variety of CNS disorders, and several of these studies employed quantification of behavior to assess functional changes in the brain. 1 Many of these studies used learning paradigms, as numerous brain disorders are associated with memory impairment and/or attention deficits. 2 With the development of genome editing techniques in zebrafish, there has been an increased need for evaluating zebrafish models of rare neurological disorders.3–5 Various paradigms have been introduced to assess behavioral changes in zebrafish, including social behavior, anxiety, choice behavior, and learning and memory.6–10 For studying learning and memory, different types of mazes have been used.11,12 Although conventional and novel methods have been developed to assess learning and memory in zebrafish, in most of these paradigms, extensive habituation and training are required. 13 For example, numerous previous zebrafish learning studies using mazes involved long habituation periods over several days to acclimatize zebrafish to the apparatus and procedure. 14 In addition, several training trials had to be performed to achieve good memory performance, that is, the acquisition of conditioned stimulus–unconditioned stimulus (CS-US) association. 15 The issue with extensive habituation and large number of training trials is not just that these procedures take a long time, but also that they involve extensive human handling. Human handling induces stress, anxiety, and fear in most laboratory organisms, including the mouse and the rat. 16 Human handling maybe particularly damaging in zebrafish,17,18 but domesticated zebrafish may be more adapted to handling stressors. 19 Therefore, minimizing the need for extensive habituation and reducing human handling are crucial for studying zebrafish models of rare neurological disorders.
To minimize these issues, here we propose a simple and fast method to assess learning and memory of zebrafish. Our method does not require extensive habituation procedures, and it can achieve significant acquisition of memory faster than most previously published methods developed or adopted for the zebrafish.10,20 We substantiate this claim by showing how control (wild-type) zebrafish behave in this novel task. We also show that this task is capable of quantifying mutation induced impairment in learning and memory. Our findings indicate that training wild-type adult zebrafish in our new apparatus leads to acquisition of memory within five trials and that the memory is retained both for the short run (1 h) and the long run (4 days). In validating this behavioral apparatus, we employed a zebrafish model with dyrk1aa deficiency, a rare neurological disorder that demonstrates anxiolytic behavior and social impairment 21 Previous findings have reported that mutations in DYRK1A are associated with microcephaly, intellectual disability, or autism in humans in addition to severe learning and memory deficits in mice. 22 The dyrk1aa homozygous knockout (KO) zebrafish exhibit quantitative impairment in their memory performance compared with wild-type zebrafish. We conclude that our novel apparatus and procedure offer a simple paradigm with which researchers may be able to evaluate learning and memory and its impairment in adult zebrafish without the need to perform extensive habituation procedures or conduct a large number of training trials.
Materials and Methods
Animals and housing
Closed wild-type and dyrk1aa KO fish, developed previously, 21 were maintained at the Zebrafish Center for Disease Modeling (Daejeon, Korea). Adult fish were reared and maintained under standard conditions. 23 Briefly, recirculating system water was filtered through a multistage filtration system. Water quality parameters, such as pH (7.2) and conductivity (1300 µs), were maintained at constant levels. Temperature was maintained at 28.5°C. Light cycle was also controlled with 14 h light and 10 h dark. All adult fish were fed three times a day with freshly hatched brine shrimp. Conditions for all behavioral assays were identical. All fish used in this experiment were fully grown, sexually mature, 3- to 12-month-old fish, ranging in size between 3.0 and 3.5 cm standard length. Experiments were conducted according to guidelines approved by the Animal Ethics Committee of Chungnam National University (202012A-CNU-170).
Tube assay: learning and memory test
The test was designed to investigate learning and memory of zebrafish by measuring the time to swim through the long arm of an L-shaped tube, which was modified from a patent application (No. 10-2019-0132718). The tube was made of 1 mm thick transparent acrylic. Its long arm was 70 cm long, and its short arm was 10 cm long. The outer diameter of the tube was originally 2.5 cm. The tube was placed in a tank filled with system water, and all air bubbles were removed from inside the tube. The water column was kept at 5 cm (Fig. 1A). Water temperature was maintained at 28°C throughout the experiments. An overhead camera (Sony, HDRCX190, Japan) was used to record the behavior of fish. Also, in one experiment, to examine the effect of colors on this apparatus, two-thirds of the tube were covered externally with opaque-colored sheets (blue or yellow). Colored sheets were attached in a way that corresponded to the color on the bottom of the horizontally positioned long tube.

Experimental design of the tube assay and overview of zebrafish training.
Protocol
Each wild-type zebrafish was netted from their home tank and transferred to a 100 mL beaker containing 50 mL system water and then carefully introduced into the short arm of the tube (positioned vertically) via a plastic funnel. During the first trial of the experiment, the fish was allowed to swim freely until it left the long tube (positioned horizontally) into the outside tank filled with water (water height = 5 cm). This allowed the fish to acclimate to the new surroundings and minimize erratic swimming behavior caused by the narrow pathway. Starting from the second trial onward, each fish was given a maximum of 5 min to escape the tube. If the fish failed to do so within this time frame, it was compelled out of the tube by flushing it with water. Subsequently, the fish is excluded from consecutive trials. In the results description, the number of fish used for analysis is specified as a fraction of the total number of fish used for each trial. The duration taken by the fish to swim from the starting point A to the exit point B in the horizontal long tube (as depicted in Fig. 1) was recorded for further analysis. Once the fish left the tube, it was allowed to swim in outer tank before it was carefully captured using a small flat skimmer net to avoid aversive stimulus induced by being netted. Once captured, the fish was reintroduced into the short arm of the tube for subsequent trials. That is, the test fish was not returned to its home tank, and there was no inter-trial interval between training trials. The movement of zebrafish was further analyzed by video tracking using EthoVision XT, version 17.
Statistical analysis
All results were expressed as mean ± SEM. Statistical analysis was performed using GraphPad Prism software (version 10.0.0 for Windows, GraphPad Software Inc., USA). The effect size estimations were calculated using web application (https://www.estimationstats.com/#/). 24 The estimates were done using 5,000 bootstraps, and the confidence interval was bias-corrected and accelerated. Group differences are normalized to Cohen’s d. Statistical significance is shown as follows: *p < 0.05, **p < 0.01, and ***p < 0.001.
Results
Overview of experimental setup and quantification of memory acquisition of wild-type zebrafish
To assess the learning and memory capabilities of adult zebrafish using this paradigm, we conducted an experiment with a single fish. The fish underwent 12 trials, navigating through the long arm of L-shaped transparent tube. A camera placed overhead recorded its movements, while we measured the time it took for the fish to swim from point A to point B (referred to as escape latency) (see Fig. 1A). During the first (habituation) trial, the fish was allowed to swim freely in the maze for as long as the fish took to exit the tube at point B. Starting from the second trial, the training trials, the trial length was limited to 5 minutes. During the habituation trial (shown as trial 1 in Fig. 1D), we observed that both fish spent a longer time within the tube. They exhibited a back-and-forth swimming pattern in the long horizontal section of the L-shaped maze. However, in the second trial, we noted a significant reduction in the time spent by the fish in the long arm of the tube. This decrease in escape latency remained relatively stable with some fluctuations throughout the subsequent trials (see Fig. 1B).
These initial observations suggested to us that within only a few trials, escape latency robustly decreased, that is, the tested zebrafish showed a plateau of improved performance indicating quickly developed memory of prior exposure to the maze. Next, to determine the number of training trials are needed to obtain significant memory performance improvement (stable or plateaued reduction of escape latency), we trained a larger sample size of individual fish over 10 trials (n = 17/20). For this, we slightly modified the apparatus: we reduced the inner diameter of the tube from 2.1 to 1.6 cm. This adjustment was made to decrease inside volume of the tube as to minimize escape latency as the fish would be navigating through the tube rather than remaining inside. We assumed this would further increase the aversive nature of the apparatus, that is, it would provide a stronger motivation (unconditioned stimulus) for the fish to learn to escape from the tube. Similarly, to the procedure described above, an individual fish was introduced to the short arm of the tube at a time, and escape latency was measured. We found fish to spend increased amount of time in the tube in the first trial, but this time was significantly reduced in the second trial [one-way analysis of variance (ANOVA), Dunnett’s p = 0.0153] and with some fluctuation also in subsequent trials (Fig. 1C). Furthermore, the analysis indicated a significant reduction of escape latency reaching a steady low level from the first to the fifth trial (one-way ANOVA, Dunnett’s p = 0.0153) that did not show appreciable further change after the fifth trial. Thus, we decided that five trials would be sufficient to train an individual fish using this apparatus and procedure.
Comparison of wild-type and autism-model zebrafish escape latency before training
Subsequently, we decided to use our newly developed memory test paradigm and examine a zebrafish neurological disease model, the dyrk1aa deficient zebrafish, with it. This zebrafish was generated in our previous studies to model human DYRK1A deficiency, which showed social impairments related to autism. 22 Here, as before, each zebrafish was introduced to the short arm of the tube, and escape latency was quantified also as described above. Escape latency of dyrk1aa KO (n = 10/12) and wild-type fish (n = 18/20) was compared (Fig. 2A). We found KO fish to take significantly more time to escape the tube compared with wild-type fish (the unpaired Cohen’s d between WT and KO is 1.21, t = 3.073, df = 26, p = 0.0025). Furthermore, movement of KO fish indicated elevated activity in the novel environment of the tube compared with wild-type zebrafish (Fig. 2B, supplementary videos S1).
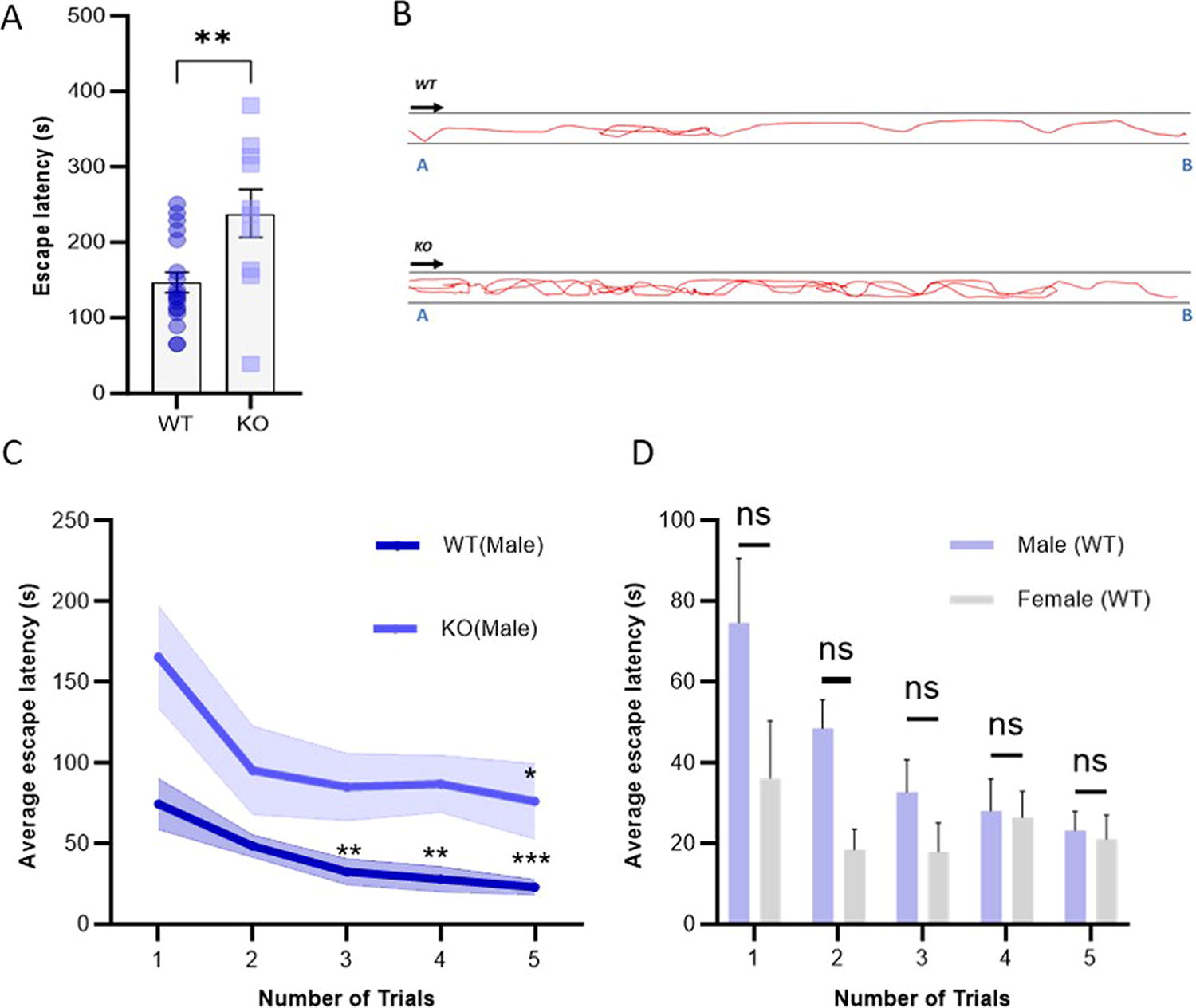
Comparison of wild-type and autism zebrafish escape time after training
To improve the escape latency, we decided to design the apparatus by reducing the length of horizontally positioned long tube. Here, the tube length was reduced to 50 cm (1.6 cm inner diameter). Learning from the above results, we subsequently trained individual wild-type zebrafish (n = 24/28) over five trials to investigate escape latencies in the shorter (50 cm long) and narrower (1.6 cm inner diameter) L-shaped tube. Compared with the first trial, the time had reduced significantly in subsequent trials (**p < 0.01, ***p < 0.001). Next, we exploited this modified apparatus to quantify escape latency exhibited by the autism model zebrafish (n = 10/10). As with wild-type zebrafish, we trained individual zebrafish over five trials. This showed a reduction of time over the number of trials indicating that the autism model zebrafish could also be trained using this method. However, the analysis indicated that only the fifth (one-way ANOVA, Dunnett’s p = 0.0478) trial was significantly reduced compared with the first trial (Fig. 2C). Furthermore, wild-type female zebrafish (n = 10/10) did not show significant reduction of escape time compared with the first trial while indicating overall reduced escape time (Fig. 2D).
Comparison of short-term memory in wild-type and autism zebrafish
To examine the ability of this apparatus to measure acquisition and short-term retention of memory in zebrafish, we decided to train both of dyrk1aa KO and wild-type zebrafish over five trials and perform a sixth trial after a 1-hour inter-trial interval between the fifth and the sixth trial. For this, each fish was housed in separate containers after their fifth trial. As described in the previous section, escape latency for individual fish was recorded using the overhead camera. Figure 3A shows that wild-type fish (n = 24/28) exhibited significantly reduced escape latency in the sixth trial compared with the first (unpaired Cohen’s d = 0.597, one-tailed unpaired t-test, t = 2.069, df = 46, p = 0.0221). However, the KO zebrafish (n = 10/10) showed no significant change in escape latency between the sixth trial (after the 1-hour inter-trial interval) and the first trial (the unpaired Cohen’s d = 0.0877, one-tailed unpaired t-test, t = 0.1960, df = 18, p = 0.4234).

Evaluation of long-term memory of wild-type zebrafish in tube assay
As described in the previous section, autism model zebrafish were not able to maintain memory after a one-hour long intertrial interval, but wild-type zebrafish showed excellent memory retention, that is, exhibited significantly reduced escape latency compared with their performance at the first trial, even after this 1-hour interval. Thus, next we examined whether wild-type zebrafish could remember even longer. For this, we trained individual fish (n = 14/15) over three trials during 3 consecutive days and measured the escape latency 4 days after the last training trial, that is, on the 7th day (Fig. 3B). Each fish was housed in separate containers during the study. The results showed a significant reduction in escape latency on the 7th day compared with the 1st day (paired Cohen’s d = −0.494, one-tailed paired t-test, t = 1.796, df = 13, p = 0.0479).
Effects of transparent versus different color painted tube walls
In a previous study, we found that zebrafish larva showed preference for certain colors. 25 We decided to investigate if coloring the walls of the tube affects habituation and learning performance of adult experimental zebrafish in the L-shaped apparatus. We covered two-thirds of the bottom side of the tube with a particular color to ensure that fish were able to see the colored background (as opposed to making the tube completely dark. We also employed the transparent tube as a control task and tested both dyrk1aa KO (n = 10/12) and wild-type (n = 30/32) adult zebrafish under these conditions. We found no significant difference in escape latency between the wild-type and dyrk1aa KO zebrafish in the colored tubes (in both the blue and yellow tube) (Fig. 3C). But in the transparent (control) tube, as before, we did find dyrk1aa KO zebrafish to exhibit significantly worse (higher) escape latency (unpaired Cohen’s d = 1.07, one-tailed unpaired t-test, t = 2.852, df = 32, p = 0.0038). Interestingly, we also found that performance of both genotypes of zebrafish was significantly worse in the blue tube compared the yellow: Two-way ANOVA, color effect [F(2, 108) = 5.229, p = 0.0068], genotype [F(1, 108) = 2.568, p = 0.1120], and interaction of color and genotype [F(2, 108) = 2.132, p = 0.1235].
Discussion
In this article, we present the development of a simple active avoidance/escape task designed to quantify learning and memory in zebrafish. We emphasize that the apparatus, the L-shaped tube, is simple and cheap to make, and we show that zebrafish trained and tested in this apparatus are able to acquire memory within five trials. We also show that our procedure using this apparatus is capable of detecting significant impairment in a mutant zebrafish, the dyrk1aa KO fish, compared with control wild-type zebrafish. Thus, we argue that our newly developed apparatus and procedure offer a simple and fast method with which one can assess learning and memory and its modification in adult zebrafish.
In recent years, zebrafish have been widely used to model a variety of CNS disorders, and several of these studies employed quantification of behavior to assess functional changes in the brain. 1 Many of these studies used learning paradigms, as numerous brain disorders are associated with memory impairment and/or attention deficits. 2 In the overwhelming majority of such learning studies, zebrafish were tested individually, that is, one at a time. However, the zebrafish is a highly social species that exhibits shoaling, strong preference to stay in proximity to one another.2,6 Thus, social isolation in these learning tasks created an aversive condition. Given that most learning tasks employed reward as the US (most often food reward, but see Al-Imari et al. 26 and Karnik et al. 27 ), the anxiety-fear-inducing effect of social isolation interfered with the rewarding effect of the employed US in zebrafish. One way to circumvent the confounding effect of fear anxiety was to employ long and repeated habituation trials.14,28 Habituation stems from both nonassociative and associative processes. More precisely, short-term habituation results from a nonassociative process, whereas long-term habituation is a consequence of an associative process. In addition, certain conditions can lead to competition between these distinct processes. 29 Traditionally in zebrafish experiments, maze habituation takes several days, but our new method allows fish to swim freely in the long tube during the first trial without time restrictions. We believe that this step allows to reduce the time used for habituation in the most maze protocols.
However, although not clearly reported, nor properly explored in the literature, such habituation trials could achieve the opposite: if the fish are not handled gently, they can lead to sensitization, that is, instead of reduced fear/anxiety, they could enhance fear/anxiety. As handling is a delicate matter in zebrafish studies, and depending on which fish were handled more or less gently, some may habituate while others may sensitize to the apparatus and the procedure, which thus leads to enhanced experimental error variation. Thus, larger number of required habituation trials does not only require more time, but may also reduce statistical power to find treatment effects significant. This may be one of the reasons that most prior learning tasks required a large number of training trials subsequent to the habituation procedure. During these sessions, the animal is introduced to the maze and encounters visual cues, aversive stimuli, or rewards. The repeated exposures to these stimuli enable the animal to learn and establish associations between the visual cues, acting as CS, and an US that elicits innate preference or avoidance responses. 13 However, this associability of a stimulus can be altered. 30 We emphasize that our simple apparatus and procedure required no such extensive habituation process, and the experimental zebrafish showed robust and lasting memory performance within as few as five training trials. We emphasize that our simple apparatus and procedure required no such extensive habituation process, and the experimental zebrafish showed robust and lasting memory performance within as few as five trials. In contrast to a number of prior methods that use a variety of maze designs, for example, T, Y, and + shaped mazes, our method employs a transparent acrylic tube that represents a restrictive, physically restraining environment. Based upon field observations as well as laboratory studies, 31 we now know that zebrafish prefer open environments and find physically restrictive places (covered or small tanks in the lab or caves or crevices in nature) aversive. Our maze design uses this ecologically/ethologically relevant feature of the species, as our study shows that the small diameter tube represents an aversive environment from which zebrafish are highly motivated to escape. In the pilot study, we noticed that during the initial trial, zebrafish exhibited a back-and-forth movement in the tube. This behavior may be influenced by the unfamiliar environment, prompting escape-like responses from the fish. The speed with which the fish go through the L-shaped tube and exit the longer arm is easy to quantify (escape latency) using a simple stopwatch, and thus not even complicated video-tracking systems may be required for the quantification of behavioral response. The escape latency has been significantly reduced over the trials indicating that the fish learn in this tube, that is, their prior experience leads to improved performance, the definition of memory and this memory acquisition occurs fast. What exactly the fish actually learn is unknown at this point, but it may include that turning in the maze is impossible or difficult, that there is an exit point, and perhaps even the specific layout of the maze, for example, that there is a short vertical and a longer horizontal part. Irrespective of what the fish actually learn, however, this simple apparatus and fast procedure we demonstrate is able to elicit significant acquisition of memory, and it is also capable of detecting a genetic manipulation–induced impairment of memory performance. Furthermore, this apparatus uses both short-term and long-term memory tasks. Short-term memory is characterized by experience-dependent changes in behavior lasting only a few minutes, 32 and this could be varied in zebrafish.10,33 Studies exploring the mechanisms of short-term memory in mammals have revealed distinctions from those involved in medium- or long-term memory, 34 and it is feasible to distinguish between short-term memory and long-term memory, each having unique features. 35 Thus, it can be used to assess both short-term as well as long-term memory in adult zebrafish. Given the simplicity of the design of the apparatus and of the procedure, and given the easy method of quantification of the behavioral response in this task, we also argue that the task will be useful for high-throughput screening. For example, if one equips the maze with two photocell detectors, one at point A (the start of the horizontal part of the tube) and another at point B (the exit point, the end of the horizontal part of the tube), one can simply and cheaply quantify escape latency. Multiple such tubes may be run in parallel, and thus the only factor limiting the throughput of the test is how fast the experimenter is able to “load” the fish into these tubes.
Although the concept of the maze worked in practice the first time we tried it, we attempted to optimize, that is, fine-tune some parameters of the apparatus and the procedure. For example, we first used a 70 cm long tube whose diameter was larger, that is, 2.1 cm inner diameter. This apparatus turned out to be suboptimal. Next, we reduced the inner diameter of the tube tunnel from 2.1 to 1.6 cm in order to minimize the ability of the fish to turn in the tube. The narrower tube led to shorter escape latencies, that is, faster runs, without the reduction of the maze’s ability to quantify learning and memory or to detect difference between our mutant and wild-type zebrafish. Next, we also reduced the length of the maze from 70 to 50 cm without jeopardizing effectiveness of the apparatus. Subsequently, we examined how many training trials we must conduct to find reliable memory performance and found that as few as five are sufficient. Admittedly, these efforts of ours to optimize the physical parameters of the maze and the procedure of the task resulted in only preliminary findings, as we did not systematically and parametrically alter the above parameters. Nevertheless, they have demonstrated that the conceptually and physically simple maze design and fast procedure is effective and efficient.
The last question we consider is the interpretation of the difference we observed in memory performance between the dyrk1aa KO and the wild-type control zebrafish as quantified by our newly developed task. The human DYRK1A gene encodes a kinase and maps to the obligate down region of chromosome 21. 36 Mutations of this gene have been shown to lead to microcephaly and intellectual disability and have also been implicated in autism. 37 Overexpression of this gene in rodents has been shown to recapitulate some of the symptoms of Down’s syndrome and severe learning and memory defects.22,38 However, KO mouse studies aimed at modeling the role of this gene in autism have been hampered by lethality found in homozygous null mutants.39,40 Given the tetraploid nature of the zebrafish genome, the KO approach is expected not to lead to this problem, and indeed a zebrafish KO model, the dyrk1aa KO zebrafish, was more successful. 21 In this model, the zebrafish were found to exhibit impaired social behavior, mimicking a major autism spectrum disorder phenotype.
Learning and memory or other behavioral functions have not been explored in the dyrk1aa KO fish until this current study. Here, we demonstrate significant memory performance deficit in the dyrk1aa KO zebrafish compared with wild-type control zebrafish, detected the first time with our newly developed learning and memory paradigm. Performance in a learning and memory task may depend upon four fundamentally distinct factors: one, learning and/or memory, two, motor function, three, perception, and four, motivation. Learning and memory itself may be further dissected into distinct components, including attention, acquisition of memory, consolidation of memory, retention of memory, and recall. Which of these factors may be affected by the null mutation in the dyrk1aa KO zebrafish, we cannot tell at this point. However, some speculative arguments may be made based upon the acquisition curves and memory performance results shown in Figure 2A. First, the performance of the dyrk1aa KO zebrafish is impaired from the first trial onward. As we do not expect the fish to be familiar with the maze, that is, to have memory of the maze at this stage of the training, a deficit at the first trial suggests that the mutant fish suffer from a performance deficit unrelated to memory, that is, may have impaired motor function, perception or motivation. It is also possible that attention is impaired in these mutant fish, as attentional problems analogous to attention-deficit hyperactivity disorders could manifest as impaired performance even at the first trial. At this point, which of these performance features may be impaired in the dyrk1aa KO fish is unknown.
A prior study 21 and our own personal observations suggest that trivial motor performance deficit is unlikely, as the fish were found to swim normally and appeared healthy. However, in novel tank assay, reduced anxiety previously reported for the dyrk1aa KO zebrafish could have contributed to the observed learning and memory performance deficit, as less anxious fish may be less motivated to escape from the aversive L-shaped maze of the current study. Reduced anxiety, however, is unlikely to explain our results suggesting that both short-term and long-term memory were impaired in the dyrk1aa KO. We speculate that these mutation effects may be unrelated to the above discussed performance factors. For example, perusal of Figure 2C shows that dyrk1aa KO zebrafish were able to improve their performance as the training progressed, that is, showed significantly reduced escape latencies compared with the escape latency at the first trial even though the improvement did not reach the level of performance of the wild type control fish. Importantly, these wild-type fish maintained their improved performance after a 1-hour inter-trial interval, whereas the dyrk1aa KO zebrafish worsened back to a level that was statistically indistinguishable from their performance at the very first trial. This memory performance deficit of the dyrk1aa KO fish is unlikely to be due to motor function, perception, or motivation (including reduced anxiety) and is more likely to be due to impaired consolidation, retention, and/or recall of short-term memory. In this study, we did not observe sex-based differences in memory and learning in zebrafish as described in previous studies utilizing mazes.15,41 In addition, we did not test dyrk1aa mutant female zebrafish using this apparatus.
Conclusion
In this study, we describe the design of a new apparatus, an L-shaped tube, and provide a novel procedure that allowed us to train zebrafish and test their memory performance. We show that in this active avoidance/escape paradigm, zebrafish are able to acquire significant memory within five trials and without extensive prior habituation procedures and that this memory is maintained for at least 4 days after the last trial. Furthermore, we also show that the paradigm is able to detect a mutation induced difference between wild-type and a KO zebrafish line, the dyrk1aa KO zebrafish. The apparatus is easy and cheap to make, the paradigm is simple to run, and the behavioral measure of memory performance is easy to quantify. Thus, we argue that this new paradigm, unlike most other learning and memory tasks developed for the zebrafish in the past, will be useful for high-throughput screening of both mutations and compounds (small molecules) for learning and memory altering properties. We also acknowledge that, as with other learning and memory tasks, our new paradigm also cannot give a final answer as to what aspects of learning and memory performance may be altered by the experimental manipulation employed. We emphasize that once a positive hit (a significant mutation effect, as in our case, or a significant drug effect) has been identified using the paradigm, subsequent follow-up studies using a battery of behavioral tests may be required to dissociate the effects of factors known to potentially influence learning and memory performance in experimental animals. Nevertheless, we believe that our newly developed simple yet efficient paradigm will be used as an excellent screening tool, and as such it will contribute to modeling of, and the identification of the mechanisms underlying, neurological, neuropsychiatric, and neurodevelopmental disorders of the human brain.
Footnotes
Acknowledgment
This research was supported by Chungnam National University.
Authors’ Contributions
Design concept: D.W.D., T.Y.K., B.N.H., J.S.L., T.H.K.; investigation: D.W.D., T.Y.K.; writing—original draft: D.W.D., R.G., C.K.; writing—Review and Editing: D.W.D., R.G., C.K.
Disclosure Statement
The authors declare no conflict of interest.
Funding Information
No funding was received for this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.