
Abstract
The transgenic (TG) zebrafish allows researchers to bio-image specific biological phenomena in cells and tissues in vivo. We established TG lines to monitor changes in the ovaries of live fish. The original TG line with ovarian fluorescence was occasionally established. Although the cDNA integrated into the line was constructed for the expression of enhanced green fluorescent protein (EGFP) driven by the medaka β-actin promoter, the expression of EGFP is restricted to the oocytes and gills in adult fish. Furthermore, we found that germinal vesicles (GVs) in oocytes of the established line can be observed by relatively strong fluorescence around the GV. In this study, we tried to capture the dynamic processes of germinal vesicle breakdown (GVBD) during meiotic cell division using the GV fluorescent oocytes. As a result, GV migration and GVBD could be monitored in real time. We also succeeded in observing actin filaments involved in the migration of GV to the animal pole. This strain can be used for education in the process of oocyte meiotic cell division.
Introduction
Meiotic cell division of oocytes (oocyte maturation) in fish is triggered by luteinizing hormone (LH) secreted from the pituitary gland, which acts on follicle cells to induce the production of maturation-inducing hormone (MIH), 17,20β-DHP, in follicle cells (Fig. 1). In zebrafish, LH secretion occurs at midnight before spawning. 1 It is predicted that the induced production of 17,20β-DHP will be completed around 2–3AM under our laboratory conditions, while the zebrafish are sleeping (in our lab we turn off the lights at 11PM and turn on the lights at 9AM). Then, 17,20β-DHP activates ovulation-inducing pathway in follicle cells by binding to nuclear progesterone receptor (Pgr) through autocrine or paracrine signaling. Fish follicle cell layer consists of an outer theca cell layer and an inner granulosa cell layer, where theca cells proceed to synthesize 17β-hydroxyprogesterone, the precursor of 17,20β-DHP, and granulosa cells catalyze the conversion of 17β-hydroxyprogesterone transferred from theca cells to 17,20β-DHP to complete the production of MIH, 17,20β-DHP. 2

Induction pathway of oocyte maturation and ovulation in zebrafish. When LH secreted from the pituitary acts on follicle cells, it induces the production of MIH, 17,20β-DHP, in follicle cells. Then, 17,20β-DHP activates the ovulation-inducing pathway in follicle cells by binding to the nuclear Pgr. Secreted 17,20β-DHP also act on mPR on the cell membrane of oocytes to induce oocyte maturation. The ovulation induction pathway in follicle cells is called as the genomic pathway because Pgr bound to MIH acts as a transcription factor and activate the transcription. Contrary, oocyte maturation-inducing pathway in oocyte called as nongenomic pathway because that is activated without transcription of genes by mPR mediated intracellular signal transduction pathway. LH, luteinizing hormone; MIH, maturation-inducing hormone; mPR, membrane progesterone receptor; Pgr, progesterone receptor.
We hypothesize that the same thing happened in zebrafish. Secreted 17,20β-DHP from granulosa cells acts on membrane progesterone receptors (mPRs) on the cell membrane of oocytes to induce oocyte maturation.3–5 Oocyte maturation is completed by 5AM on the morning of the spawning day. Then ovulation of oocytes is completed by 8AM. After light at 9AM, fish wake up and start sexual behavior for mating if females have ovulated eggs in their abdomen. After oocyte maturation, the name oocyte changes to egg. Mating continues for 1–2 h (9–11AM), so fertilization of eggs occurs during this period. 1
Ovulation-inducing pathway in follicle cells called as genomic pathway because that is activated by transcription of genes by MIH-bound Pgr acting as transcription factor. 6 In contrast, oocyte maturation-inducing pathway in oocyte called as nongenomic pathway because that is activated without transcription of genes by mPR mediated intracellular signal transduction pathway. 7 The nucleus of the oocyte germinal vesicle (GV) is located midway between the animal pole and the center of the oocyte. Upon stimulation with 17,20β-DHP, GVs begin to move toward the animal pole (Fig. 2). After reaching at animal pole, GV will disappear by nuclear breakdown (germinal vesicle breakdown, [GVBD]) during first meiotic cell division. Oocyte continues meiotic cell division until metaphase of second meiotic cell division and then division will be stopped. During these steps, the oocyte becomes transparent as shown in the photo on the bottom right (Fig. 2).

GV migration during oocyte maturation. The GV is located at the center of the oocyte. After stimulation by MIH, GV starts moving to the animal pole. After lysis at the animal pole, GV will disappear by nuclear breakdown GVBD onset of first meiotic cell division. Oocyte continues meiotic cell division until metaphase of second meiotic cell division and then division will be stopped. During these steps, the oocyte becomes transparent as shown in the photo on the bottom right. GV, germinal vesicle; GVBD, germinal vesicles breakdown.
The transgenic (TG) technique is well established in zebrafish. 8 For example, a TG with germ-specific expression of green fluorescent protein (GFP) is a useful tool to visualize germ cells produced by fusing the GFP gene to the regulatory regions of the vasa gene. 9
Hsiao et al. established a TG zebrafish line with a fluorescent ovary. 10 The cDNA is integrated into this strain to express enhanced green fluorescent protein (EGFP) driven by the medaka β-actin promoter; however, EGFP expression is restricted to oocytes and gills in adult fish. The nuclei (GVs) of oocytes in the TG line are fluorescent because EGFP is loosely bound to the nucleus. 11
The bodies of wild zebrafish are naturally opaque and their ovaries are not externally visible. This makes noninvasive studies of the ovaries difficult, even in the TG line. Therefore, we tried to produce transparent zebrafish with fluorescent ovaries by crossing the wild-type TG fish and transparent strain, roy.
We succeeded in establishing a TG line with transparent bodies and ovarian fluorescence (named β-roy). 12 Using the β-roy fish, we successfully monitored the changes in the ovaries during a sex reversal experiment. 13 In this study, we used oocytes of the strain to monitor the migration of GVs during meiotic cell division. As expected, we were able to observe GV migration in real time. We believe that the strain can be an instructive source for meiotic cell division and the dynamics of movement of materials in cells by microfilaments.
Materials and Methods
The TG zebrafish line
The TG line, β-roy, was established by normal mating of the original TG line TG (β-actin: EGFP): the cDNA for the expression of EGFP driven by the medaka β-actin promoter 10 was integrated into this line with roy−/−. The strain was named β-roy, TG (β-actin: EGFP);roy (transparent zebrafish) and maintained as a closed colony for research. 14 The strain has been deposited in the National BioResource Project under the identifier TG (β-actin: EGFP);mpv17+/a9 (the gene responsible for the roy strain has been identified as mpv, mitochondrial inner membrane protein MPV17, and given a unified name for several different strains including roy mutations as a9). Zebrafish were maintained in an aquarium at 28.5°C with a 14-h light/10-h dark cycle. 15 All zebrafish experiments were conducted with the approval of the Institutional Ethics Committee of Shizuoka University, Japan (approval No. 2022F-3 and 2023F-9); the guidelines for animal use established by this committee were strictly followed.
Microscopic observation of GV migration
Zebrafish ovaries were isolated from sacrificed females and placed in fresh zebrafish Ringer's solution (116 mM NaCl, 2.9 mM KCl, 1.8 mM CaCl2, and 5 mM HEPES, pH 7.2) and washed with the same solution. Oocytes were manually isolated from the ovaries using fine forceps. Fully grown immature oocytes were exposed in vitro by incubating the oocytes in zebrafish Ringer's solution containing 17,20β-DHP (diluted from a 1000-fold stock in ethanol) at 25.0°C or room temperature. A binocular microscope with a GFP filter (SZX12, Olympus, Japan) was used for fluorescence imaging. Images were captured using a digital camera (DP-70) and stored using software (DP controller).
Observation of actin filaments during GV migration
Acti-stain™ 488 fluorescent phalloidin (Amanita phalloides) was purchased from Cytoskeleton, Inc., (Denver, CO). The fluorescent agent was microinjected into the oocyte cytoplasm after stimulation of oocyte maturation by 17,20β-DHP or Org OD-02. 16 After injection, the microfilaments in the oocyte were observed using a binocular microscope with a GFP filter (SZX12, Olympus, Japan).
Results and Discussion
We tried real-time observation of GV migration using oocytes of ovarian fluorescent TG zebrafish, β-roy (Fig. 3). GVs in oocytes of β-roy grow more strongly that cytoplasm by GFP fluorescent. After stimulation by 17,20β-DHP, the migration of GV could be monitored under the microscope until the completion of GVBD. Fluorescence images were taken at 5-min intervals (Fig. 4). At high magnification, micropyle could be observed on the animal pole (Fig. 5). The micropyle is a tubular structure formed during oogenesis at the animal pole of fish oocytes as an entry point for spermatozoa. The micropyle is closed when a single sperm enters the oocyte and is thought to play a role in polyspermy rejection.
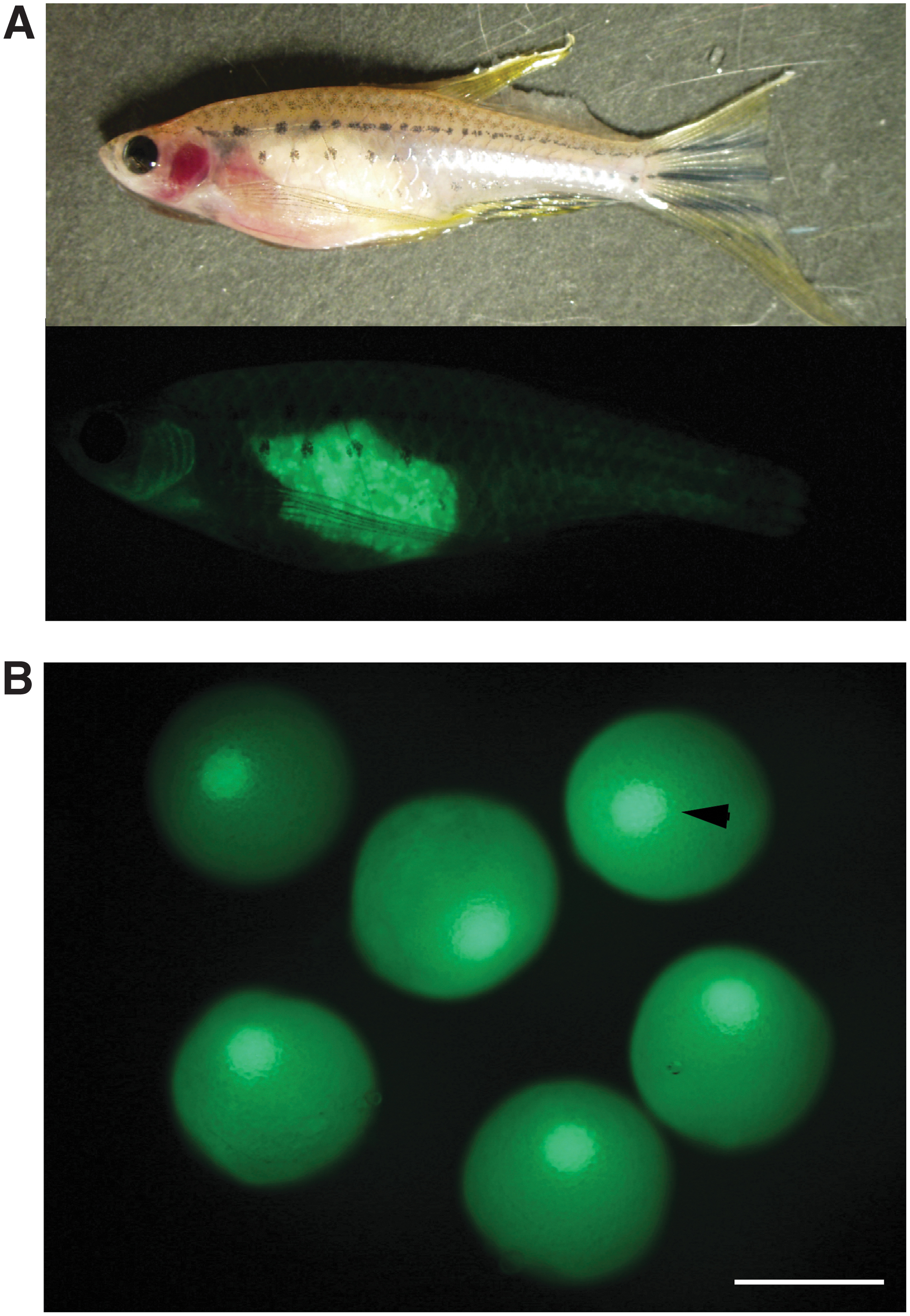
GV of β-roy can be observed by fluorescence of EGFP. Ovaries of β-roy female grow by EGFP fluorescence in live zebrafish as shown in panel
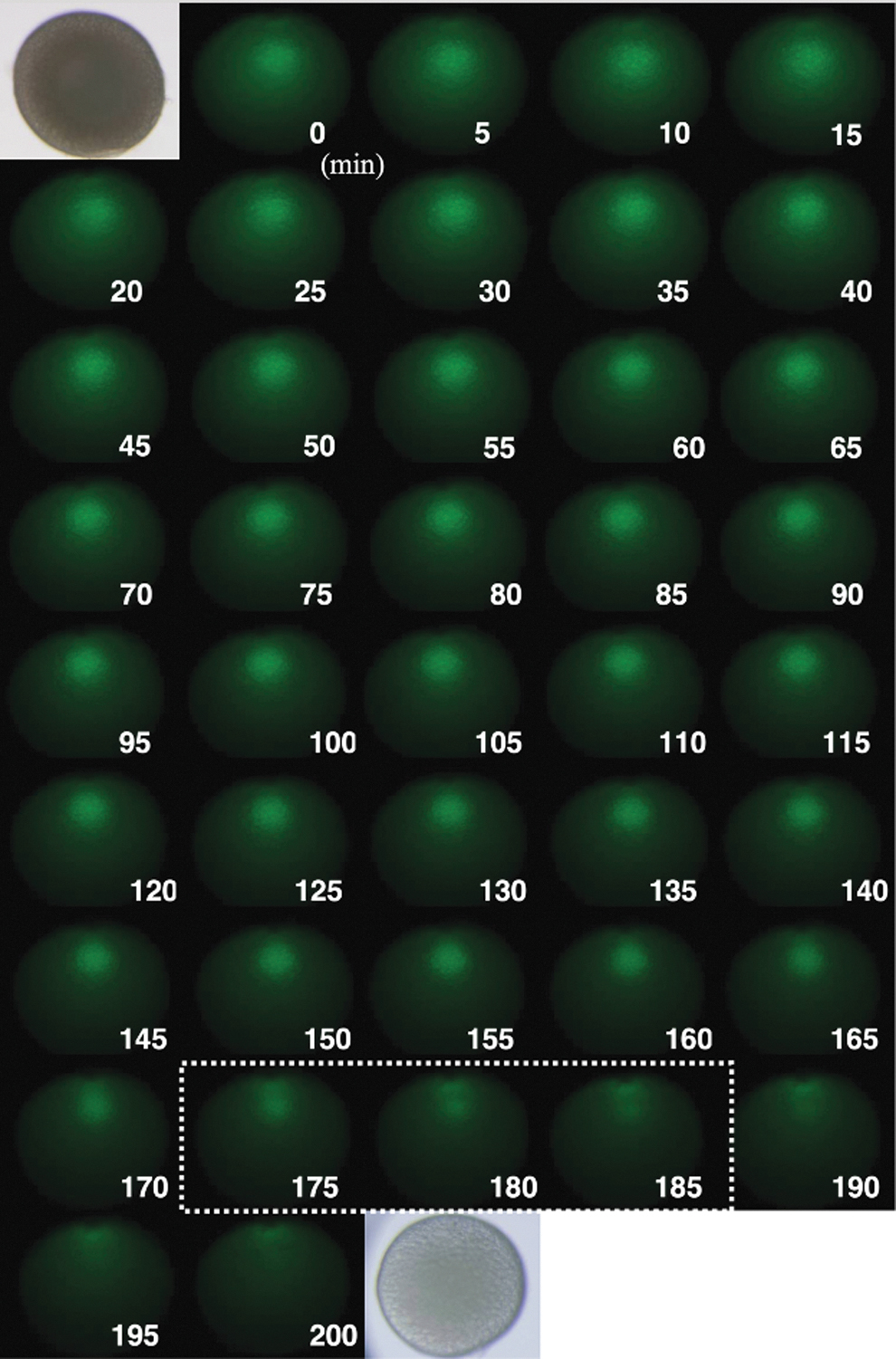
Real-time observation of GV migration during oocyte maturation. After stimulation with MIH, GV in the oocyte was photographed under GFP filter views at 5-min intervals. In this oocyte, GVBD occurred during 175 to 185 min after the addition of MIH. Photographs of the oocyte under bright field before and after incubation are also shown in the upper left and lower right. GFP, green fluorescent protein

Enlarged image of EGFP-expressing oocyte of β-roy. GFP filter views of β-roy oocyte during GV migration were photographed. The micropyle can be seen at the center of the animal pole. GV are migrating toward micropyle. There is a gradient of contents inside the oocyte from vegetal pole to animal pole and vice versa as indicated by an arrow. A movie prepared by stacking photographs during GV migration is available as Supplemental Movie.
We prepared a short time movie by stacking the photos before and after GVBD (Supplementary Movie S1). In the movie, migration of GV toward the micropyle and GVBD after reaching the animal pole can be observed. We also tried to observe actin filaments, which are shown to make the force of GV movement in goldfish. 17 By injecting fluorescence-labeled phalloidin into oocytes during oocyte maturation, we were able to stain the actin filaments formed during GV migration. Actin fibers were formed as they surrounded the GV during migration (Fig. 6A). Also, spreading actin filaments surrounding GVs were observed immediately after GVBD (Fig. 6B). This localization of actin fibers suggests that bundles of actin fibers move the GV to the animal pole in zebrafish.
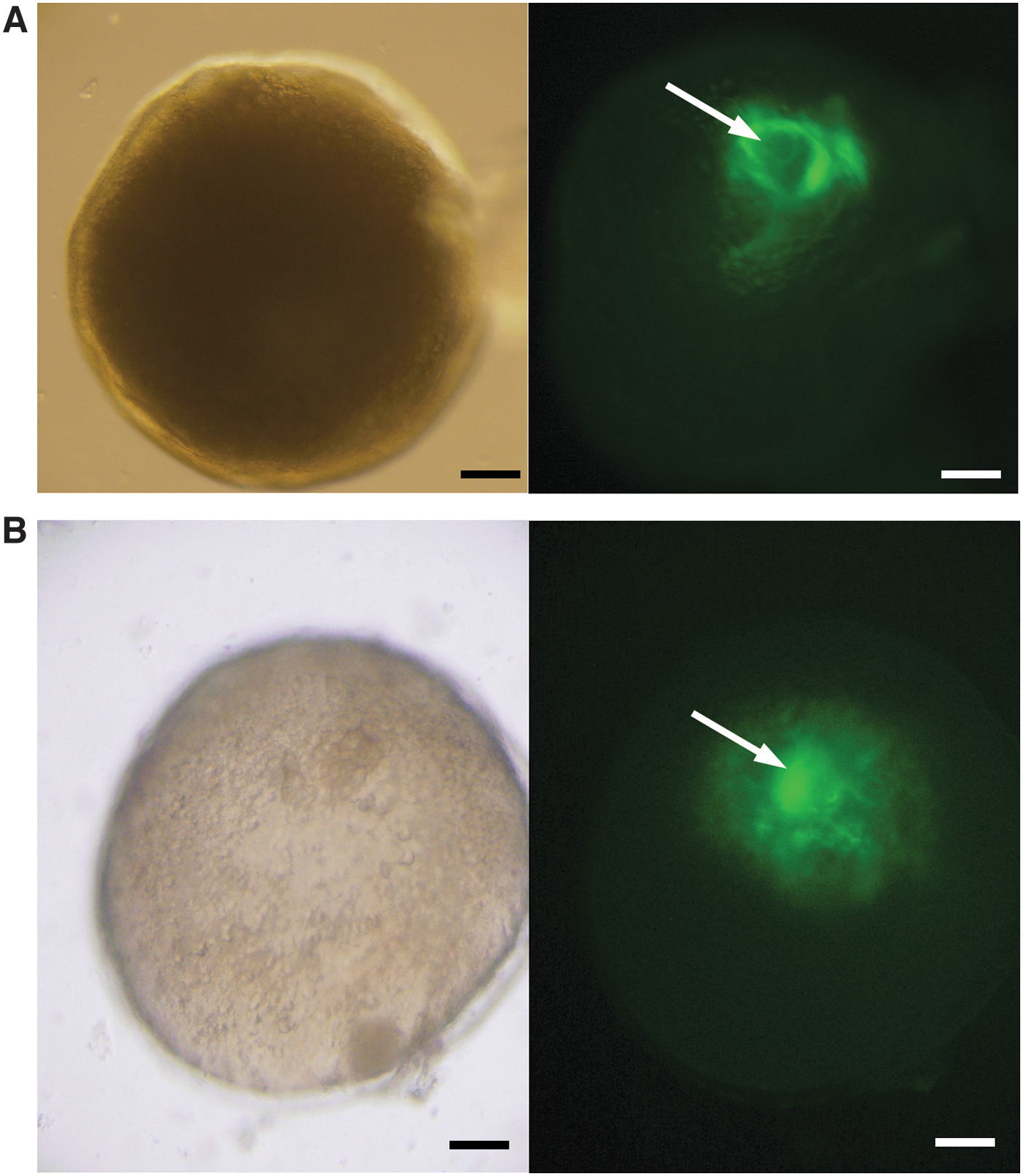
Actin filaments surrounding GV during migration. Actin filaments in oocytes were visualized with Acti-stain™ 488 fluorescent phalloidin. GFP filter views of Roy's oocyte during GV migration were photographed. GV is surrounded by actin filaments during migration
In this article, we presented the use of our TG strain to visualize the movement of the nucleus of zebrafish oocytes. The strain is fertile and capable of maintaining a population. In the β-roy fish, the ovaries in living fish are visible from the outside of the body with the naked eye or with a fluorescence microscope. Although not shown in this study, this fish model has useful research applications because it allows continuous observation of the internal organs of the same individual throughout its life, as demonstrated by the observation of EGFP expression during development. Thus, this strain provides a useful alternative model for noninvasive studies of normal and pathological changes in gonadal structures at later stages of life and the genesis of other diseases and changes caused by exposure to chemical substances. 13
Finally, the following tables show how to observe this GV migration in a teaching laboratory (Table 1) and tips for using the laboratory course in biology education (Table 2).
Method of Observing Migration of Germinal Vesicle
GV, germinal vesicle; GVBD, germinal vesicles breakdown; MIH, maturation-inducing hormone.
Tips for Using the Experimental Course in Biology Education
LH, luteinizing hormone.
Footnotes
Acknowledgments
We are grateful to Dr. N. Sakai at National Institute for Genetics for valuable advice for zebrafish maintenance.
Authors' Contributions
T.T. has consistently studied the molecular mechanisms of oocyte maturation and ovulation in frog and fish since his undergraduate and graduate studies. Recently, he has been focusing on the structure and function of steroid membrane receptors, which are receptors for oocyte MIHs. In addition to teaching courses in the areas of basic biology and developmental biology, he has been teaching experiments on meiosis induction and degradation of yolk proteins during early development in zebrafish, as shown in this study. Based on this educational experience, he planned this study. The slides and movies shown in this study are those used in the lectures.
E.T. has been conducting functional analysis of ovulation induction-related genes in zebrafish in the Tokumoto Laboratory as a graduate student by establishing gene knockout lines through genome editing. Currently, he is continuing his Ph.D. research on the mechanism of egg membrane formation after fertilization. In this study, he was mainly responsible for observing actin fibers that bind to GVs using microinjection techniques.
Disclosure Statement
No competing financial interest exists.
Funding Information
This work was supported by Grants-in-Aid for Scientific Research on Priority Areas from the Ministry of Education, Culture, Sports, Science and Technology of Japan; JSPS KAKENHI Grant Number 23K05830 (to T.T.).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.