
Abstract
Fipronil is a broad-spectrum insecticide that has off-target effects in developing vertebrate embryos. In this study, we investigate treatment of zebrafish embryos with fipronil over the course of 5 days and examine the effects on body length, the cardiovascular system, and craniofacial morphology. We found the insecticide caused shorter body length and a decrease in eye size. By examining specific heart chamber morphology, as well as jaw angle and length, we quantified defects including enlargement of the heart and increases in jaw length and width. Further studies are needed to assess the mechanisms of fipronil's effect on vertebrate development for both environmental and human health concerns.
Introduction
Fipronil [5-amino-3-cyano-1-(2, 6-dichloro 4 trifluoromethylphenyl)-4-trifluoromethyl sulfinyl pyrazole] is a broad-spectrum insecticide of the phenylpyrazole class. 1 Fipronil was developed in the early 1990s to protect against termites, cockroaches, fleas, and ants, among other pests.2,3 It is currently one of the most commonly used broad-spectrum insecticides, approved for use in more than 120 countries around the world. 4 Through skin absorption or ingestion, fipronil antagonistically binds with the gamma-aminobutyric acid (GABA)-regulated chloride channel receptor, thus inhibiting chloride ion movement and causing continuous central nervous system activity, leading to hyperexcitation, convulsions, paralysis, and death.1,3,5 Fipronil is a popular insecticide due to its long persistence after application and its high-binding affinity to the GABA-receptor in target insect invertebrates, making it highly toxic to target pests.3,4
However, recent studies have begun to show the unintended consequences of fipronil on nontarget invertebrate and vertebrate organisms, along with their ecosystems. In the environment, fipronil can undergo many diverse chemical reactions producing numerous metabolites. 3 Fipronil has a half-life of over 1000 days in the soil, and some fipronil metabolites have an even longer half-life, resulting in their extended persistence in the environment.3,4
This prolonged environmental contamination has resulted in the bioaccumulation of fipronil in nontarget organisms. 6 In one case, fipronil was sprayed to control migratory locust pests; however, nonpest harvester termites (Hodotermitidae) were also killed, resulting in the indirect population reduction of two reptiles (Chalarodon madagascariensis and Mabuy elegans) whose diet heavily relied on the termites. 7 For blue land crabs (Cardisoma guanhumi), fipronil exposure led to decreased protein content and both hematological and oxidative damage of the crabs' oxy-hemocyanin. 8 Long-term, low-dose fipronil exposure can impact behavior in female mice, including hyperlocomotive behaviors, indicating the possible accumulative effect of fipronil in organisms. 9
Agricultural insecticides additionally have the potential to contaminate human food sources. One such large-scale egg contamination incident in 2017 resulted in fipronil entering the human food chain through fipronil-contaminated chicken eggs. 10 Particularly concerning is the fact that large rainfalls can flush fipronil from agricultural fields or residential areas, leading to contamination of waterways and bioaccumulation in aquatic organisms. 6 Environmentally relevant concentrations of fipronil (∼1–200 nM) can affect aquatic life. 11 Fipronil impacts the aquatic food chain as indicated by impaired medaka (Oryzias latipes) growth after fipronil treatment that reduced the density of invertebrate food available. 12
Zebrafish are an ideal model organism to investigate the effects of fipronil exposure to aquatic vertebrates during embryogenesis as they have fast generation times and thrive in captivity. They are prolific oviparous breeders producing eggs that are transparent during embryogenesis, making them ideal for observation of development. 13 Much is already understood about zebrafish biology and physiology, and since zebrafish share genes and cell signaling pathways with many vertebrates including humans, results gathered from studies utilizing zebrafish can often be translated to similar conditions or disorders in humans. 14
In previous studies, fipronil damaged lateral hair cells and brain tissue in zebrafish, reduced survival rate, impaired locomotive function, 15 and caused notochord degeneration, shortened body axes, muscle contraction malformations, and abnormal tail development at fipronil concentrations of 0.7 μM or greater. 2 Other studies have indicated cardiac malformations including an increased risk of heart edema, 16 increased heart rate, increased heart area, decreased cardiac function, disrupted blood vessel formation, 17 and increased intersegmental vessels. 18 Another study has reported a truncation of the lower jaw 16 ; however, the effect of fipronil on specific heart chambers and on specific jaw defects has not been quantified. Thus, we aim to investigate how dose–dependent fipronil exposure might affect the embryonic development of zebrafish (Danio rerio) in these areas. Fipronil concentrations in other studies range from nanomolar range 19 to over 30 μM 17 ; so we focused on concentrations near and above 0.7 μM where development neurotoxicity has been previously reported. 2
We hypothesized that increasing concentrations of fipronil would lead to increasingly severe developmental abnormalities, specifically abnormal eye size, body length, heart chamber morphology, and jaw morphology. Treatment of zebrafish embryos at concentrations of 0.7, 1.0, and 1.5 μM fipronil between fertilization and 5 days postfertilization (dpf) indicated dose-dependent changes in those developmental areas.
Materials and Methods
Fish husbandry and treatment
Adult zebrafish (AB strain) were kept in an enclosed room on a light/dark cycle of 14 h/10 h, respectively. Embryos were obtained by setting up groups of adult zebrafish overnight followed by embryo collection at 1–2 h postfertilization as zebrafish typically lay soon after room lighting turns on. The embryos were kept in an incubator at 28.5°C in E3B (5 mM sodium chloride, 0.17 mM potassium chloride, 0.33 mM calcium chloride, 0.33 mM magnesium sulfate, and 0.00025% methylene blue). Fipronil (Sigma 16785) was dissolved in methanol to create 1000 × stock solutions.
Control embryos were treated with 0.1% methanol dissolved in E3B. Embryos in fipronil treatment groups were treated with 0.7, 1.0, and 1.5 μM fipronil dissolved in methanol and diluted in E3B for a final methanol concentration of 0.1%. Treatment conditions never exceeded 0.1% methanol. Treatments began at 6 h postfertilization (hpf) and were refreshed daily until 5 days postfertilization (dpf) when final data were collected. Three biological replicates (using three clutches of embryos) were performed for each treatment concentration with total sample sizes between 30 and 75 as described below. Protocols were approved by the Roanoke College IACUC as study #14BIO76.
Length, eye, and jaw measurements
Length, eye, and jaw measurements were taken using an Olympus FSX-BSW. Embryo length, eye diameter, and eye area were measured by positioning the embryo laterally at 3, 4, and 5 dpf.
Initial studies for jaw differences were performed using Tg(sox10:gfp), which expresses GFP in cartilage. For more accurate measurement in fixed specimens that could be oriented more precisely, Alcian blue was used to stain zebrafish embryonic cartilage for jaw measurements (Fig. 1). Embryos at 5 dpf were fixed with 4% paraformaldehyde at 4°C overnight. Embryos were stained as previously described.20,21 In brief, embryos were washed in PBS before being stained overnight in 0.1% Alcian blue, washed in acidic ethanol 5 × for 1 h each time to de-stain and dehydrated using 100% ethanol. Once stained, embryos were photographed using an Olympus FSX-BSW. Length and angle measurements were computed using the Olympus FSX software and NIH ImageJ as previously described.22–24

Jaw measurements used to determine craniofacial abnormalities in zebrafish embryos.
Heart area and rate
Structural heart measurements at 5 dpf were taken using an Olympus FSX-BSW using transgenic fish that express green fluorescent protein in the developing heart, Tg(cmlc2:gCaMP) also known as Tg(myl7:GCaMP). 25 Area measurements of the atrium and ventricle were taken using Olympus FSX-BSW software.
Heart rate at 3, 4, and 5 dpf were collected using AB zebrafish. Heart rate was measured for a period of 15 s using a manual tally counter while observing embryos under a dissection microscope; the data were converted to beats per minute.
Statistical analysis
After confirming normality, most data sets were analyzed using ANOVA for p < 0.05, followed by Tukey's post hoc test for pairwise comparisons where p < 0.05; data for the prevalence of edema were analyzed using a z-score for p < 0.05. Samples consisted of three biological replicates (three clutches of embryos). Total sample size at each concentration was 75 for length, eye diameter, eye area, edema, and heart rate, 60 for heart area, and 30 for the jaw measurements.
Results
Fipronil results in morphological defects in zebrafish embryos
We first assessed the gross morphological defects in zebrafish (Danio rerio) embryos exposed to fipronil over the course of 5 days postfertilization. Embryos at 3 dpf exhibited gross morphological differences in both body length and heart size (Fig. 2). Embryos exposed to fipronil at 0.7, 1.0, and 1.5 μM had significantly shorter mean body lengths at 3, 4, and 5 dpf compared to the control embryos (Fig. 3, n = 75, p < 0.05 compared to control). There was no significant difference in mean body length between treatment groups of 0.7, 1.0, and 1.5 μM fipronil (Fig. 3). We assessed the mortality of embryos exposed to fipronil from 2 to 5 dpf and found no significant difference in mortality rate at these concentrations (data not shown).

Whole body morphological defects in zebrafish embryos exposed to 0, 0.7, 1.0, and 1.5 μM fipronil at 3 dpf. Note the shorter body length in treated embryos. Edema can be seen in the highest concentration treatment. Scale bar is 500 μm.
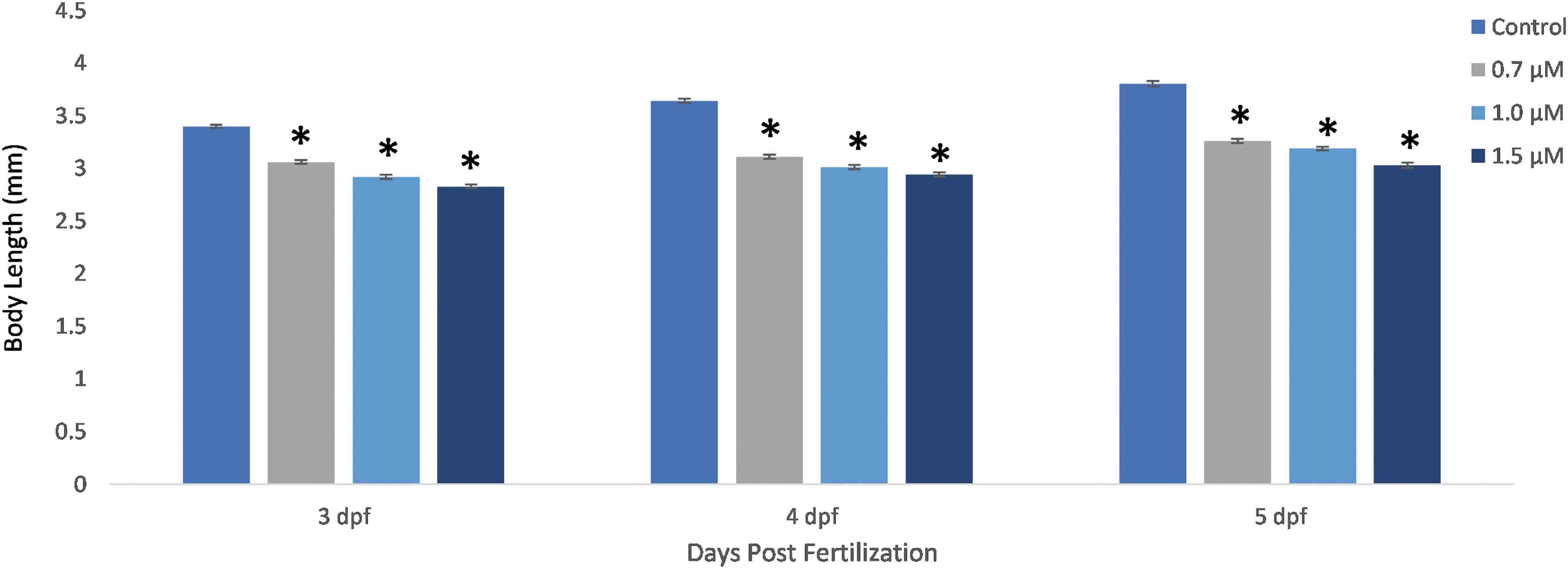
Mean body length of 3, 4, and 5 dpf Danio rerio embryos decreased significantly when treated with fipronil (p < 0.05). n = 75 embryos in each treatment (three replicates of 25 each, *significantly different from control for each day, p < 0.05).
Fipronil affects cardiovascular development in zebrafish embryos
We next investigated the effects of fipronil on zebrafish cardiovascular function and structural development. There was a significant increase in the presence of edema in the 1.5 μM fipronil treatment group compared to the control (Fig. 4A, n = 75, p < 0.05 compared to control). The heartrate of embryos was next assayed after 5 days of treatment at 5 dpf. The mean heartrate was significantly increased in the 0.7, 1.0, and 1.5 μM fipronil treatments groups compared to the control (Fig. 4B, n = 75, p < 0.05 compared to control). There was no difference in heart rate between fipronil treatment groups (Fig. 4A, B).

Heart function is significantly impacted in D. rerio embryos treated with fipronil (p < 0.05).
Structural changes in cardiovascular development were also examined at 5 dpf (Fig. 5). The total mean heart area was significantly greater when embryos were exposed to 0.7 and 1.5 μM fipronil compared to the control mean heart area (Fig. 5A, n = 60, p < 0.05). There was no difference in mean heart area between the 1.0 μM fipronil treatment and the control (Fig. 5A). The mean area of the atrium was also examined. Only the 0.7 μM fipronil treatment group had a significantly larger mean atrium area compared to the control (Fig. 5B, n = 60, p < 0.05). There was no difference between the 1.0 and 1.5 μM fipronil treatments and the control for average atrium area (Fig. 5B). In the 0.7 μM fipronil treatment group, the average atrium area was 112% larger than the control. The total heart area was 111% larger than within 0.7 μM fipronil-treated embryos and 108% larger within 1.5 μM fipronil-treated embryos compared to the control.

Fipronil affects craniofacial development in zebrafish embryos
Finally, the craniofacial development of zebrafish was investigated for ocular and gnathic changes. There was a significant decrease in mean eye area and diameter between the 0.7, 1.0, and 1.5 μM fipronil treatments groups compared to the control at 3, 4 and 5 days postfertilization (Fig. 6, n = 75, p < 0.05 compared to control). The jaw morphology was investigated via Alcian blue staining as it allows for accurate measuring of viserocranium cartilage. We examined Meckel's angle, the ceratohyal (CH) angle, the palatoquadrate (PQ) angle, the PQ to CH angle, and the PQ to Meckel's angle, along with the PQ length and the length from the ceratohyal CH to Meckle's cartilage.

There was a significant increase in the mean CH-Meckel's length in the 0.7 μM fipronil treatment group compared to the control (Fig. 7F, n = 30, p < 0.05). There was no difference between the 0.7 μM fipronil treatment and the control for the other jaw measurements (Fig. 7A–E). The 1.0 μM fipronil treatment resulted in increased mean PQ-Meckel's length, CH angle, Meckel's angle, and CH-Meckel's length compared to the control (Fig. 7A, C, E, F, n = 30, p < 0.05). There was no difference between the 1.0 μM fipronil treatment and the control for PQ length ad PQ-CH angle (Fig. 7B, D). Only the highest fipronil concentration of 1.5 μM resulted in significant jaw developmental differences of all lengths and angles compared to the control (Fig. 7A–F, n = 30, p < 0.05). These subtle differences can be visualized with Tg(sox10:gfp) zebrafish expressing GFP in the developing jaw (Fig. 8).
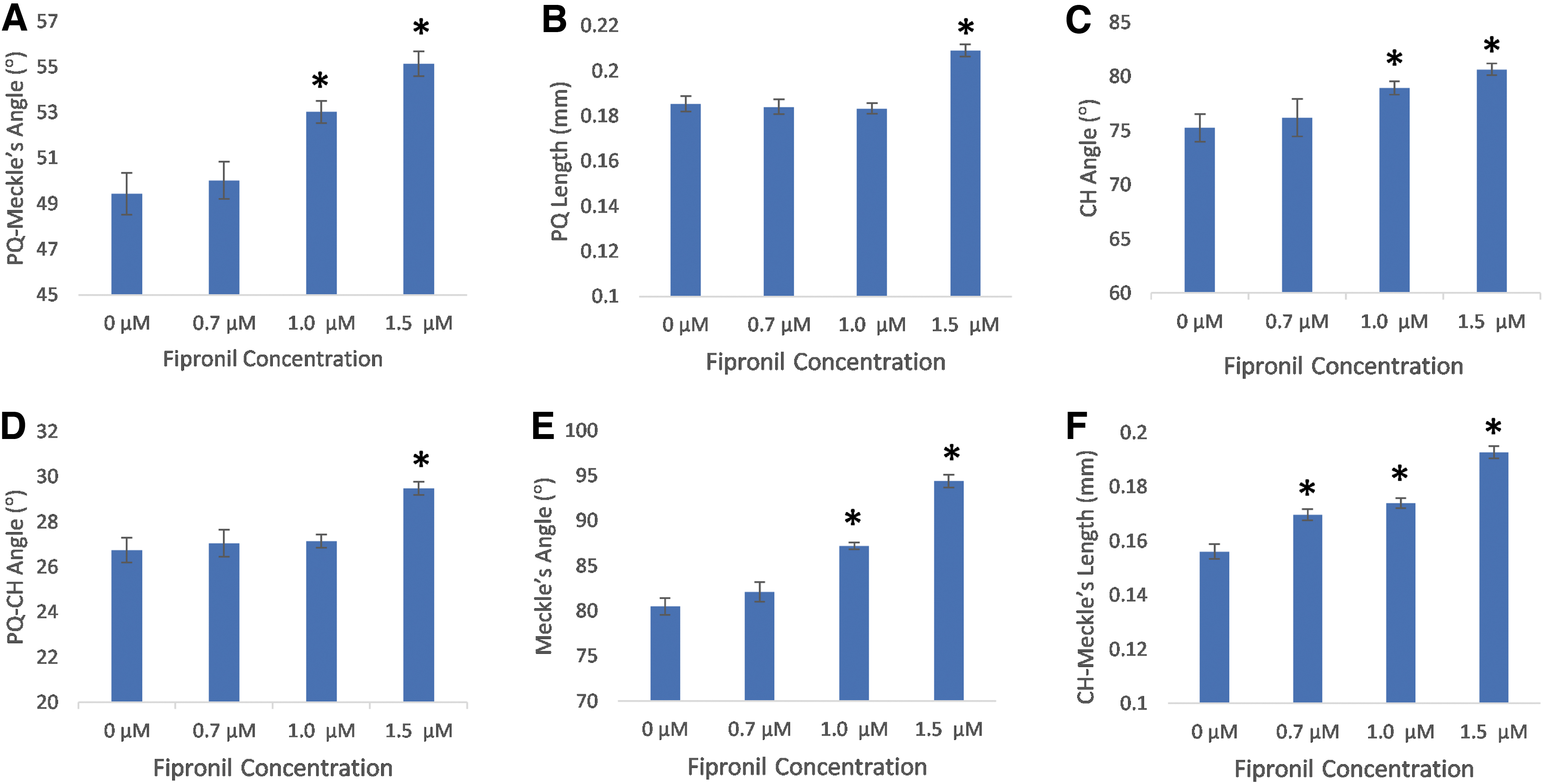
Average jaw angles and lengths of 5 dpf D. rerio embryos are significantly increased after treatment with fipronil.
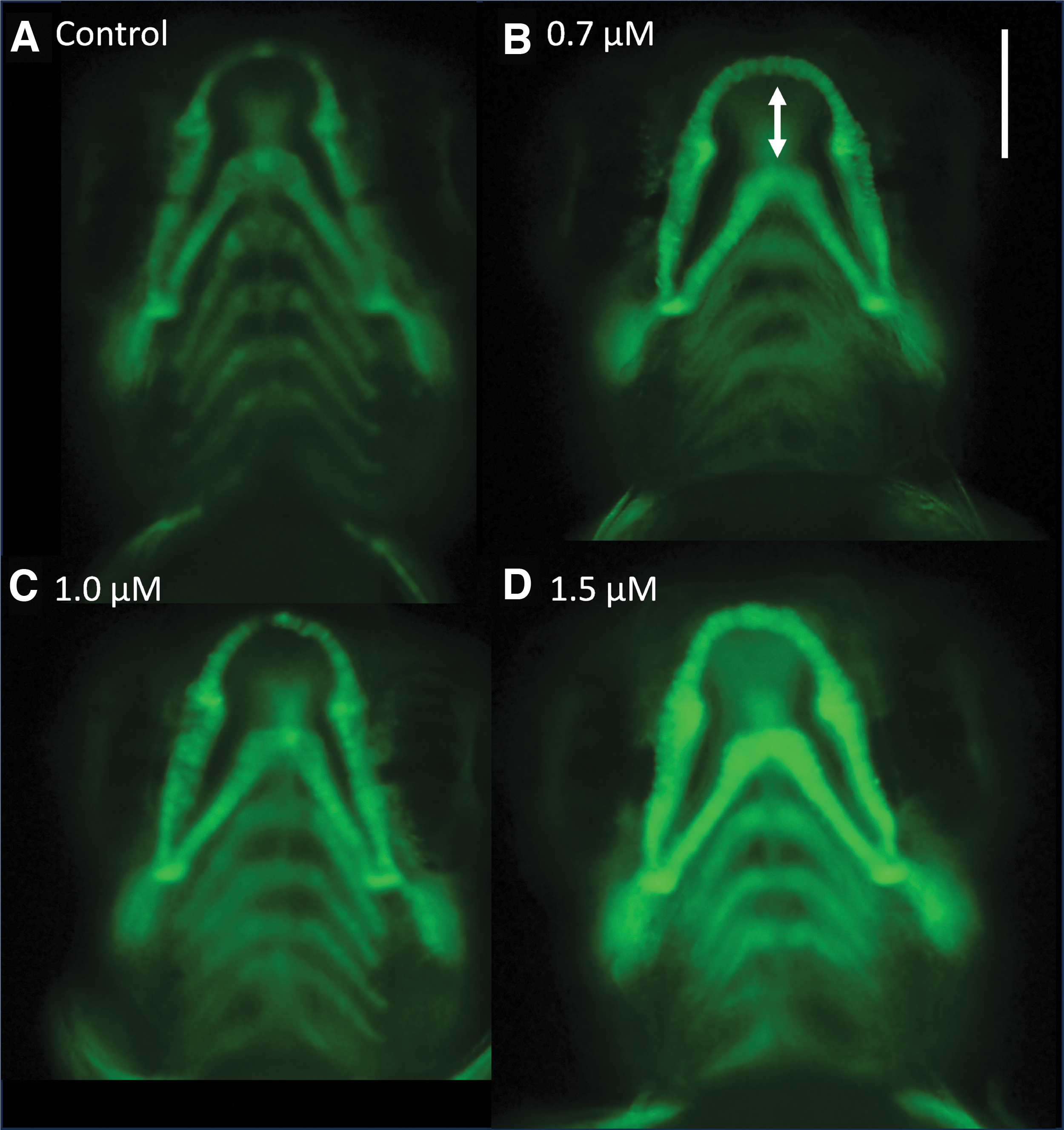
Subtle changes in jaw development in 5 dpf D. rerio embryos visualized using Tg(sox10:gfp).
Discussion
Fipronil, a common agricultural and residential insecticide, targets the invertebrate nervous system through antagonistic GABA receptor binding, causing continuous central nervous system signaling and results in the death of the pest. Fipronil was reported to be selective to invertebrates, but recent studies have shown the adverse effects of fipronil in nontarget vertebrates as fipronil and its degraded metabolites can interact with the structurally similar GABAA receptor, a ligand-gated ion channel GABA receptor.26–28
In this study, zebrafish embryos exposed to concentrations of 0.7, 1.0, and 1.5 μM fipronil displayed numerous developmental abnormalities of the body axis, cardiovascular, and craniofacial systems. The highest concentration of 1.5 μM fipronil caused a decrease in body length and eye area and an increase in total heart area, heart rate, rate of heart edema, and jaw angles and lengths. Lesser concentrations of fipronil at 0.7 and 1.0 μM caused a less severe decrease in body length and eye area, with only some jaw lengths and angles being impacted, indicating a dose–dependent relationship. These deformities reveal both a structural and functional impairment of important organ systems in the presence of fipronil.
Zebrafish exposed to fipronil had a shorter body axis with increasing concentrations, causing greater decreases in body lengths. Several other studies have reported similar decreases in body length after treatment with fipronil at 3 or 4 dpf,16–18 including a notable increase in intersegmental vessel number. 18 A spinal curve phenotype was observed in our study at high concentrations, corroborating similar observations of fipronil causing spinal curvatures in Japanese medaka 29 and zebrafish. 19 Mortality rates were not significantly different when embryos were exposed to fipronil in our study, though others treating at higher concentrations have found an increase in mortality.17,18
Fipronil also caused cardiovascular deformities. All concentrations of fipronil resulted in an increase in the prevalence of pericardial edema and an effect on heart rate, similar to previous studies.16,17 Interestingly, others have found fipronil decreases heart rate in zebrafish embryos at 3 dpf at concentrations of 6–34 μM, 17 whereas our study demonstrated an increased heart rate at 5 dpf at 0.7–1.5 μM. If fipronil is affecting glycine receptor signaling pathways, those pathways or the particular glycine receptors being expressed may change as the heart matures in zebrafish larvae.
At a concentration of 0.7 μM fipronil, atrium area was significantly greater, leading to a significantly greater total heart area. At a concentration of 1.5 μM fipronil, there was no difference in atrium area, but there was a significant increase in overall heart area. An increase in ventricle size may be contributing to the overall heart area increase. Additionally, the increase in total heart size and increase in heart rate indicates an increase in cardiac output and cardiac dysfunction. Similar studies found an increase in total heart area and increased intersegmental vessel numbers with higher concentrations of fipronil.17,18
Importantly, our results suggest that fipronil causes decreased eye area and specific jaw morphological defects. Previous studies also suggest that some pesticides, including fipronil, can decrease eye size and optic diameter.16,17,30 We found that eye area decreased in a dose-dependent manner with increasing fipronil concentrations. Additionally, analysis of jaw length and angle malformation was reported. At a concentration of 1.5 μM of fipronil, all jaw lengths and jaw angles were increased, with lesser deformities observed in the 0.7 and 1.0 μM concentrations of fipronil treatments. These findings provide specific measurement of jaw lengths and angles adding to previous studies that have reported a general truncation of the lower jaw. 16
These results show the marked effect of fipronil on zebrafish embryo development. However, the exact mechanism of action causing these effects in vertebrates is still unknown. The mechanism of action for fipronil in insects involves inhibition of insect GABA receptors. 2 Chloride channels in neurons of the central nervous system are regulated by the inhibited GABA receptors. 31 Fipronil antagonistically binds to the GABA receptor, which induces a stable nonconducting conformation of the chloride channels. The inhibition of neuronal chloride channels results in convulsions, increased excitement of the central nervous system, and death in insects. 32 The GABA mechanism has demonstrated a degree of selectivity for insect GABA receptors. 2 However, the structural similarities between vertebrate and insect GABA receptors poses a possible explanation for the results of this study.
Fipronil's impact on zebrafish development may also be due to inhibition of glycine receptors. As part of the ligand-gated chloride family, glycine receptors are structurally related to GABA receptors. The structural similarities allow for the inhibition chloride channels by fipronil acting as an antagonist to glycine receptors. 2 Fipronil's inhibition of glycine receptors is not exclusive to zebrafish and has been demonstrated in humans. Human glycine receptors α1, α1β, α2, and α3 were all inhibited by fipronil with the pesticide binding to both a pore and nonpore site on the α1 glycine receptor. 33 The demonstrated teratogenic capability of fipronil on vertebrates affirms the importance of continued research into the effects of fipronil developmental biology. Further studies are needed to investigate the mechanism of fipronil's impact on cardiovascular and craniofacial development that was demonstrated in this study.
Conclusion
In conclusion, fipronil impacts the embryonic development of zebrafish (Danio rerio). Gross morphological differences as well as malformations of the cardiovascular and craniofacial systems were observed. Dosing of 1.5 μM fipronil for 5 dpf caused a decrease in body length while also causing an increase in heart area, heart rate, eye area, jaw angles, and jaw lengths. Lower doses of fipronil at 0.7 or 1.0 μM showed some deformities to the aforementioned organ systems though not as drastic, indicating a dose–dependent relationship. This article provides novel evidence for the dose–dependent effects of fipronil on heart chamber and craniofacial morphology. Since fipronil use is still common, understanding the unintentional ecological implications of its continued application is critical. More studies are needed to understand the full effect of fipronil on both environmental welfare and human health. Future work is needed to elucidate the precise signaling mechanism causing craniofacial and cardiovascular abnormalities after fipronil treatment in zebrafish embryos.
Footnotes
Acknowledgment
The authors would like to thank members of the Lassiter Lab for zebrafish husbandry including Jacob Barrett, Olivia Brichter, Frank Marino, Stephen Miller, Austin Parker, Hannah Petty, Jill Van Doren, and Madalyn Wheeler.
Authors' Contributions
K.L.C.: Kasey started her undergraduate work at Roanoke College and is currently an undergraduate student majoring in mathematics and biology at Ursinus College. She contributed to the collection of cardiovascular data, overall data analysis, and the writing of the article.
Z.G.K.: Zoe has a BS in biochemistry and public health studies from Roanoke College and an MS from Cedars-Sinai Graduate School of Biomedical Sciences. She contributed to eye, body length, and heart rate data, and she assisted in reviewing and editing.
B.D.F.: Bennet has a BA in public health studies from Roanoke College and is currently an MD candidate at Louisiana State University School of Medicine in New Orleans, Louisiana. He did measurement and analysis of the jaw data.
B.S.W.: Benjamin has a BS in biology from Roanoke College, an MD from Virginia Tech Carilion School of Medicine, and completed the Internal Medicine residency program at Virginia Tech Carilion. He helped conceptualize the experiment and did initial literature review as well as jaw, eye, and length data collection.
A.G.K.: Alexander has a BS in biology from Roanoke College and a MS in aquaculture/aquatic sciences from Kentucky State University. He helped conceptualize the experiment and did initial literature review as well as jaw, eye, and length data collection.
J.R.M.: Jonathan has a BS in biology from Roanoke College and an MBA from the University of Maryland. He helped conceptualize the experiment and did initial literature review as well as jaw, eye, and length data collection.
C.S.L.: Christopher has a BS in biology from Furman University and a PhD in genetics from Duke University. He is a professor of biology at Roanoke College. He provided supervision, mentorship, resources, and assisted in reviewing and editing.
Disclosure Statement
The authors have no conflicts of interest to declare.
Funding Information
Funding for this project was from the Roanoke College Summer Scholars program and the Roanoke College Biology Department.