
Abstract
Over the years, scientific research with fish models has grown at a rapid pace, and issues such as animal welfare are becoming increasingly important in various areas of animal husbandry and experimentation. Here, we evaluated whether Danio rerio behavior is affected by long-term maintenance (75 days) in an enriched environment or a chronic stress (CS) situation. In addition, we evaluated some biochemical parameters related to redox status. We concluded that long-term maintenance of zebrafish in enriched environment might induce an anxiety-like behavior pattern when these fish are faced with an acute subsequent stressor. These anxiety results, the increased school cohesion, and the absence of oxidative damage allow us to hypothesize that the fish maintained in environmental enrichment (EE) situation is more reactive, showing a strong protective reaction to the stress. From an applicable perspective, we show that both too much stress and too little stress are not ideal for zebrafish stocks. In CS situations, fish can habituate and might not respond optimally to test conditions. In opposite, the low stress promoted by environmental enrichment also renders the fish incapable of dealing with occasional stressors optimally, because now even normal conditions appear stressful to them and may elicit fear behaviors they normally would not exhibit.
Introduction
Over the years, the use of fish models in scientific research has grown. In this context, issues such as animal welfare are becoming increasingly important in various areas of animal husbandry and experimentation. 1 Fish welfare research is at a less advanced stage compared with terrestrial animals. Thus, this is a promising area to explore, for example, the assessment of the effects of different welfare conditions that fish are maintained. Adequate management in research facilities, associated with good maintenance and welfare conditions, contributes to the health of animals, 2 consequently providing better and more accurate scientific results.3,4 On the opposite, low levels of welfare can harm the animals, both behaviorally and metabolically. Long-term or chronic stress (CS) is a state that can induce physiological changes, including in the central nervous system (CNS), leading to behaviors such as anxiety, 5 as well as impaired memory and learning. 6
Stress permeates endocrine and cellular responses, including oxidative pathways, which may increase the production of free radicals and influence their elimination.7,8 Antioxidant enzymes are part of the enzymatic antioxidant system and act against cellular oxidative changes, removing reactive species and preventing the establishment of a condition known as oxidative stress. One of the main consequences of oxidative stress is damage to proteins and lipids, which directly affects cell permeability. 9 Furthermore, the effects of oxidative stress can be observed in entire systems, and when decompensated or uncontrolled, it can lead to irreparable damage to the CNS.
The enzymatic antioxidant defense system plays an important role in avoiding damages induced by oxidative stress. The activity of the enzymes superoxide dismutase (SOD), catalase (CTL), and glutathione-S-transferase (GST) can show how the antioxidant system is working, showing a real image of the redox state. 10 If these enzymes are not functioning well or are not enough to prevent oxidative damage, we can observe damages in the cell such as lipid peroxidation. Lipid peroxidation is readily accessed by detecting the thiobarbituric acid reactive substances (TBARS). 11 Therefore, as oxidative stress can lead to damages in the cells of the CNS, changes in the redox profile can directly influence the physiological well-being of individuals.
One of the ways to improve the welfare level of the animals and help them better cope with stressful situations is environmental enrichment (EE), which offers animals greater control over the environment, thereby improving their welfare status. There is ample evidence that EE improves the welfare of fish such as zebrafish12–14 and Nile tilapia. 15
In this sense, understanding the impacts of welfare conditions on adult fish can make a difference in animal experimentation. In short, whatever species they are, fish in research facilities must receive management that promotes good welfare. Here, we evaluated whether improved and worsened welfare conditions of adult zebrafish interfere with the behavioral repertoire or biochemical changes, serving as an indicator of the quality of management and welfare. In addition, we aimed to evaluate whether these maintenance situations can interfere with the results obtained from these animal models in subsequent experiments.
Materials and Methods
Study strategy
Here, we compare the control, EE, and CS maintenance conditions in adult zebrafish. Operationally, we kept fish for 75 days in these three conditions and, after this period, submitted them to behavioral tests. Afterward, we used the fish tissues to determine their redox status by analyzing antioxidant enzymes and lipid peroxidation.
Zebrafish husbandry
From a mixed-sex (50 male: 50 female) stocking population, we used a total of 144 adult zebrafish older than 8 months and of reproductive age in the experiments. The fish were of wild phenotype, of unknown origin, but born and raised in the Fish Physiology Laboratory at the University of Passo Fundo for three generations.
Fish were kept in 45-L glass aquaria (50 × 30 × 30 cm, length × width × height with a 28 cm of water column) in a closed system (without recirculation), with constant aeration (by air stone) and biological filtration (hang-on biological filter, Sunsun HBL-501, 400 L/h, WilTec, Eschweiler, Germany). The temperature of 26°C–28°C was maintained by a thermostat, and an artificial photoperiod of 14 h (light)/10 h (dark) was used in the experimental room. The density was 1.5 L of water per fish, totaling 21 individuals per tank, with 14 females and 7 males (sexed visually by observing phenotypic characteristics). Food was provided twice a day (8:00 am and 5:00 pm) with flakes (Alcon Basic, Alcon, Camboriú, Brazil) and Artemia sp. nauplii. The physicochemical conditions of the water were checked periodically, once a week (pH = 7.0 ± 0.4, total ammonia < 0.01 mg/L, total hardness at 50 ± 5 mg/L, and alkalinity at 40 ± 3 mg/L calcium carbonate), always maintaining the appropriate standard. Cleaning and removal of dirt and feces were carried out three times a week, with 25% water exchange.
This experimental setup (Fig. 1) was conducted twice, and data were collected from fish pooled from these two experimental trials.

Schematic representation of the experimental design.
Procedures
Fish were kept for 75 days in three different welfare conditions, with two aquariums containing 21 fish for each experimental group. The first condition was the control, where fish were kept in appropriate maintenance and management conditions according to the species (as recommended in The Zebrafish Book 16 ). The second condition was the EE, where fish were kept under the same conditions as the other groups, but the environment was enriched with gravel/stone substrate, polyvinyl chloride pipes for refuge, and artificial plants. 14 The third condition was the CS protocol, in which fish were kept under the same conditions as the control group but were subjected daily, always at the same time, to different stressors as described in Table 1. Fish from the CS group were subjected to a daily episode of stress for 75 days, always at the same time (7:00 am), as described in Table 1. At the end of the 10th stressor, the 1st stressor was started again and so on. At the end of the 75 days of exposure to CS, fish were subjected to behavioral tests and then euthanized for biochemical determinations.
Procedure of the Chronic Stress Protocol
Behavioral tests
After 75 days of maintenance under different conditions, fish from the three groups were subjected to the Novel Tank Test (NTT), Social Preference Test (SPT), 17 and Shoaling Test (ST). 18 All behavioral tests were conducted in the morning, between 8 and 11 am. After the behavioral tests, each fish was euthanized with ice, and the spinal cord was cut, and then they were frozen in liquid nitrogen 19 to perform the biochemical tests. LED white bulbs provided light intensity in the experimental units’ room (9 W, 6500 K, 900 lumens).
NTT and SPT
Behavioral analyzes of NTT and SPT were performed according to the methodology described by Kirsten et al. 17 We used glass aquariums (24 × 8 × 20 cm, length × width × height, with an 18 cm of water column) filled with chlorine-free water for both sequential tests. For the NTT, the fish were filmed for 6 min using a Logitech c920 HD webcam camera positioned on a tripod in front of the test aquarium. The videos were analyzed using the automated tracking software ANY-maze® (Stoelting Co., Wood Dale, USA), in which the test tank was divided into the top, middle, and bottom zones. The behavioral parameters evaluated were the absolute turn angle (◦), time immobile (s), time in the top zone (s), latency to top (s), and time in the bottom zone (s). After filming each fish, the water was renewed. After NTT, two opaque barriers were removed from each side of the aquarium, and the exposed fish could see a group of conspecifics on one side and an empty aquarium on the other. The fish were left in acclimatization for 30 s after the removal of the opaque barriers. After that, the test was started, and the fish were video recorded for 1 min. SPT was carried out according to Gerlai, 20 and the parameters evaluated were the time spent in the conspecific segment (s), latency to enter the conspecific segment (s), and the time spent in the empty segment (s). After NTT and SPT, fish were euthanized in an ice bath and sent for biochemical analysis. The number of animals used for both the NTT and the SPT was 12 animals each.
ST
ST was performed as described by Miller and Gerlai 21 and adapted by Tamagno et al. 18 in an acrylic aquarium (34 × 34 × 34 cm [w × h × d]) filled with 10 cm of water with the same characteristics as the experimental aquarium. Four fish from each group were placed in the aquarium and were kept for 1 min during the acclimatization period. After that, the fish were video recorded for 6 min in a way so that we could see the back of each fish composing the shoal. Each video was then separated into 12 photos, each of 30 s. The distance between all four fish was measured (a total of 6 measures per picture) in each picture using ImageJ Software, and the final value was presented as the distance in centimeters among the fish in a shoal. We analyzed 45 animals in this test.
Biochemical tests
For the biochemical tests, fish were ice-anesthetized and dissected. The whole brain was removed from the skull cap, and three brains were pooled per sample. In addition, the muscle of the one ventral side of the fish (0.05 g) was removed and homogenized per sample. The homogenization was carried out in an ice bath using a potter homogenizer for 1.5 min by adding 800 uL of 50 mM Tris-HCl, pH 7.5. 22 The protein content was quantified at the beginning of the tests, and all the calculations followed the protein concentration. The biochemical tests performed were the levels of malondialdehyde (MDA) by the TBARS technique, which was quantified in muscle and brain. The activities of the antioxidant enzymes CTL, GST, and SOD were also quantified in both tissues. Lipid peroxidation was carried out by quantifying the concentration of weak bases that reacted with thiobarbituric acid. 23 The activity of the enzyme CTL was measured by the conversion of hydrogen peroxide under 240 nm.24,25 The activity of GST was measured by the rate of conversion of reduced glutathione by dinitrobenzene.26,27 The SOD activity was performed by the quantification of the epinephrine inhibition. 28
Statistical analysis
Data from the three experimental groups were analyzed by one-way analysis of variance followed by Tukey’s post-hoc test or Kruskal–Wallis test followed by Dunn’s test, depending on the data normality (assessed by the Kolmogorov–Smirnov test). Graphs were constructed using GraphPad Prism 8.0.1 software. Each graph is represented by mean ± standard error.
Results
Behavioral tests
NTT
The absolute turn angle (Fig. 2A) was reduced in the CS group compared with the control and EE (F2,37 = 11.71; p < 0.0001). Immobile time (Fig. 2B) was increased in the CS group compared with the EE (H = 6.291; p < 0.05). The time spent in the top zone (Fig. 2C) was reduced in the EE group compared with the control and CS groups (F2,39 = 5.315; p < 0.01). The latency to enter the top (Fig. 2D) was unchanged in any of the exposure groups (H = 0.1739; p > 0.05). The time spent in the bottom zone (Fig. 2E) was increased in the EE group compared with the control and CS groups (F2,37 = 7.322; p < 0.01).
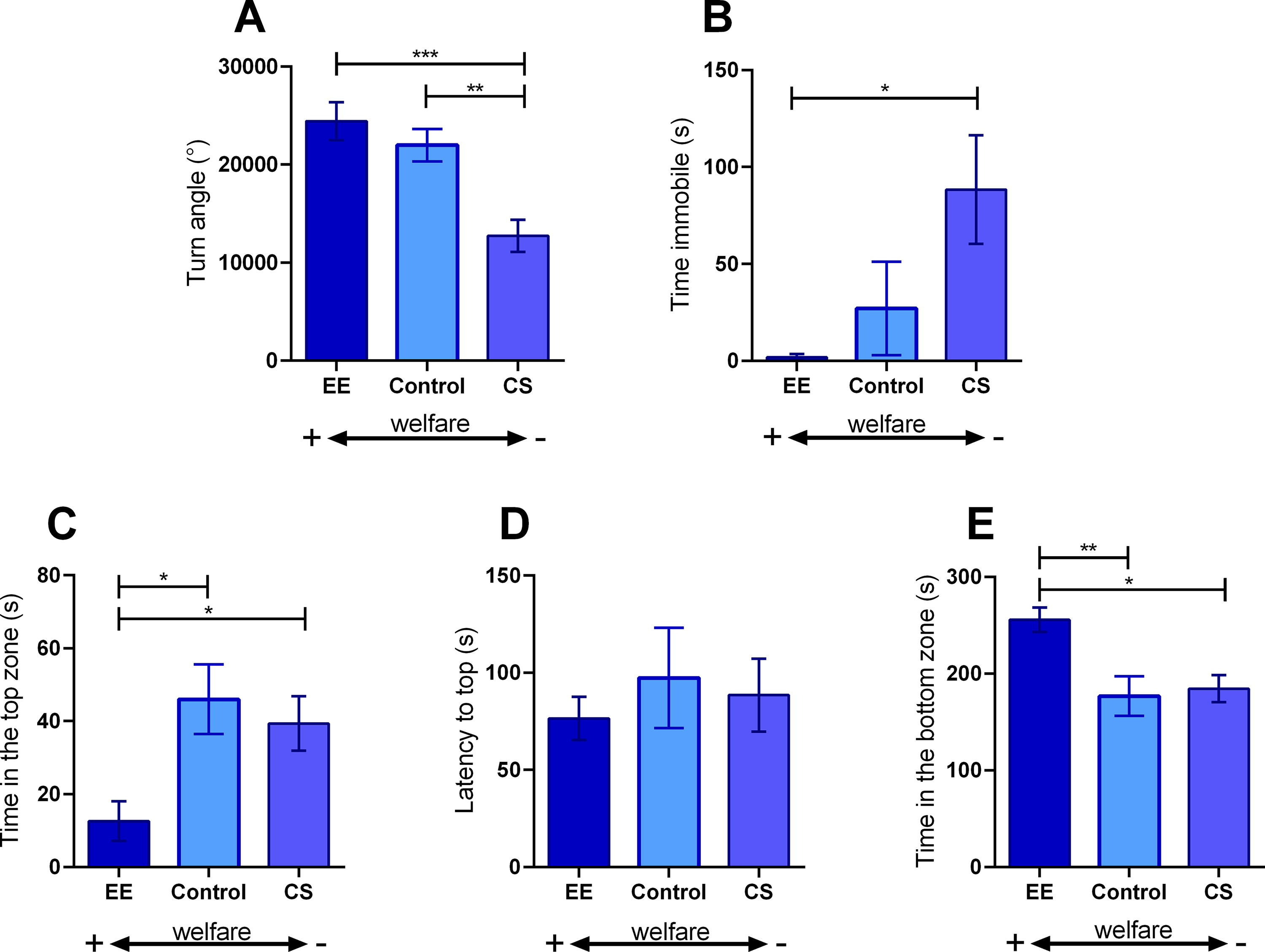
Novel tank test (NTT) of fish kept for 75 days in standard conditions (control), enriched environment (EE), or under chronic stress (CS). Data of absolute turn angle
SPT
No significant alteration was observed in the comparisons between the exposure groups (Table 2).
Social Preference Test of Fish Kept for 75 Days Under Standard Conditions (Control), Enriched Environment, or Chronic Stress
Data expressed as mean ± SEM analyzed by ANOVA followed by Tukey’s post-hoc test.
ANOVA, analysis of variance; CS, chronic stress; EE, enriched environment; SEM, standard error of mean.
ST
The distance between the individuals in the shoal was relatively smaller in the EE group compared with the control and CS groups (Fig. 3; H = 35.11; p < 0.0001).
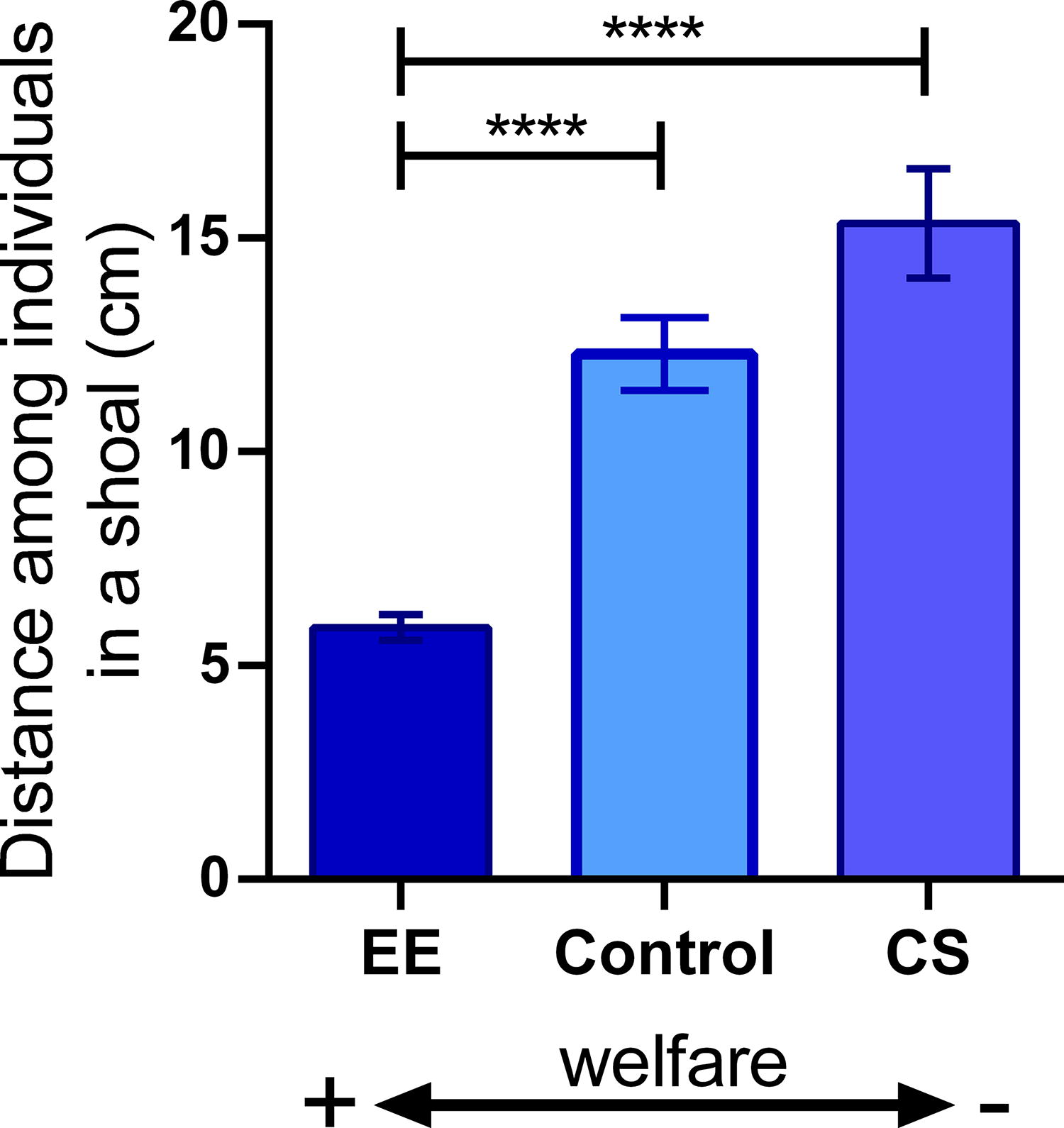
Shoaling test (ST) of fish kept for 75 days under standard conditions (control), enriched environment (EE), or chronic stress (CS). Data expressed as median ± interquartile range (panel B) and analyzed by Kruskal–Wallis test followed by Dunn’s test (n = 9–14). ****p < 0.0001.
Biochemical analyzes
Status redox
The TBARS levels were decreased in the muscle (Fig. 4A) of CS fish when compared with the control and EE fish (H = 12.89; p < 0.01). In brain tissue (Fig. 4E), MDA levels were decreased in the group maintained on EE compared with the control and CS (H = 9.926; p < 0.01).

Redox status of fish kept for 75 days under standard conditions (control), enriched environment (EE), or chronic stress (CS). TBARS levels in muscle
In CTL activity in muscle tissue (Fig. 4B), no change was observed between exposure groups (F2,45 > 1; p > 0.05). In the brain (Fig. 4F), CTL activity was increased in the group maintained in the CS condition compared with the control and EE group (H = 13.96; p < 0.001).
The GST activity in the muscle (Fig. 4C) was decreased in the EE group compared with the control and CS groups (H = 22.24; p < 0.0001). In the brain (Fig. 4G), none of the exposure groups had statistically significant changes (F2,38 = 2.064; p > 0.05).
Finally, SOD enzyme activity in muscle (Fig. 4D) was increased in the group maintained on CS compared with the EE group (H = 7.351; p < 0.05). Brain SOD activity (Fig. 4H) was increased in the CS group compared with the EE group (H = 7.215; p < 0.05).
Discussion
In this study, we evaluated whether D. rerio behavior was affected by long-term maintenance (75 days) in an enriched environment or a CS situation. In addition, we evaluated biochemical parameters related to redox status.
Regarding the behavioral responses, in the NTT, we observed that EE fish presented locomotor parameters (total distance, absolute turn angle, and time immobile) indistinguishable from the control fish. However, EE fish spent less time in the upper zone of the tank and more in the bottom, characterizing an anxiogenic behavior phenotype. Analyzing the social behavior, both EE and CS were similar to control data in the SPT. However, in the ST, EE strongly increases shoal cohesion, as seen by a three-fold reduction in the distance between fish (Fig. 3).
Our main hypothesis to explain these behavioral results is that EE fish experienced fewer stressful situations during the 75 days and responded more actively to the subsequent acute stress provoked by the NTT. 29 Reinforcing this hypothesis, the CS fish react less in the NTT, probably because of the modulation caused by the CS on the subsequent acute stress caused by the test. This modulation was verified in other fish species as the Nile tilapia 30 and the Atlantic salmon (Salmo salar). 31 Other important factors that reinforce our hypothesis are the absence of locomotor alterations in EE fish, the strong social response of EE in ST, and the reduced lipid peroxidation. Reduced locomotion and impaired social behaviors are part of the stress behavior phenotype of zebrafish and are observed in CS fish in our study and in the scientific literature.22,32–34 The absence of oxidative stress in EE fish corroborates the behavioral data, as there was no destabilization of the redox state at the brain level, which explains such a robust behavioral effect. In addition, behavioral responses to stress are greatly influenced by the environment, as environmental factors affect behavioral responses to stress. 35
These behavioral responses of EE fish seen in the NTT and ST are essential both at individual and population levels. NTT evaluates the locomotor and exploratory behavior when fish are faced with a novel environment. 29 The changes observed in EE fish in the NTT (anxiogenic) demonstrate the intact ability of these individuals to demonstrate an anxious behavior in the face of stress provoked by the test, in a way it is expected that healthy fish demonstrate different behavioral changes that guarantee their survival in the face of different stress conditions. 36 In line with the NTT results, EE fish presented a marked response of increasing the school cohesion in the ST (Fig. 3). This effect is a classical protective behavior.37,38 The zebrafish, as a social species, use shoaling not only to interact with their conspecifics but also to protect themselves from predators. 37
Regarding the possible mechanisms underlying both EE and CS fish responses to the behavioral tasks, we evaluated the oxidative status of fish.
Intrinsically related to the central effects demonstrated, changes in the enzymatic antioxidant system of zebrafish were observed under EE and CS conditions. The decrease in lipid peroxidation may be an attempt to combat reactive oxygen species (ROS). SOD and CTL were decreased, possibly because they may be below the detection limit and probably, they have already acted on combating ROS and therefore decreased lipid peroxidation levels, and when dosed, possibly already returned to baseline levels. On the contrary, for prevention of initial damage, it can be assumed that they stabilized since they were evaluated after permanence in EE, but there was not enough oxidation to have peroxidation in lipids, possibly due to adaptive effects or even prevention. 7
As commented earlier, EE fish that were kept in improved conditions, with a safe and peaceful environment, presented an anxiety-like pattern, which can be interpreted as a strong reaction to a stressful and challenging NTT environment. 29 In opposite, the CS fish were stressed every day and adapted to a routine of stress challenges. Therefore, these animals were not anxious and, theoretically, did not respond as well to the stress imposed by the behavioral tasks, as they had already adapted to a sequence of stressors. 35
Regarding the GST enzyme activity, in the muscle of CS fish, the activity of the enzyme was increased. These changes can be detrimental to an individual’s second line of antioxidant defense. GST acts by inserting glutathione groups into nonpolar toxic compounds to transform them into polar ones, thus facilitating their excretion. 39 In the present study, no toxic compound responsible for the alteration of the GST was administered; therefore, stress not only plays a crucial role because of the stress response but can also influence the organism’s normal physiological response to a toxicant. Therefore, if under environmental conditions, individuals come into contact with toxic compounds, their physiological response may be largely affected because of stress, something that is not observed under EE conditions, 10 reinforcing this hypothesis.
Taken together, the antioxidant biomarkers and lipid damage are closely related. The EE seems to be able to prevent lipid peroxidation in the brain. Thus, our results strongly suggest that EE was crucial for the prevention of oxidative damage, cognitive maintenance, and preservation of the natural behavioral repertoire. On the contrary, and fostering this hypothesis, in CS fish, there was the formation of peroxidized lipids in the brain, even with the increase in antioxidant enzymes, which seem not to have been enough to prevent cell damage.
In addition, another possible explanation for the CS group presenting increased SOD, CTL, and GST activity could be due to the increased production of ROS that is influenced by the fast temperature increase 40 used as one of the stressors. In addition, the opening of sodium ion channels is observed in severe temperature increases, which can highlight the acetylcholinesterase increase mainly in the muscles due to high influx of serum ionized calcium 41 in the neuromuscular junction and exacerbated release of acetylcholine in the synaptic cleft.
Our study has some possible limitations. First, we did not measure the physiological stress response (i.e., cortisol levels) after behavioral testing to confirm the stress caused by the CS protocol. A second limitation is that we did not consider the sex of fish in the results, not confirming the visual sexing in euthanized fish. However, it is worth noting that fish are typically maintained in sex-mixed stock tanks, aligning with the primary focus of our experiment. A last limitation is that we did not take intermediate assessment time points and, thus, cannot evaluate whether the fish habituate to CS. However, these technical aspects do not negate the overall relevance of our results.
A final comment pertains to the definition of EE, which emerges as a way to improve animal welfare by promoting control over the environment, reducing the chance of negative experiences such as fear and frustration, and consequently improving their degree of welfare. 42 Mounting shreds of evidence show that EE improves fish welfare, including in zebrafish12–14 and Nile tilapia. 15 Zebrafish is an animal model widely used in research, and the maintenance conditions are very important to generate good results, highlighting the environment in which the fish are kept, which also influences the behavior of the zebrafish. 43 In the present work, the main result is the clear anxiety-like behavioral pattern in EE fish, which we explain by the maintenance of proper welfare conditions. As these fish do not face previous stress, they become more sensitive and reactive to the subtle acute stress imposed by the behavioral tasks. The present results also reinforce our previous work, 43 showing EE as an interfering factor in the research results.
We concluded that fish kept in EE become more susceptible to anxiety-like behavior when exposed to a stressful stimulus. Combined with the unchanged locomotor parameter, the increased school cohesion, and the absence of oxidative damage, we hypothesized that the EE fish is more reactive, showing a strong protective reaction to stress, which may be translated as the maintenance of the natural behavioral repertoire of the species. From a practical point of view, we demonstrate that both too much stress and too little stress are not ideal for zebrafish stocks. In stressful situations, fish habituate to CS and might not respond optimally to test conditions. On the contrary, conditions with low stress, as promoted by environmental enrichment, also render the fish incapable of dealing with occasional stressors optimally because even normal conditions now appear stressful to them and may elicit fear behaviors they normally would not exhibit.
Footnotes
Acknowledgments
The authors would like to thank the Coordenação de Aperfeiçoamento de Pessoal de Nivel Superior (CAPES), the University of Passo Fundo, and the Federal Institute of Rio Grande do Sul, Campus Sertão, for providing the laboratory structure and funding for this research.
Authors’ Contributions
F.U.I.A.: Conceptualization, Methodology, Writing—Reviewing and Editing, Data Curation, Formal Analysis, Investigation, and Project. A.P.: Data Curation, Formal Analysis, and Writing—Original Draft. P.A.D.A.: Methodology, Writing—Reviewing and Editing, Data Curation, and Formal Analysis. N.F.: Methodology, Writing—Reviewing and Editing, Data Curation, and Formal Analysis. L.D.D.S.D.P.: Methodology, Writing—Reviewing and Editing, Data Curation, and Formal Analysis. W.A.T.: Methodology, Writing—Reviewing and Editing, Data Curation, and Formal Analysis. G.R.: Formal Analyses. A.L.R.V.: Methodology. J.A.D.L.: Methodology. H.C.D.S.: Methodology. A.G.: Methodology. L.J.G.B.: Supervision, Writing—Original Draft, Writing—Reviewing and Editing, and Funding Acquisition.
Ethics Approval
This study was approved by the Ethics Committee for the Care and Use of Animals of the University of Passo Fundo under protocol number 004/2022. In addition, this study was registered in the Sistema Nacional de Patrimônio Genético e do Conhecimento Tradicional Associado and complied with their guidelines (registration code A14E252).
Availability of Data and Material
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This study was supported by