
Abstract
Hydrocephalus is a cerebrospinal fluid-related disease that usually manifests as abnormal dilation of the ventricles, with a triad of clinical findings including walking difficulty, reduced attention span, and urinary frequency or incontinence. The onset of congenital hydrocephalus is closely related to mutations in genes that regulate brain development. Currently, our understanding of the mechanisms of congenital hydrocephalus remains limited, and the prognosis of existing treatments is unsatisfactory. Additionally, there are no suitable or dedicated model organisms for congenital hydrocephalus. Therefore, it is significant to determine the mechanism and develop special animal models of congenital hydrocephalus. Recently, zebrafish have emerged as a popular model organism in many fields, including developmental biology, genetics, and toxicology. Its genome shares high similarity with that of humans, and it has fast and low-cost reproduction. These advantages make it suitable for studying the pathogenesis and therapeutic approaches for various diseases, specifically congenital diseases. This study explored the possibility of using zebrafish as a model organism for congenital hydrocephalus. This review describes the characteristics of zebrafish and discusses specific congenital hydrocephalus models. The advantages and limitations of using zebrafish for hydrocephalus research are highlighted, and insights for further model development are provided.
Introduction
Hydrocephalus is a common neurological disorder characterized by abnormal accumulation of cerebrospinal fluid (CSF) with or without an increase in intracranial pressure.1,2 Moreover, it can be classified as congenital hydrocephalus (CH) or acquired hydrocephalus. The former is typically caused by genetic factors, such as mutations in L1CAM, MPDZ, CCDC88C, and other related genes. 3 The latter is often triggered by infections, inflammation, traumatic injury, intracranial hemorrhage, and other associated factors. Genetic defects leading to CH often result in ciliary abnormalities, blood–brain barrier (BBB) disruptions, and structural changes in brain ventricles and other regions. CH may lead to macrocephaly, cognitive impairments, and even fatality. Currently, the standard clinical approach for treating CH involves reducing the CSF volume through neurosurgical shunting. However, these treatments may yield suboptimal outcomes. 4 To enhance the therapeutic approaches and prognoses of CH, a comprehensive investigation into establishing a suitable model organism holds significant promise.
Zebrafish is currently a widely used model organism in the study of congenital neurological disorders because of its high genome similarity to humans, rapid reproductive capabilities, and cost-effectiveness. Zebrafish can undergo external fertilization, and their embryos are transparent, allowing for direct microscopic observation of tissue and organ development, blood flow, heartbeats, and other physiological indices. Moreover, zebrafish can readily use CRISPR/Cas9 gene editing technology, facilitating the generation of precise mutants tailored for specific research objectives.5,6 It has also been considered a promising newcomer in behavioral neuroscience. 7 However, in the current study, using zebrafish as a model organism, we noticed that hydrocephalus is frequently regarded as a complication of other congenital diseases (scoliosis or renal cysts). No research has been dedicated to the establishment of zebrafish models for CH.
Consequently, this article provides an overview of zebrafish model organisms with complications of CH, focusing on their pathogenesis, complications, advantages, and limitations. This study aimed to gain a better understanding of the application of zebrafish in the CH. Through this approach, we hope to establish a foundation for developing a stable and specific zebrafish model of CH, which can subsequently contribute to the study of its pathogenesis and crucial therapeutic targets. Ultimately, this research endeavors to enhance treatment approaches for CH and strive to improve its prognosis.
Hydrocephalus and Existing Animal Model Types
Hydrocephalus is a neurological disorder caused by abnormal accumulation of CSF in the brain ventricles. Excess fluid widens the ventricles, increases intracranial pressure, and has harmful effects on the brain tissues. Hydrocephalus may occur shortly after birth or may result from damage or injury in adults. It is difficult to determine exactly how many people suffer from hydrocephalus because it occurs at all ages and can develop later in life. Some researchers report one to two of every 1,000 babies born with CH, an estimated 400,000 new cases of pediatric hydrocephalus each year globally, and ∼1.5% of the population (more than 5% of individuals older than 80 years) suffer from idiopathic normal pressure hydrocephalus (iNPH), with a higher prevalence in men than women.1,8–10 Although there are many people of all ages suffering from hydrocephalus in the world, the pathogenesis remains unclear. However, some scientific hypotheses are significant for understanding the generation of many hydrocephalus, such as the theory of circulating flow, penetration, glymphatic system, and others.
According to the theory of circulating flow, the pathogenesis of hydrocephalus can be divided into several specific ways. One is obstruction, such as the mass effect of the tumor near the posterior cranial fossa, blocking the circulating pathway of the CSF, and making it abnormally build up in the ventricles. The other is ciliary defects (including cilia genesis and movement). Genetic mutations or inflammations can lead to the development or injury of the ciliary ependymal cells lining the surface of the ventricles, which determines the flow direction of the CSF. Besides, brain hemorrhage, inflammation, and infection may destroy the choroid plexus, resulting in excess CSF secretion, which finally leads to hydrocephalus. Impairment of the BBB is also an important factor in hydrocephalus, which can be affected by congenital and acquired reasons.
The theory of penetration emphasizes that the brain parenchyma is permeable, and the cause of hydrocephalus may also be due to the accumulation of hypertonic substances in the ventricle and the transport disorders of water and other substances in the CSF. Channel proteins such as AQP4 and NKCC1 and CSF substances such as ferroprotein have all been found to be linked to hydrocephalus.11,12
Recently, a correlation between the glymphatic system and hydrocephalus was reported. The glymphatic system, first identified by Maiken et al. in 2012, is a unique extracellular space clearance system in the brain. 13 It performs its clearance function through the exchange of substances between the brain interstitial fluid and the CSF in the perivascular space. Damage to the glymphatic system has been confirmed in patients with iNPH. Moreover, inflammation, stroke, and brain trauma can affect the function of the glymphatic system, resulting in hydrocephalus.14,15
Besides the theories mentioned above, gene mutations that disrupt brain morphogenesis alter the biomechanics of the CSF-brain interface, leading to abnormalities in the brain ventricle and can also lead to infant hydrocephalus. 1
Due to our limited recognition of hydrocephalus, suitable animal models should be established. Other researchers have attempted to develop hydrocephalus models in rats, mice, rabbits, dogs, and pigs for adult posthemorrhagic hydrocephalus (PHH). Hydrocephalus is a severe complication of intraventricular hemorrhage (IVH), and many PHH animal models are based on it. 16 These animal models can be divided into four main categories: (1) primary IVH and hydrocephalus, (2) IVH secondary to intracerebral hemorrhage and hydrocephalus, (3) IVH secondary to traumatic brain injury and hydrocephalus, and (4) subarachnoid hemorrhage and hydrocephalus. Autologous blood, iron, or thrombin are used to trigger hydrocephalus in mammalians via intraventricular, striatum, basal ganglia injection, lateral fluid-percussion, or endovascular perforation. 17
Although there are some animal models of PHH, model organisms of other types of hydrocephalus are very limited, particularly those of CH. CH is thought to be associated with genes involved in Sylvius aqueduct defects, cilia growth and movement, and nervous system development. However, the numbered models were mainly produced by infection, such as attenuated influenza A virus vaccine in Rhesus monkeys, and did not target the specific meaningful genes. 18 Some gene defect CH models are established, such as Cwh43, Wdr16, and Wdr78, but the main purpose was to research CH, and these models have obvious other diseases.19,20
Consequently, further research into the development of effective animal models of CH will facilitate research on the pathogenesis of CH, specifically in the field of drug prevention and therapy.
Advantages and Applications of Zebrafish as a Model Organism
Recently, zebrafish (Danio rerio) has emerged as an immensely valuable animal model for many medical research. The genomic similarity between zebrafish and humans exceeds 70%, with over 80% of human disease-related genes can be found in the zebrafish genome. Therefore, zebrafish can be utilized to investigate pathogenic mechanisms in many diseases.6,21,22
Zebrafish models for ophthalmology
Diseases of the photoreceptor cells can lead to irreversible vision loss and blindness in humans; however, effective therapies for these patients are lacking. The development of suitable animal models of cone disease is challenging because many of the popular model animals used are nocturnal, which means that they are rod-dominant visual, and their cones are dissimilar to human cones. However, zebrafish, whose retina is rich in cone cells, provide an opportunity to mimic photoreceptor disease in diurnal vertebrates. 23 Aged zebrafish retinas (>30 months) exhibited age-related changes similar to those observed in humans. 24 Zebrafish develop many pathologies and various types of photoreceptor diseases (developmental, rod, cone, and mixed) resembling macular degeneration. 25 Compared to other model organisms, zebrafish are better adapted for investigating single-cell transcriptomics (SCT) during initial development. Many SCT studies (not including zebrafish) that analyze ocular components are limited to very few or only one developmental stage because of the adverse effects of postmortem extraction. 26 However, zebrafish can overcome these difficulties due to their high reproductive rate and rapid initial development (the basic components of the eye develop within 5 days after fertilization). 27 Besides, they are well-suited for multi-timepoint SCT analyses and subsequent functional gene investigations of vertebrate development. 28 Consequently, zebrafish offer distinct advantages for research in the field of ophthalmology due to the conservation of their eye morphology, physiological tissues, and regulatory signaling pathways.28–30 The applications of zebrafish ophthalmology disease models are summarized in Table 1. As animal models for ophthalmology, zebrafish models with a single copy of a functionally conserved gene accurately reproduce the human phenotype, such as the pde6c or rho mutant lines. Although some lines have phenotypes that differ from humans (for example, the cep290 mutation causes cone-specific degeneration in a zebrafish model, whereas the CEP290 mutation causes Leber congenital amaurosis in humans), animal models are intended to accurately reproduce human disease manifestations and to provide insights into photoreceptor function. Therefore, zebrafish provide unique opportunities to elucidate the molecular mechanisms underlying photoreceptor disease and degeneration.
Similarities and Differences Between the Nervous Systems of Zebrafish and Humans
BBB, blood–brain barrier; ChP, choroid plexus.
Zebrafish models for toxicology
Zebrafish are widely used for toxicology research from molecular mechanisms to organismal phenotype and behavior, particularly in the aspects of organ system toxicology,31,32 investigating mechanisms of action, ecotoxicity, environmental toxicants, chemical toxicity screening, and drug screening. Toxicological outcomes in zebrafish are easy to establish, including LC50, morphological and physiological parameters that indicate acute toxicity or death, such as embryo coagulation, nondetachment of the tail, lack of somite formation, absence of heartbeat, the curvature of the spine (scoliosis), pericardial and yolk sac edema, and alterations in the swimming bladder. 33 Besides, zebrafish embryos and larvae are transparent. In some transgenic zebrafish lines, fluorescent proteins are activated in the presence of contaminants or environmental stressors, making them easy to observe (Table 1).34,35
Zebrafish models for developmental biology
As a model organism, zebrafish have close developmental trajectories of many systems with humans, and the formation of some structures is similar to that of mammals (such as the aortic arch). Therefore, many researchers have used zebrafish as a tool in various developmental biology studies, particularly in the fields of the heart, eye morphogenesis, nervous system, and skeleton. 36 Bergmann et al. developed a new transgenic zebrafish strain (NBT: GCaMP3) that stably expressed a fluorescent calcium reporter gene in all major brain regions of larval brains up to 21 days of age, which can be used to characterize the receptor field of optic tectum neurons in the late larval stage. 37 In the field of neurodevelopmental disorders, zebrafish were used to test prenatal risk factors for autism spectrum disorder and intellectual disability. 38 The advantages of zebrafish in heart research can also be used to establish human congenital heart disease models. 39 Because of the transparency of zebrafish embryos and larvae, morphological changes in the cardiovascular system during development can be visualized by transmission light or fluorescence imaging (Table 1). These studies revealed the potential use of zebrafish models to effectively evaluate novel genetic variants, contributing to the early identification of pathogenic factors and accelerating disease diagnosis and treatment.
Zebrafish models for aberrant cognitive disease
Zebrafish possess unique characteristics as a model organism for neurological research. 40 Although the central nervous system of zebrafish originates from different neurogenic processes, it shares similar tissue organization with mammals, displaying the same main brain structures, such as the forebrain, midbrain, hindbrain, and spinal cord (Fig. 1A and Table 1). Additionally, at the cellular level, zebrafish and mammals exhibit parallels, exemplified by astrocytes, ependyma cells, and cerebellar Purkinje cells (Table 1).51–53 In zebrafish, the observation of hydrocephalus is notably more straightforward, allowing for confirmation through multiple indicators, such as ventricle transparency and volume and magnetic resonance imaging. 54 Currently, a range of imaging techniques have been employed to investigate the pathophysiology of brain-related disorders in zebrafish. These techniques include fluorescence microscopy, laser scanning microscopy, transmission electron microscopy, and high-throughput imaging systems.55–57
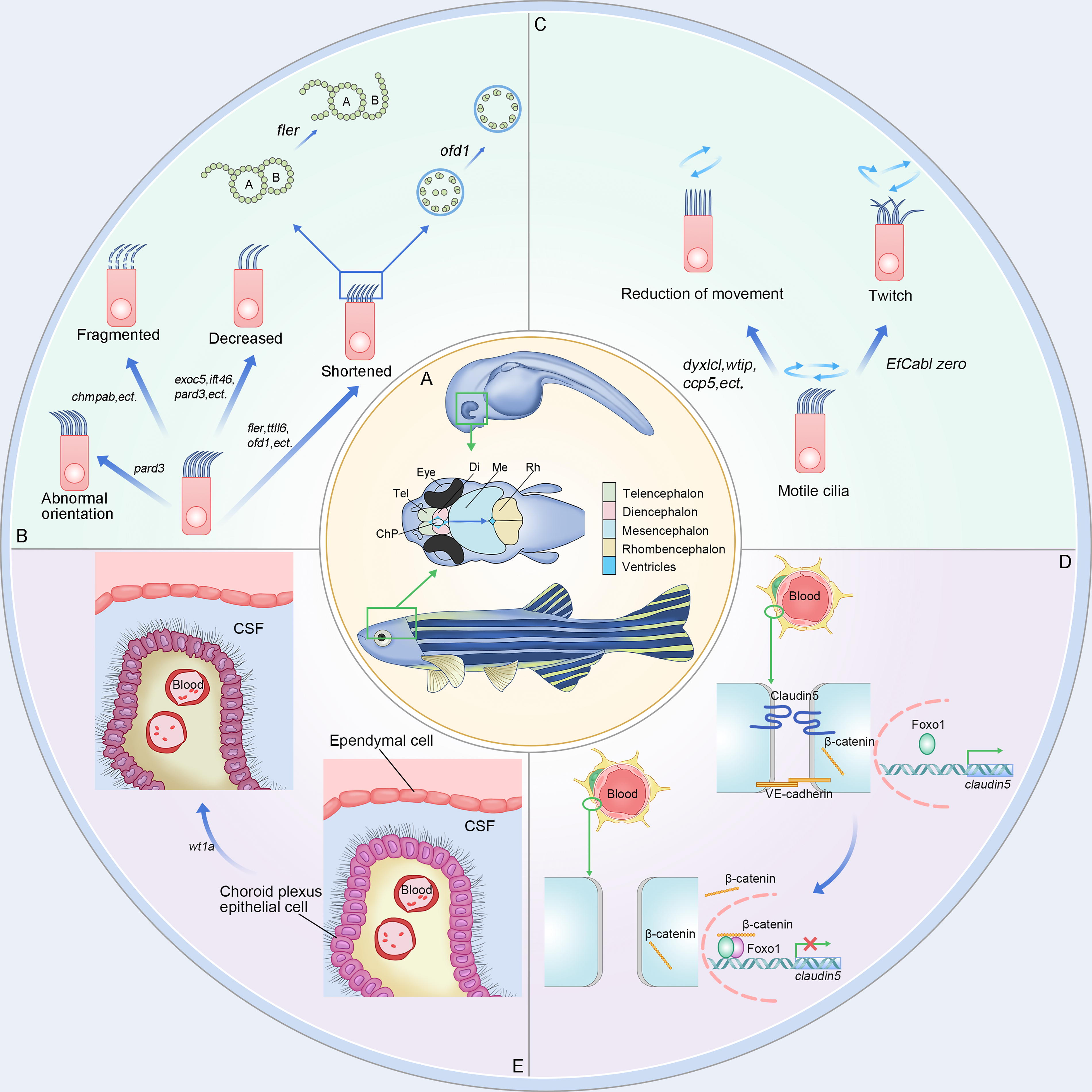
Pathogenesis of zebrafish CH.
Besides, many behavioral tests that have been used to study cognitive phenotypes in rodents, such as the open-field test, the light-dark box, and the T- or Y-maze, have been successfully applied to zebrafish. 58 Zebrafish can recognize geometric figures and form spatial memories. 59 These novel findings and breakthroughs indicate the potential of zebrafish for studying aberrant cognitive and neurodegenerative diseases. To date, several orthologs of major disease-associated genes have been found in zebrafish, including Alzheimer’s disease, attention deficit hyperactivity disorder, epilepsy, Huntington’s disease, Parkinson’s disease, schizophrenia, and others (Table 2). Transgene, knockdown, knockout, or overexpression of mutants of these genes suggests that they have conserved functions in the development and survival of aberrant cognitive disease and neurodegenerative diseases.
Applications of Zebrafish as Model Organisms
Molecular Targets of Congenital Hydrocephalus in Zebrafish Models
BVS, brain ventricular system.
Applications of Zebrafish as a Model Organism Related to Hydrocephalus
Hydrocephalus in zebrafish and human
Zebrafish serve as a potential model organism for studying hydrocephalus. The hydrocephalus phenotype in zebrafish typically manifests as ventricular enlargement, head swelling, reduced eye size, decreased pigmentation, weakened blood circulation, and pericardial edema. These phenotypes can be observed in the early developmental stages (20 hpf) and worsen over time. Detection methods include microscopy, gene expression analysis (such as in situ hybridization or qPCR), advanced imaging techniques (MRI or CT), and behavioral tests to assess neurological function. 59 The pathophysiology of hydrocephalus in zebrafish is similar to that in humans, involving disruption of CSF circulation, absorption, or overproduction. 85 At the cellular and developmental levels, ependymal cells in zebrafish and humans play crucial roles in maintaining CSF flow and ventricular structure, with ciliary dysfunction leading to CSF flow abnormalities. Rapid development and transparent embryos make zebrafish ideal for studying the dynamic changes in hydrocephalus during development, providing insights into the mechanisms and potential therapeutic approaches.
Cilia are hair-like structures that extend from the cell surface and are unique in eukaryotic organisms. They consist of a matrix, axoneme, and ciliary membrane. The axoneme, which is mainly composed of microtubules, serves as the main structure of cilia. 86 Cilia can be categorized into two forms: motile and nonmotile cilia. Motile cilia move in a regular rhythm and direction, while nonmotile cilia serve as sensory organelles. Numerous studies have suggested that cilia dysfunction can disrupt the circulation of CSF, potentially contributing to the development of CH. Ciliary defects are primarily caused by genetic mutations. Presently, several zebrafish models with CH phenotypes are designed based on ciliary abnormalities (Fig. 1B, C). Here, we summarized these zebrafish models with ciliary defects and classified them into two subtypes: ciliogenesis and cilia movement defects (Table 3).
Zebrafish model with CH caused by ciliogenesis defects
Ciliary generation defects can be broadly classified into two categories: alterations in ciliary morphology and number. Currently, there is an increasing emphasis on investigating the pathological repercussions arising from alterations in ciliary morphology (Fig. 1B).
Kramer-Zucker and his colleagues initially explored the impact of ciliary morphology on the organ development and intraorgan fluid dynamics of pronephros, brain, and Kupffer’s vesicles in zebrafish. Subsequently, they identified that defects in ciliary morphology or motility could also contribute to the onset of CH. 119 Subsequently, researchers discovered that mutations in zebrafish genes, such as exoc5/sec10, 87 cdc42,88,89 and ift46, 43 were associated with truncated or absent cilia in the choroid plexus, ultimately contributing to hydrocephalus. Knockout of the wdr16 gene disrupts osmotic regulation and brain water balance, causing severe hydrocephalus, which is also correlated with cilia. 20 Afterward, research has delved deeper, shifting the focus toward the study of the structural components of cilia, particularly microtubules. In 2010, E. Hong elucidated that, during neurogenesis, microtubules and the polarity protein collaboratively regulate the positioning of centrosomes. Pard3 deficiency in zebrafish can induce aberrant cilia growth and orientation, eventually leading to the development of CH. 90 This study revealed a novel molecular mechanism associated with the pathogenesis of CH and provided valuable insights for developing novel zebrafish models of this condition. Notably, in contrast to the other models, there were nearly no complications in the pard3 mutation CH zebrafish models. In 2020, pertinent research further advanced to the protein subunit level. The knockout of chmp4b, which codes for a subunit with the function of endosomal sorting complexes, was found to interfere with cilia assembly. It even triggers the fragmentation of existing cilia, resulting in the onset of hydrocephalus. 92
In zebrafish, besides the fact that several gene defects related to ciliary generation have been identified as causative factors for CH, other studies have revealed a correlation between the number of cilia and CH. In studies on the correlation between microtubule protein polyglutamylation and cilia formation in zebrafish, knockout of ttll6 (responsible for microtubule protein polyglutamylation) was observed to induce CH. 41 This study substantiates the significance of polyglutamylation of microtubule proteins in cilia generation and motility. Then, N. Pathak and colleagues extended this investigation and conducted a more detailed analysis of the expression patterns of ccp during zebrafish development. Their findings revealed that among the ccp family, ccp1 expression is confined to the nervous system. Only the knockout of the ccp5 gene led to elevated glutamylation of cilia microtubule protein, triggering ciliary dysfunction and ultimately manifesting as a CH phenotype. 46
Additionally, studies have revealed that mutation of wtip, encoding the LIM domain protein WTIP, can result in the functional impairment of ccp1 and ccp5, culminating in a decrease in ciliated cells. 44 These studies have established a framework for identifying ciliary genes pertinent to the nervous system.120,121 Furthermore, studies have pointed out that the combined knockout of the calb2a and calb2b genes, transient knockdown of cdc42, sec10, dcdc2, and ccdc28b, as well as reducing the expression of nherf1a/slc9a3r1, collectively lead to the diminished genesis of cilia and severe CH in zebrafish.88,93–96 Among these, the dcdc2 knockout and low expression of nherf1a/slc9a3r1 are related to the classical Wnt signaling pathway, which is important in the development of CH. Overexpression of dcdc2 inhibits β-catenin in the Wnt pathway and modulates the pathogenesis of hydrocephalus. Similarly, nherf1a/slc9a3r1 also functions through the Wnt/PCP pathway.
Despite the conventional concept linking CH to cilia defects disrupting the flow of CSF, recent research indicates an additional correlation between brain ventricular anomalies and motile cilia development. Specifically, gmnc and foxj1a mediate the transition from solitary ciliated cells to multiciliated cells, resulting in an enlarged brain ventricle and CH. 98
Although the above studies have successfully constructed a zebrafish model of CH (except for the model of E. Hong), hydrocephalus only appears as a complication of other prominent conditions (such as kidney cysts, asymmetrical malformations, curved bodies, and others). Consequently, the CH zebrafish model designed for ciliary generation defects is in its nascent stage. To address these multifaceted complications, it is necessary to find a specific gene exclusively affecting the brain cilia to refine CH zebrafish model organisms.
Zebrafish model with CH caused by cilia movement defects
In most circumstances, genes affecting cilia generation have adverse effects on cilia movement. For instance, wtip, gmnc, and ccp5 reduce cilia number, morphology, and motility. Except for the mutations and abnormalities mentioned above, alterations in cilia number, structure, and the absence of basal roots can all impact ciliary motility (Fig. 1C). In zebrafish, four tetratricopeptide repeat-containing (TTC) proteins (ttc4, -9c, -30, -36, and -39c encoded) are crucial for cilia formation and motility. 122 Defects in these proteins can cause CH. Notably, hydrocephalus, attributed to impaired ciliary motility, always correlates with obstruction of CSF flow.
However, alterations in ciliary motility do not come just from the three factors mentioned above. Various genes governing ciliary motility still exist. For instance, the knockout of the dynein arm protein-encoding gene dnaaf3 in zebrafish disrupts the assembly of dynein arms and ciliary motility, ultimately leading to the development of hydrocephalus. 100 In 2013, the study conducted by G. Chandrasekar and his team investigated the tetrapeptide repeat domain-containing protein DYX1C1, a factor associated with rodent neuronal migration. Researchers have discovered that dyx1c1 is expressed across multiple ciliated tissues in zebrafish. Zebrafish with dyx1c1 mutations exhibit deficiency in both the outer and inner arms, resulting in ciliary motility disorders and subsequent CH. 45 Similarly, within the same year, the research highlighted that the knockout of two genes, bbs1 and nphp7.2, both influencing ciliary motility in zebrafish, leads to hydrocephalus. 101 Furthermore, in 2019, it was found that the calaxin mutation in zebrafish triggers irregular ciliary beating and inversion, inducing abnormal CSF circulation and, consequently, CH. 47
Regrettably, CH zebrafish models developed based on ciliary motility disorders still face challenges because these genes are hard to localize precisely in the brain. Consequently, these models may also lead to other typical diseases in other systems (such as kidney cysts), where cilia also play a significant role. These facts make research focusing on CH difficult.
Zebrafish model with CH caused by BBB disruption
The BBB serves as a semi-permeable protective partition that segregates circulating blood from the brain microenvironment. Its composition comprises three layers from the inside out: tightly connected brain capillary endothelial cells, continuous basement membrane, and end feet of neuroglial cells. The BBB prevents harmful substances from entering the brain microenvironment. Studies have demonstrated that disruption of the BBB, including increased permeability, is associated with the development of hydrocephalus (Table 3). Some genes are related to BBB development.
Claudin-5a, encoded by cldn5a (claudin-5), is a tight junction protein specifically located in the brain that is notably abundant at the BBB in zebrafish (Fig. 1D). Phosphorylation plays a pivotal role in both its expression and function by regulating claudin-5 expression or the protein itself. Several phosphorylation sites and activation mechanisms have been revealed to impair the integrity and tightness of endothelial cells. 48 Foxo1 and β-catenin inhibit the transcription of claudin-5 by binding to its promoter. Cldn5a gene expression can be regulated by VE-cadherin. VE-cadherin expression triggers Akt activation, subsequently leading to the phosphorylation of FoxO1. This phosphorylation limits β-catenin translocation to the nucleus, thereby alleviating its inhibitory effect on claudin-5 transcription. 123 In 2010, a zebrafish model of CH was developed by inactivating VE-cadherin. 124 This manipulation resulted in reduced claudin-5 expression, thereby causing BBB abnormalities and the emergence of hydrocephalus. Expansion of the zebrafish embryonic brain before the establishment of the embryonic BBB is essential. However, an investigation demonstrated that mutations in claudin5a prevented this procedure. The reduced brain ventricular volume was caused by a decrease in paracellular tightness of the cerebral–ventricular barrier.
Moreover, this discovery established a foundation for comprehending the collaborative influence of claudin-5a and Na+/K+-ATPase during ventricular expansion. 125 Zebrafish claudin-5a serves as a direct homolog of human CLDN5. No additional complications have arisen from zebrafish model organisms by claudin-5 modification. 126 Therefore, developing a CH zebrafish model using claudin-5a as a specific molecule is promising for further research.
The choroid plexus is also a crucial component of the BBB (Fig. 1E). The Wilms tumor suppressor gene WT1 in humans encodes a zinc-finger transcription factor. Furthermore, its paralogs in zebrafish, wt1a and wtlb, are necessary for normal choroid plexus formation and function, which explains why wt1a deficiency causes CH in zebrafish. 49 Similarly, human esophageal cancer-related gene-4 (ECRG4) and its protein Aurrin are associated with choroid plexus epithelial cell development. Knockout of the ecrg4 gene can induce a dose-dependent CH zebrafish model, and this phenotype can be reversed by coinjection of antisense morpholinos with ecrg4 mRNA. 50 Moreover, components of the classical Notch signaling pathway, such as notch1b, deltaA, and deltaD, can also mediate choroid plexus-related disorders and cause hydrocephalus in zebrafish. 127
The CH zebrafish models mentioned above, based on the BBB, exhibit relatively high specificity. However, the lifespan of the zebrafish is limited. For instance, wt1a mutant zebrafish survive for <10 days. Furthermore, many studies have focused on zebrafish embryos, leaving uncertainties regarding the feasibility of surviving into adulthood for further investigative purposes. Accordingly, it reminds us that the lifespan should also be considered when developing CH zebrafish models.
Zebrafish model with CH caused by brain ventricular system abnormalities
The brain ventricular system (BVS), surrounded by ependymal cells, is formed by interconnected cavities filled with CSF. BVS is vital for regulating neurogenesis, brain function, and internal homeostasis. Dysregulation of the BVS can lead to CH.8,128,129 Brain ventricular expansion in hydrocephalus is primarily caused by an imbalance between the production and reabsorption of CSF or obstruction of its flow. Moreover, patients with CH manifest additional structural brain anomalies, suggesting that hydrocephalus may not solely arise from CSF homeostasis disruption but could also be due to the abnormal development of the brain parenchyma. 130 Current research indicates that BVS-related genes are associated with embryonic brain development, ventricular structure, brain vascularization, cell adhesion, and brain epithelial cells (Table 3).
Nestin is a type IV neurofilament protein. Zebrafish nestin is a direct homolog of the mammalian counterpart NES and is broadly expressed in the developing period of the nervous system. Studies have revealed that nestin knockdown in zebrafish embryos prompts developmental anomalies of the brain and ocular structures, leading to microcephaly and CH. During embryonic development, these zebrafish embryos manifest hindbrain and midbrain shrinkage symptoms, accompanied by ventricular enlargement. 105 The effect of nestin on brain development is dose-dependent, implying the possibility of ameliorating embryonic microcephaly and CH by partially moderating nestin expression levels. This study also suggests the potential of developing an adult CH zebrafish model.
Previous studies elucidating the etiology of hydrocephalus have suggested that iron accumulation could influence the Wnt pathway. 131 The pantothenate kinase 2 gene (PANK2) in humans and its paralog pank2 in zebrafish are expressed in the brain, and their mutations are closely related to pantothenate kinase-associated neurodegeneration.132–134 Notably, research reveals that pank2 is also vital for brain vascular formation, and the downregulation of pank2 causes an abnormal zebrafish brain structure and CH. 106
The impact of L1CAM on the ventricular structure has been studied for a long time in humans. As early as 1995, researchers found that human L1CAM mutations can cause ventricular expansion, intellectual disability, corpus callosum agenesis, and hydrocephalus. 135 Subsequent research illustrated that L1CAM LOF mutations can cause human X-linked hydrocephalus. 136 The absence of these genes in Xenopus laevis and mice also causes hydrocephalus.137,138 In zebrafish, l1cam exists in dual forms, namely l1cama and l1camb. Chl1a closely resembled l1cam. Adam10 codes a protease that orchestrates the detachment of the L1CAM protein from the cell surface by regulating the FNIII domain. 108 Zebrafish chl1a demonstrates 46% homology to humans and governs ventricular system development and cell adhesion. Notably, chl1a-deficient zebrafish embryos have Reissner fiber formation deficiency, resulting in obvious hydrocephalus symptoms. 107
Thioredoxin (encoded by thioredoxin 1) is a 12 kDa intracellular oxidoreductase. It plays a pivotal role in protecting cells and tissues against oxidative stress-induced apoptosis through its active center cysteine residue,139,140 impending the proapoptotic function of signal-regulating kinase 1 via ubiquitination and degradation and regulating the transcription of angiogenic genes by blocking NF-κB activation.141–143 Knockdown of thioredoxin 1 amplifies apoptosis in ventricular epithelial cells, culminating in hydrocephalus. However, mouse models reveal embryonic lethality upon this knockout. To establish a CH zebrafish model targeting thioredoxin 1, a survival-permitting strategy must be devised. Notably, environmental chemicals such as arsenic, cadmium, mercury, 1,1-bis-(4-chlorophenyl)−2,2,2-trichloroethane (DDT), and 2,3,7,8-tetrachlorodibenzo-p-dioxin affect thioredoxin expression by predominantly diminishing its mRNA levels. 109 As a result, a zebrafish model for CH could be developed by diminishing thioredoxin expression via chemical exposure in zebrafish embryos.
Recent investigations have revealed the significance of voltage-gated potassium (Kv) channels in the emergence and maturation of BVS, which is attributed to their pivotal role in cell proliferation. 144 Research has illuminated that silencing the zebrafish kcng4b, a homolog of the human gene encoding Kv6.4 subunit, results in the inhibition of neuroepithelial proliferation within the ventricles. This alteration is mediated by the modulation of Kv2.1 activity, which shifts neuroepithelial integrity. Therefore, ventricular expansion is induced. Consequently, zebrafish with kcng4b gene deficiency manifest the development of CH.110,145,146
Mutations in the leucine-rich glioma inactivated 1 (LGI1) gene render individuals susceptible to hereditary epilepsy syndrome in human.147–149 In zebrafish, there are two homologs of the LGI1 gene: lgi1a and lgi1b. Lgi1a anomalies are linked to epilepsy-like hyperactivity, concomitant with brain cell apoptosis,150,151 while knockdown or mutation of the lgi1b gene leads to abnormal development. It does not induce epilepsy-like behavior; instead, lgi1b triggers brain cell apoptosis and brain volume reduction, resulting in enlarged ventricles and CH. 111
BVS crucially regulates neurogenesis, brain function, and homeostasis. Dysregulation of the BVS can lead to CH, which is driven by CSF accumulation or developmental anomalies. Nestin deficiency in zebrafish demonstrates its importance in brain development and offers a potential therapeutic target for embryonic brain anomalies. The role of iron in CH via the Wnt pathway represents a novel etiological approach. Genes such as pank2, l1cam, and lgi1 influence the brain structure, providing insights into CH mechanisms. The impact of thioredoxin suggests a chemical-induced zebrafish model organism for CH, and Kv channels are linked to ventricular development. These findings enrich our understanding of CH and facilitate zebrafish model organism advancement.
Zebrafish model with CH caused by ions-related factors
The ion-transporter is crucial for the secretion of CSF and the dysregulation of the balance of Na+, Cl-, and HCO3−, and then the secretion of CSF can also lead to hydrocephalus. In zebrafish, several ion-related gene defects or mutations have been identified as causative factors of CH (Table 3). Polycystin-2 (coded by pkd2) is a cation-permeable transient receptor potential ion channel that plays a pivotal role. During embryonic development, disruptions in pkd2 expression lead to its mislocalization on apical cell surfaces, culminating in hydrocephalus. This condition is rescued by human PKD2 mRNA expression.152,153 Simultaneous disruption of the pkd1a and pkd1b genes, responsible for encoding the polycystic protein PKD1, leads to sustained upregulation of collagen mRNA expression. This alteration influences the secretion and assembly of the extracellular matrix, perturbing matrix integrity and ultimately resulting in CH.99,154 Slc41a1 downregulation induces hydrocephalus in zebrafish but is accompanied by severe kidney lesions, which are also ions-related. 113 Na+/K+ ATPase is a transmembrane ion pump crucial for ion gradients and maintains basic cellular functions. In zebrafish, atp1a3, primarily expressed in neurons, exhibits two homologs with distinct brain expression patterns: atp1a3a and atp1a3b. Knockdown of atp1a3a and atp1a3b leads to membrane ion distribution imbalance, resulting in CSF accumulation, ventricular enlargement, and finally, hydrocephalus. 114
Zebrafish model with CH caused by other reasons
Arhgef7b/βPix knockdown results in the immature development of cerebral blood vessels in embryonic zebrafish, leading to hydrocephalus and intracranial-specific hemorrhage during early embryogenesis. This particular phenotype is attributed to specific βPix splice variants in the progression of CH. 89 Both aberrant splicing of psen1 exon 8, a direct homolog of the human PSEN1 gene, and reduced PSEN1 protein activity in zebrafish can inhibit Notch signaling, resulting in obvious hydrocephalus.115,155 HRG1 mutations perturb heme balance and influence CNS development and blood flow in humans. Transient hrg-1 knockdown in zebrafish leads to CH. 116 Mutation of the heat shock protein spg gene affects cell skeleton structure, axon growth, and neuron migration during early embryogenesis, leading to CH. 156 Knockdown of ispd or gtdc2 as glycosyltransferase genes contributes to Walker–Warburg syndrome-associated hydrocephalus, which is potentially linked to brain integrity damage by muscle fiber formation defects and protein glycosylation.117,118
Various gene defects and mutations in zebrafish can contribute to CH, making zebrafish a valuable model organism for studying genetic etiology and exploring potential therapeutic targets. It is prudent to acknowledge that although zebrafish is a widely employed model organism in biological investigations, additional endeavors should be made to establish a more human-related and stable model organism.
Discussion
This study provides a comprehensive review of the pathogenesis, complications, advantages, and limitations of zebrafish as a model organism for inducing CH. In models based on ciliary defects, the hydrocephalus phenotype in zebrafish is often accompanied by severe renal complications, making it less specific. However, the degree of hydrocephalus is controllable, and zebrafish typically survive to adulthood. The CH zebrafish model organism based on BBB disruption allows for better localization to the brain with fewer complications. However, the degree of hydrocephalus is usually more serious and potentially leading to embryonic mortality. This may complicate future research. A zebrafish model based on BVS abnormalities exhibits characteristics similar to those of the BBB model but may induce other neurological disorders such as epilepsy, encephalitis, or Alzheimer’s disease, which reduces its specificity. Several other CH zebrafish models based on other mechanisms do not align with the current mainstream pathogenesis of CH and are unsuitable as a starting point for studying the pathogenesis and targeted therapies of hydrocephalus.
Regarding whether zebrafish can serve as a model organism for hydrocephalus, some factors must be considered. 157 In mammals, the BVS consists of two lateral ventricles (telencephalon) connected to the third ventricle (diencephalon) through the Monro foramen. The third ventricle is linked to the fourth ventricle via the Sylvian aqueduct, which drains CSF into the subarachnoid space and central canal of the spinal cord. Each ventricle contains a choroid plexus. 158 However, the BVS of zebrafish comprises only three cavities, and only two satisfy the morphological criteria for ventricles. However, the nomenclature of zebrafish BVS is still controversial. 159
Furthermore, the molecular mechanisms underlying choroid plexus development in mammals and zebrafish may differ. For example, while Otx2-deficient mice exhibit abnormal development of all four choroid plexus structures, zebrafish otx2 expression is restricted to the diencephalon. 160 Additionally, anatomical differences between zebrafish and mammals may render them unsuitable as animal models for certain studies, such as degenerative diseases, due to the regenerative capacity of some zebrafish cells. The significant evolutionary distance between zebrafish and humans also requires additional effort to develop functional assays to study specific human-like phenotypes.
Despite the popularity of zebrafish as a model animal, further understanding of the specific role of specific genes in hydrocephalus is required when developing CH models. A comparison of zebrafish and human protein-coding genes has revealed many interesting features. We can find that the human gene has at least one zebrafish ortholog; however, one human gene may correspond to multiple zebrafish orthologs. 6 For instance, CALB2 in humans corresponds to calb2a and calb2b in zebrafish, and WT1 corresponds to wt1a and wt1b. Notably, these zebrafish orthologs may have different biological functions. Lgi1a and lgi1b are two paralogs of human LGIL, but only lgi1b is correlated with hydrocephalus, whereas lgi1a is associated with epilepsy. Therefore, when we consider a particular gene related to hydrocephalus in humans, we should identify all its orthologs and their functions in zebrafish.
In conclusion, although many zebrafish models present hydrocephalus symptoms, their original research objectives are limited to establishing specific CH zebrafish model organisms. To better investigate the pathogenesis and targeted therapies for CH, it may be worthwhile to identify a target gene specifically localized to cilia in the brain. It can also contribute to the establishment of a more precise and controllable CH zebrafish model. Another strategy could be to identify a brain-specific gene that exclusively triggers hydrocephalus or that has minimal side effects. By modulating the expression of this gene, we can control the degree of hydrocephalus for research purposes by adjusting its expression level. While a highly specific CH zebrafish model has not yet been fully established, existing research indicates substantial potential for the development of such a model organism. Based on previous studies, it remains feasible to create a zebrafish model and offer a focused platform for investigating CH.
Conclusion
Given the limited understanding of CH, there is a compelling need for further investigation into its pathogenesis and effective treatment. This review comprehensively examines the pathogenesis, complications, advantages, and limitations of utilizing zebrafish as an animal model of CH. These CH zebrafish models are based on ciliogenesis defects, ciliary movement defects, BBB disruption, BVS abnormalities, ion-related factors, and other etiological factors. Numerous zebrafish models manifest CH symptoms, but their primary objective is not to develop precise CH models. Consequently, a highly specific CH zebrafish model has not yet been definitively established, and establishing a stable model is imperative. This refined model has the potential to enhance our understanding of the pathogenesis of CH, enable earlier prenatal diagnosis, and facilitate the discovery of targeted interventions. Ultimately, these advancements could lead to improved prognosis, reduced mortality, and decreased burdens on affected families.
Footnotes
Acknowledgments
All the researchers who have helped with this study and all the peer reviewers for their opinions and suggestions are thanked.
Authors’ Contributions
K.W.: Conceptualization, methodology, software, and writing—original draft. Z.T.: Conceptualization, methodology, funding acquisition, and writing—original draft. Y.Y.: Investigation and visualization. Y.G.: Visualization and writing—reviewing and editing. Z.L.: Writing—reviewing and editing and project administration. Z.S.: Writing—reviewing and editing. X.L.: Conceptualization, supervision, validation, and project administration. G.X.: Conceptualization, supervision, validation, project administration, and funding acquisition.
Disclosure Statement
The author(s) report no conflicts of interest in this work.
Funding Information
This work was supported by the National Natural Science Foundation of China (Nos. 82371362 and 82171347), the Hunan Provincial Natural Science Foundation of China (No. 2022JJ30971), the Health Research Project of Hunan Provincial Health Commission (No. 202204040024), and the Students Innovations in Central South University of China (Nos. XCX2022176, S2022105331024, XCX2022218, 2022105330190, and S202310533489).