
Abstract
Intestinal permeability plays a crucial role in intestinal barrier function. Altered intestinal permeability is well documented in numerous chronic diseases and may serve as a risk factor for disease onset as well as a target for innovative therapeutic strategies. While reliable and sensitive approaches for studying intestinal permeability have been established in animal models, such as mice and zebrafish larvae, methods for investigating this in adult zebrafish remain a considerable challenge. The zebrafish has emerged as a valuable model for studying intestinal development, physiology, and disease. Moreover, zebrafish offer certain advantages over rodent models, such as the ability to evaluate the dynamic interactions of labeled markers in vivo and in real time. In this study, we present a comprehensive pipeline for assessing in vivo intestinal permeability in adult zebrafish using fluorescent-labeled dextran. Detailed protocols for fish handling, reagent preparation, optimization of reagent dosage and delivery routes, and quantification of fluorescent markers in extraintestinal sites are provided. Our findings suggest that zebrafish hold promise as an alternative model for in vivo investigations of intestinal permeability induced by genetic, pathophysiological, and/or pharmacological events.

Introduction
The intestinal barrier serves as a critical line of defense against pathogens and ingested toxins. Its function is modulated by tight junction (TJ) permeability and the integrity of epithelial cells. Dysfunction of the intestinal epithelial barrier and heightened permeability have been implicated in a range of intestinal disorders.1–3 Likewise, leaky gut syndrome has been associated with higher susceptibility to a variety of extraintestinal diseases, including autoimmune conditions, metabolic, and neurodegenerative diseases.4–7 Therefore, the barrier function can serve as a crucial biomarker of disease severity and is frequently assessed in experimental and/or clinical investigations. 8
Intestinal permeability is often characterized as the measurable rate of luminal content diffusion through the mucosal barrier into the bloodstream, and it is a parameter commonly employed to evaluate the integrity of the gut barrier.4,5 Current methodologies including ex vivo assessment of electrical conductivity across excised intestinal tissues (Ussing chambers), or in vivo tests based on the administration of water-soluble probes by oral or intravenous routes, followed by subsequent identification in biological samples.4,8,9 In preclinical models with induced intestinal barrier dysfunction, the assessment of intestinal permeability is commonly performed with fluorescein isothiocyanate—conjugated dextran (FITC-D), followed by detection of fluorescence in plasma on a spectrophotometer.4,10,11
Murine models have been traditionally employed to investigate intestinal permeability. These studies have significantly advanced our understanding of the complex and multifactorial processes maintaining intestinal barrier integrity. 12 However, there is a growing need for alternative animal models that can offer additional methodologies not explored in rodent studies, thereby providing a different perspective in this field. The zebrafish (Danio rerio) provides an interest contribution to our understanding about gut development, pathophysiology, and the molecular mechanisms underpinning intestinal barrier integrity.13–18 Notably, zebrafish possess an intestinal architecture and physiology that are conserved with mammals, as well as most classical intestinal transport pathways.13–17 In some aspects, zebrafish offer advantages over rodents, such as the ability to evaluate the dynamic interaction of labeled markers in vivo and in real time.16,17 Also, zebrafish are tractable to genetic manipulation, providing comprehension about human disease. 19
Studies evaluating intestinal barrier integrity in vivo in zebrafish larvae have been previously reported.17,20,21 In the current study, we chose to use adult zebrafish due to some limitations associated with the larvae stage, such as the higher complexity of microgavage and the need for specific equipment for the procedure. In addition, the complete development of intestinal folds and intestinal cell lineage specification in zebrafish typically occurs after 2 weeks postfertilization.13,14 To the best of our knowledge, we found just one study that asses paracellular permeability in excised gut of adult zebrafish. 22 Therefore, we present an in vivo approach for evaluating gut solute paracellular permeability in adult zebrafish, which can be applied to studies requiring functional assessment of intestinal permeability in alternative models as well as in models of intestinal inflammatory disease and metabolic disorders.
Material and Methods
Ethical approval
Fish handling procedures were conducted in accordance with ethical guidelines and were approved by the Animal Research Ethics Committee of the Institute of Biosciences and the Institute of Biomedical Science at the University of São Paulo under protocol numbers 389/2021 and 8914030821, respectively.
Zebrafish husbandry
The fish were provided by the Zebrafish Facility of the Institute of Biosciences at the University of São Paulo. A total of 40 female AB zebrafish and 30 Casper zebrafish of both sexes, all 7–8 months old and full siblings, were raised at the facility following standard housing and husbandry recommendations.23,24 Zebrafish were housed in a recirculating aquaculture system at 28°C (14 h/10 h light/dark cycle). The water pH (7.5 ± 0.1) and dissolved oxygen (>6 mg.L−1) were measured daily. Water quality parameters were measured once a week using commercial kits (Labcon Ammonia Alcon Fresh Water; Labcon Test Nitrite Alcon, Brazil). Fish were maintained at density of 3 adult fish/L. All animals were fed to apparent satiety three times daily, receiving a diet comprising a mixture of Artemia and Zeigler’s Adult Zebrafish AP400 (Zeigler Bros, Germany).
Reagents setup
All salts, acids, bases, and solvents used in this study, as well as tricaine methanesulfonate (MS-222) and rhodamine B isothiocyanate–Dextran (RITC-Dextran), were purchased from Sigma Aldrich, USA. Proteases and phosphatase inhibitors to supplementation of the lysis buffer were purchased from Roche, USA. The kit to protein quantification was purchase from Thermo Scientific, USA. All solutions were prepared using deionized water (ddH2O), from a Millipore water purification system (Milli-O IQ Element, MERCK, Germany). Fluorescent images were obtained on a ZEISSTM Axio Zoom.v 16 fluorescence stereoscope using the ZEN 3.5 (blue edition) software (Carl Zeiss, Germany). The absorbance and fluorescence spectrum data were recorded using an Agilent BioTek SYNERGY Mx® spectrofluorometer in a 96-weel microplate. The reagents and equipment employed in this experimental protocol, including their catalogue reference, are outlined in Supplementary Table S1 and Supplementary Table S2, respectively (Supplementary Data).
Gavage solution
Prepare a 5% RITC-dextran stock solution by dissolving the dextran powder in deionized water (ddH2O). Once the dextran is fully dissolved, aliquot the solution into 50 μL volumes and store at −20°C to minimize freeze–thaw cycles.
RIPA lysis buffer
For the extraction of cellular proteins, radio-immunoprecipitation assay (RIPA) buffer was prepared in accordance with the protocol described by Ngoka. 25 The final concentrations of the components in the RIPA buffer are as follows: 150 mM sodium chloride, 1.0% Igepal CA-630, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate, and 50 mM Tris-HCl at pH 8.0. Aliquot the RIPA solution into 10 mL volumes and store at −20°C to minimize freeze–thaw cycles. Supplementation with protease and phosphatase inhibitors should be added at the time of cellular lysis.
Experimental protocol
Kinetic data and identification of fluorescent signal in ex vivo tissue samples using cell image analysis systems
To better understand the dynamics of marker passage through the zebrafish intestinal tract, we conducted a pilot study prior to our main experiment using the Casper strain. Each animal was gavaged with FITC-Dextran 4,000 MW following standard protocols (Supplementary Fig. S1 and Supplementary Fig. S2, Supplementary Data), as described in the next section. The distribution of the fluorescent dye was evaluated in ex vivo samples from intestinal and extraintestinal sites, including the liver, spleen, heart, kidney, and gonads, at various time points (5, 30, 60, 120, 240, 360, and 480 min postgavage; Supplementary Fig. S2). For this, tissue samples were prepared and placed on a 24-well plate, ensuring uniform distribution to minimize signal variability. Imaging was conducted on the IVIS Spectrum system (PerkinElmer, Waltham, MA, USA), which was calibrated specifically for FITC fluorescence detection. System parameters were optimized with an excitation filter set to 465 nm and an emission filter at 520 nm. The exposure time was carefully selected to maintain a high signal-to-noise ratio without saturation, and the field of view was adjusted to cover the part of 24-well plate. Medium binning (2 × 2) was used to balance resolution with sensitivity. Images were acquired under consistent conditions across all samples to ensure reliable comparisons. For each well/tissue, background correction was applied to isolate the FITC fluorescence signal. Fluorescence intensity was quantified using Living Image software (PerkinElmer), with regions of interest defined for each sample. Background fluorescence was subtracted, and the mean fluorescence intensity (MFI) was calculated and then exported for further statistical analysis (Supplementary Fig. S2).
RITC-dextran (10,000 MW) permeability assay
Day 1: the day before gavage
Prior to the procedure, fish were submitted to 36 h fast to ensure an empty intestine. It is crucial to fast the fish for a minimum of 24 h to ensure an empty intestinal bulb. We observed better results with a 36-h fast, which minimized regurgitation and loss of the solution through the mouth and gills due to a partially full intestinal tract.
Thaw the 5% RITC-dextran stock solutions at room temperature. Subsequently, dilute the stock to prepare a working solution of 2% RITC-dextran in phosphate-buffered saline (PBS). Store it at 4°C.
Prepare a solution of 25% ethanol in ddH2O as be use as the positive control.
Day 2: on the day of gavage
Preparing the anesthetic solution:
Prepare a 150 mg/L solution of MS-222 from a 400 mg/L stock solution in system water. Set up postprocedure individual recovery tanks filled with system water that is free from MS-222.
Preparing for gavage:
Carve a groove into a sponge using a scalpel blade. Saturate the sponge in system water that is free from MS-222. Place the moistened sponge on a flat surface.
Preparing RIPA lysis buffer:
Thaw ∼10 mL of RIPA buffer solution on ice. Add one protease inhibitor tablet and one phosphatase inhibitor tablet to the solution. Mix until the tablets are completely dissolved. Aliquot 450 µL of this prepared solution into Precellys tubes containing 0.4 g of zirconium beads. Maintain the tubes on ice.
The procedure for oral gavage was adapted from Collymore and coworkers. 26 Individual fish (n = 15) were sedated in separate containers using 150 mg/L of MS-222 (tricaine methane sulfonate buffered solution) in system water until they lost their righting reflex and became unresponsive to a tail fin pinch, yet maintained opercular movement. This typically took about 2–3 min, as described by Matthews and Varga. 27 Once properly sedated, each fish was removed from the sedation container and positioned in a groove carved into a sponge that had been soaked in system water, with the head protruding slightly.
Oral gavage was performed using a micropipette fitted with a 10 µL tip. The tip was gently inserted ∼1 cm into the mouth, ensuring it passed beyond the orobranchial cavity and esophagus to reach the intestinal bulb. A volume of 3 µL of 2% RITC-Dextran solution (corresponding to 2 mg/100 mg body weight) was then carefully deposited into the anterior segment of the digestive tract, and the tip was gently withdrawn. Care was taken to ensure that the solution did not escape through the gills or mouth. Limit the gavage volume to more than 3 µL of any solution to prevent regurgitation and loss of the solution through the mouth and gills.
Following oral gavage, the fish were removed from the sponge and placed in a Petri dish for evaluation and imaging under a fluorescence stereoscope. Subsequently, place zebrafish in individual recovery tanks and document the time of gavage for each tank. The procedure was then repeated for each subsequent fish, with a recommended 20-min interval between each gavage (see Notes in Supplementary Section).
Fish were imaged under a fluorescence stereoscope using consistent exposure settings immediately (0 h), and at 2 h and 4 h postgavage (refer to Fig. 1 for details). Images were processed using Zen Blue 3.5 software to monitor the transit of the dye through the intestine. To investigate in vivo intestinal permeability, we firstly assessed the presence of the fluorescent marker in blood vessels. Four hours postadministration of the probe, fish were euthanized via immersion in buffered MS-222 at a concentration of 400 mg/mL.

Dextran assay to access intestinal permeability in adult zebrafish (Danio rerio). In this assay, fish were gavaged with a 2% RITC-Dextran solution with a molecular weight of 10,000 Da. Representative images illustrate the transit of the fluorescent marker through the intestine for the negative control (vehicle;
For negative controls, a separate group of fish (n = 7) was gavaged with an equivalent volume of PBS. As a positive control, 15 fish were gavaged with 3 µL of 25% ethanol 3 h prior to the oral administration of the fluorescent marker. Ethanol exposure is known to compromise epithelial barrier integrity.28–30
Gut solute permeability evaluation
Due to the challenges associated with obtaining sufficient blood volume for assessing labeled-dextran absorption in individual fish, we developed a protocol to evaluate the distribution of the dye in extra-intestinal tissues to assess intestinal permeability in adult zebrafish. Four hours post-RITC-dextran gavage, fish were euthanized via immersion in buffered MS-222 at a concentration of 400 mg/mL, and tissues were harvested. As described in “kinetic data and identification of fluorescent signal in ex vivo tissue samples using cell image analysis systems” section, we examined the distribution of the fluorescent dye in extraintestinal sites such as the liver, spleen, heart, kidney, and muscle in ex vivo samples using fluorescence stereoscopy and cell image analysis systems (Supplementary Fig. S2, Supplementary Section). Our findings indicate that the liver was the main extraintestinal site displaying the fluorescent signal. As a result, we chose to quantify fluorescence intensity in the liver as an indicator of intestinal permeability.
Gut and liver samples were carefully dissected under stereomicroscope (Zeiss Stemi 305—refer to Supplementary Table S2) and placed in 0.1 M phosphate saline buffer to remove any adhered non-lever tissue or substances, including the fluorescent probe, fat, and spleen, with fine forceps. Immediately following dissection, both tissues were imaged under a fluorescence stereoscope using consistent exposure settings (Supplementary Fig. S3, Supplementary Section). Then, the tissues were placed in Precellys tubes containing 0.4 g of zirconium oxide beads and 450 µL of cold RIPA buffer supplemented with protease and phosphatase inhibitors. Tissues were then homogenized using a Precellys® 24 Homogenizer system (refer to Supplementary Table S2) in a cycle of 2 × 12,000×g for 20 s. The homogenates were centrifuged at 12,000×g for 15 min at 4°C to remove insoluble debris, and the supernatant was stored at −80°C. To normalize for variations in organ size, we standardized the concentration of total protein to 100 µg per sample. Total protein concentration was determined using a bicinchoninic acid assay, following the manufacturer’s protocol (refer to Supplementary Table S1). Recovery of rhodamine B probe was measured using excitation wavelengths of 550 nm and emission wavelengths of 590 nm, using a Synergy H1 multimode plate reader (BioTek). Livers and guts from fish gavaged solely with PBS were processed identically to those in the RITC-dextran groups and served as negative controls. The blank consisted solely of RIPA buffer without any sample. Values were calculated by subtracting the mean of the blank readings. All analyses were performed in triplicate. Both the total protein concentration and the fluorescence signal in the liver were measured on the same day for all samples to standardize the procedure.
Confocal microscopy imaging
We performed ex vivo confocal microscopy imaging technique to identify the ability of the zebrafish intestinal epithelium to take up fluorescent dextran (MW ∼ 10 kDa). Four hours after the RITC-dextran gavage, fish were euthanized (immersion in buffered MS-222 at 400 mg.L−1), and the intact intestine (n = 3 per group) was collected, washed with PBS 1×, and stored at −20°C. Freeze-fresh fragments from anterior (Supplementary Fig. S4), middle (Supplementary Fig. S5), and posterior intestine were sectioned in ∼2 mm and mounted in slides and stained with Hoechst 33342 (Thermo Fisher Scientific, USA). Tissue sections were examined using the inverted laser-scanning confocal microscope Zeiss LSM 780-NLO (Carl Zeiss, Germany) at 405–453 nm to detect nucleus and at 543–613 nm to detect the fluorescent red marker (Supplementary Fig. S4 and Supplementary Fig. S5). Images were acquired using a 63×/1.4 NA Oil objective and 3× zoom in 1024 × 1024-pixel images.
Statistics
Statistical analyses were conducted using GraphPad Prism 9.0. The distributions of variables were assessed using the Shapiro-Wilk test. Data are expressed as mean ± standard error of the mean (mean ± SEM). The study groups were compared using Kruskal-Wallis multiple comparison test. Statistical significance was set at p < 0.05.
Results and Discussion
In the present study, we describe a detailed protocol for evaluating gut solute paracellular permeability in adult zebrafish. This is achieved through the detection and quantification of 10 kDa RITC in extraintestinal sites as an indicator of in vivo intestinal permeability. In addition, the results show that intestinal paracellular transport of macromolecules is significantly higher in the pretreated alcohol group, reinforcing its prejudicial acute effects on the integrity of the gut barrier.
Intestinal permeability involves three pathways: the pore, leak, and unrestricted pathways. Pore and leak pathways require TJ flux, while the unrestricted pathway is TJ independent. Concisely, the pore pathway is a high-capacity route regulated by pore-forming claudin proteins that are selective based on charge and size allowing solutes with a maximum diameter of 0.6 nm to pass through. In contrast, the leak pathway is a low-capacity route, is not charge-selective, and permits the passage of molecules 20 times larger (∼ 12.5 nm) than those transported by the pore pathway. 31 In our experimental assay, we proposed to evaluate intestinal permeability to a large solute due our interest to investigate whether the gut of adult zebrafish is permeable to a large solute under physiological and/or impaired conditions. Given that molecules with a molecular weight of 10,000 are too large to be absorbed through the pore pathway (10kD dextran = ∼23Å),32,33 they should remain within the intestinal lumen if the barrier integrity is not compromised. Although 10 kDa dextran could potentially be transported via the leak pathway, this route is likely to exhibit slower kinetics. 32
To our knowledge, no prior studies have evaluated intestinal permeability in vivo in adult zebrafish or any similar fish model that lacks a stomach. To better understand the dynamics of marker passage through the zebrafish intestinal tract, we performed a pilot study in the Casper strain (see Supplementary Fig. S1 and Supplementary Fig. S2). Based on prior research involving zebrafish larvae 17 and mice,11,34 we tested different time points to visualize the transit of the fluorescent marker through the intestine. During this pilot, we observed that the fluorescent probe took more than 8 h to reach the posterior intestine, and liver uptake of FITC 4 kDa was lower at 6–8 h compared with 2–4 h postgavage (Supplementary Fig. S2), as observed through fluorescence stereomicroscopy. According with these findings, we concluded that the absorption of 4 kDa dextran occurred between 2 and 4 h, and we determined that a 4-h time frame was a reasonable time for the macromolecule to be transported through the leak pathway. In addition, the midgut appears to be the primary region for uptake of large molecular weight molecules in this model (Supplementary Fig. S2 and Supplementary Fig. S5). These results align with previous studies conducted on zebrafish larvae.13–15
In the initial trial with Casper, we utilized FITC-dextran due to its widespread use and compatibility with our imaging setup. However, during the experiments, we observed a significant background signal when using the green fluorescence detection filter (Supplementary Fig. S1). To address this issue, we opted to switch to RITC-dextran, which emits in the red fluorescence spectrum and exhibited substantially lower background interference in our imaging system. In the present study, no significant differences were detected in the distribution of 10,000 MW RITC-dextran between the 20- and 60-min marks, or between the 60- and 120-min marks, in different intestinal regions among the animals in vivo. To minimize repeated anesthetic exposure, which could be lethal and also reduce intestinal motility, 35 we opted for evaluations immediately postgavage (0 h) and at 2- and 4-h post-administration of the intestinal permeability marker. Immediately postgavage, the fluorescent probe was localized in the anterior segment of the intestine, with a minor fraction reaching the mid-intestine (Fig. 1A–C). Up to 2 h postgavage, RITC-dextran with a molecular weight of 10 kDa remained largely localized to the intestinal lumen (mid-intestine) in all animals (Fig. 1D–F). At 4 h postgavage, a substantial amount of labeled dextran remained in the mid-intestine of all RITC-dextran gavaged fish (Fig. 1G–I), while a smaller quantity of the marker was observed in the hindgut of some test animals (Fig. 1I).
Cocchiaro et al. 17 showed that when 10-kDa Texas-Red was orally administered to zebrafish larvae, it was largely confined within the intestinal lumen, similar to our observations (Fig. 1). However, when co-gavaged with 20 mM of ethylenediaminetetraacetic acid (EDTA), a known disruptor of epithelial TJ, 36 the fluorescent dextran rapidly translocated to the basolateral side of the epithelium and dispersed to other tissues. We attempted to replicate Cocchiaro et al.’s 17 approach by gavaging adult fish with 20 mM of EDTA to induce TJ damage. Unfortunately, all the test animals perished. Most fish lost consciousness within approximately 1 h postgavage, and some exhibited opercular bleeding. Even at lower concentrations, EDTA proved lethal. Known for its chelating properties, EDTA reversibly opens TJs by depleting Ca2+ and Mg2+, which maintains the structure and conformation of TJ proteins. 36 The lethal effects of EDTA on zebrafish warrant further investigation. Conversely, concentrations of ethanol ranging from 25% to 50% have been used as a vehicle for inducing chemical enterocolitis in adult zebrafish28,37 and can lead to gut inflammation in a reversible manner. 28 In our study, we used acute ethanol exposure as a positive control to induce epithelial barrier disruption28,30 and thereby increase intestinal permeability to macromolecule, as demonstrated in Figures 2 and 3.

Dextran is released into bloodstream after gavage in adult zebrafish (Danio rerio). In this assay, fish were gavaged with a 2% RITC-Dextran solution with a molecular weight of 10,000 Da (n = 15). Representative images demonstrate an increase in fluorescence intensity within the vascular lumen (black arrows) of the caudal fin in the positive control group (RITC-dex + ethanol;
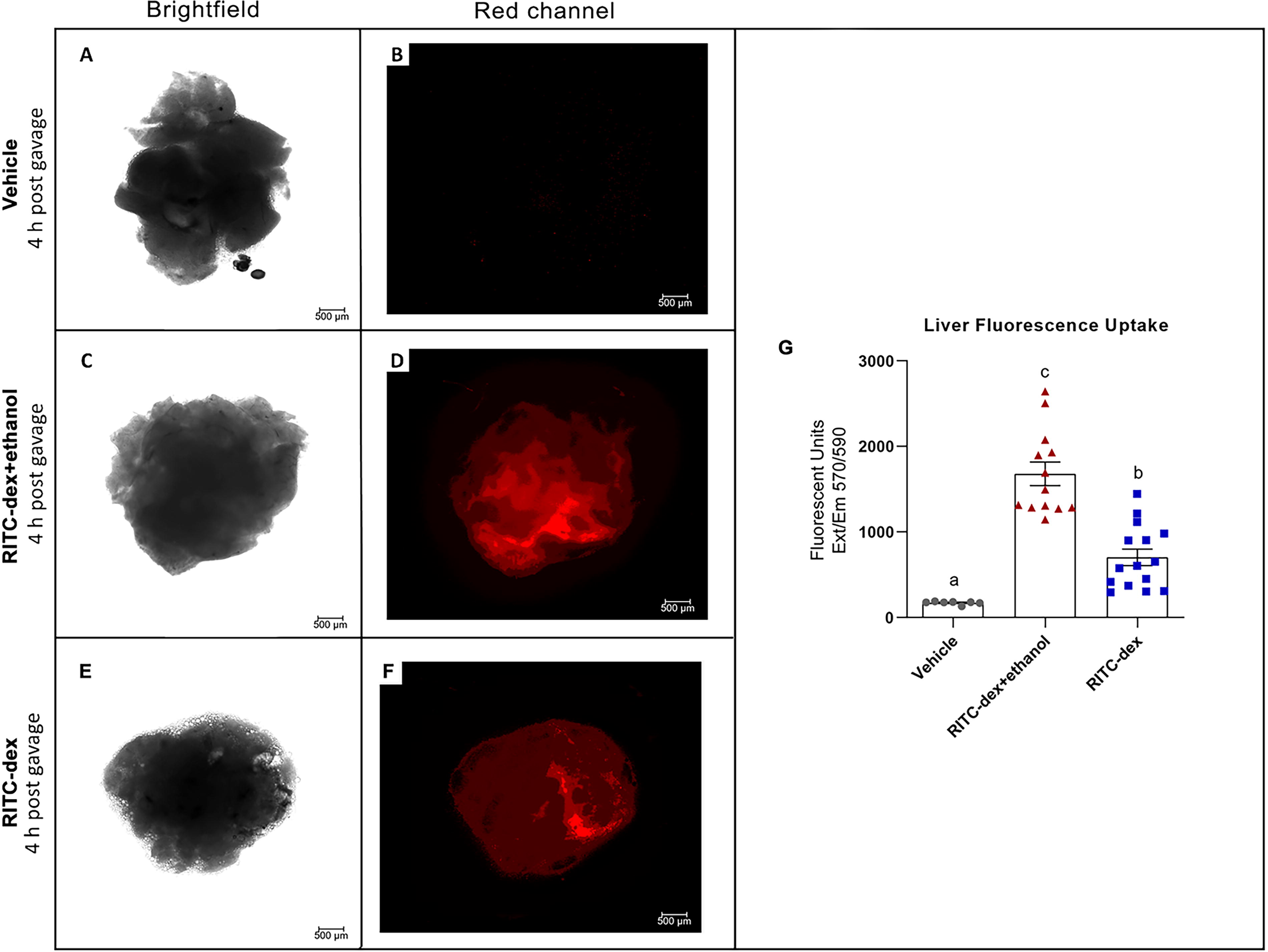
Dextran assay for epithelial barrier integrity in adult zebrafish (Danio rerio). Ex vivo fresh liver samples were obtained 4 h postgavage with a 2% RITC-Dextran (10,000 MW) solution from the negative control (Vehicles,
We also detected the presence of the fluorescent marker within the vascular lumen of the zebrafish caudal fin at 2 h postgavage (Fig. 2A–I). Similar results were observed in adult mice, where blood serum levels of 10-kDa FITC-dextran peaked 2 h after oral administration. 34 This suggests that the kinetics of intestinal transport for a 10,000 MW labeled-dextran may be comparable between zebrafish and mice. A stronger and higher fluorescence signal intensity was identified within the vascular lumen in the positive control group (Fig. 2D–F) related to the test group (Fig. 2G–I). However, the fluorescence signal within the vessels was detected in only a few animals that received the dye. Due to this variability, we decide to complement this approach with the quantification of fluorescence labeled marker in other extraintestinal sites. Image data revealed an increase in the fluorescent signal corresponding to liver uptake of labeled dextran in both the test and positive control fish (Fig. 3D and F). In contrast, no fluorescent signal was observed in the livers of fish gavaged solely with vehicle (Fig. 3B). As previously mentioned, we quantified the total protein concentration to account for variations in organ size among the fish. As expected, the normalized MFI in the liver of the positive control group was significantly higher compared to that of the test group (Fig. 3G). Consistently, oral pre-exposure to 25% ethanol effectively enhances intestinal paracellular permeability to a large solute in adult zebrafish suggesting a decrease in the tissue integrity of the intestinal wall.
An important aspect of the present methodology is the observation of resistance to the passage of the gavage tip in some fish, even under confirmed sedation. In these cases, attempts to deepen the gavage resulted in perforation of the intestinal tract and widespread intraperitoneal diffusion of fluorescence. As illustrated in Figure 1B and 1C, a portion of the fluorescence was observed overlapping the gills immediately after gavage, with the signal absent at subsequent evaluated time points. The gills, as the primary organ responsible for ionic and acid–base homeostasis in adult zebrafish, possess a high density of ionocytes that facilitate ion transport across the epithelium. 38 This tissue is a key focus of studies on aquatic toxicity, and despite its importance in absorption contexts, the literature lacks specific data on the size of molecules capable of diffusing into circulation through the gills. Osborne et al. (2015) 39 demonstrated that 100 nm silver nanoparticles (∼1,000 MW) were not deposited in the gills of adult zebrafish and exhibited lower toxicity, potentially due to limited bioavailability. 38 Thus, we hypothesize that while limited diffusion of the high molecular weight marker through the gills may occur in some animals, this process is proportionally less significant compared to intestinal permeability. This reinforces the importance of methodological precision to avoid confounding factors such as unwanted marker distribution or tissue damage during gavage procedures. Leal et al. 22 described the use of the Ussing chamber for functional analysis of the intestine (at the epithelial level) in adult zebrafish. Although this technique provides a robust experimental framework, some are challenging such as the size of the zebrafish intestine, as well as tissue viability throughout the experiment, must be taken into consideration for its execution. Furthermore, there is a delicate balance between paracellular permeability and barrier function. 31 Based on extensive literature, it seems reasonable that the microbiota plays an important role in regulating the integrity and functionality of the gut barrier, maintaining homeostasis of the whole organism. 40 Thus, extraction of host tissue may not represent the complexity of the process that occurs in vivo, including host–microbe interactions. 4 Recent researches have shown sex-related differences in gut microbiota composition particularly in relation to microbial community aggregation strategies in the face of external environmental pressures.41–43 Interestingly, mammalian studies indicate that estrogen has beneficial effects on intestinal barrier function. It promotes bicarbonate secretion into the gut and enhances the expression of TJ-proteins, especially occludin. 44 To mitigate potential gender-related variables, only female fish were used in our first investigation. Future experiments may be conducted to determine if this model system is applicable to both sexes and serve as a reference guide for expand studies on intestinal permeability induced by genetic, pathophysiological, or pharmacological events.
Conclusion
In summary, we introduce a protocol for assessing in vivo intestinal permeability in adult zebrafish based on the extraintestinal diffusion of high-molecular weight dextran. Based on our findings, we conclude that measuring the fluorescence intensity in extraintestinal tissues is a more reliable method than measuring the fluorescence in the bloodstream in vivo for adult zebrafish. Key advantages of this protocol include the ability to monitor the passage of the fluorescent marker through the intestine in real time and its applicability for detecting the presence of the dye in extraintestinal sites.
We propose the use of zebrafish as a model for studying intestinal permeability in vivo due to its potential versatility to complement methodologies that are challenging to implement in rodents, such as live imaging and high-throughput screening.17,18,21 It is worth noting that the increasing availability of genetically engineered zebrafish lines enables the study of specific pathological or genetic conditions. 19 Therefore, the development of reliable and sensitive methods for evaluating intestinal permeability is crucial for advancing both basic and translational research.
Limitations
This study was designed to evaluate intestinal permeability in a manner accessible to all zebrafish strains, including those with pigmented backgrounds. However, pigmentation in the AB strain posed a significant challenge, restricting our ability to observe the fluorescent marker to tail-specific circulation. This limitation underscores the difficulty in visualizing fluorescent signals in pigmented fish, particularly in nonperipheral regions. Furthermore, detecting small quantities of the marker in the bloodstream in vivo would ideally require high-resolution imaging systems, such as confocal microscopy. Unfortunately, we were unable to access any facilities with high-resolution imaging technology suitable for use with adult zebrafish. In addition, the use of flexible tubing or pipettes could help mitigate the risk of intestinal tract perforation during gavage procedures and would enable deeper insertion of the marker directly into the intestine, reducing its retention in the gills.
Footnotes
Acknowledgments
The authors thank Ricardo Soares Oliveira for the student financial support (PROPESP-FURG/Brazil). Dr Mario Costa Cruz helped in the process of standardization of permeability intestinal assay and image assistance. Meire Hiyane and Marlise Bonetti Montes for laboratory technical assistance. Caroline Castro for figures assistance.
Authors’ Contributions
M.G.M.: conceptualization; methodology; formal analysis; validation; investigation; writing—original draft; and writing—review and editing. C.I.M.F.: conceptualization; methodology; formal analysis; and writing—review and editing. B.F.N.: conceptualization; and writing—review and editing. N.O.S.C.: supervision; methodology; funding acquisition; and writing—review and editing. L.F.M.: supervision; project administration; conceptualization; methodology; funding acquisition; and writing—review and editing.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (Grant numbers 2017/05264-7 and 2015/21644-9). M.M. is a doctoral student financed by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Finance Code 001). LFM is a research fellow from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Proc. 309634/2018-0).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.