
Abstract
The zebrafish (Danio rerio) has become an essential model in fields such as developmental biology, toxicology, genetics, and regenerative medicine due to its low cost, small size, transparent embryos, and genetic similarity to humans. Nowadays, this model is increasingly recognized as a valuable tool in other fields, including veterinary medicine and animal production research, particularly aquaculture, due to its unique characteristics that make it suitable for studying economically significant diseases affecting production species. However, unlike established models such as mice, zebrafish lack standardized protocols for housing, feeding, anesthesia, and sample collection, which affects study reproducibility. In addition, it is a common practice to use whole zebrafish larvae or juveniles for metagenomic studies rather than analyzing individual organs, despite the fact that gene expression can vary between organs. This approach complicates the attribution of findings to specific biological processes. To address this, various authors proposed protocols for sample collection in larvae, juveniles, and adult zebrafish; however, comprehensive studies encompassing nearly all fish organs are scarce. Understanding zebrafish anatomy and the technical requirements of the study is essential for accurate sample collection. Some challenges present during zebrafish dissection include the small size of the fish, the fragility of their organs, and the faster onset of autolysis and heterolysis after the death of the animal. Using magnifying lenses, microdissection tools, and conducting dissections on cold surfaces can help mitigate these issues. This article aims to improve sample collection for histopathological and genetic studies in adult zebrafish by establishing a comprehensive, organized, and systematic dissection protocol that accounts for the anatomical specifics of this experimental model.
Introduction
The zebrafish (Danio rerio) has emerged as an invaluable model organism in scientific research, playing a pivotal role in a wide variety of disciplines such as developmental biology, toxicology, genetics, cell line research, and regenerative medicine. 1 , 2 Some of the characteristics that make this model appealing include its low cost, its small size, the transparency of its embryos, and its genetic homology with humans (70%), sharing more than 84% of the genes involved in disease, highlighting its relevance in biomedical research.3–5 This model is also increasingly recognized as a valuable tool in veterinary medicine and animal production research, particularly aquaculture. Its unique characteristics make it suitable for studying economically significant diseases affecting production species caused by pathogens such as Mycobacterium marinum, 6 Vibrio anguillarum, 7 viral hemorrhagic septicemia virus (VHSV), 8 and Ichthyophthirius multifiliis. 9 In addition, zebrafish is being utilized in areas such as nutrition research, facilitating the testing of a large number of novel diets and the evaluation of the incorporation of nutraceutical molecules into feed. 10 The results obtained from these studies are often transferable to commercially relevant aquaculture species, enhancing their applicability to production systems. 11
Unlike more established models, such as the mouse, which benefit from highly standardized housing and sampling protocols due to their long history of use, the zebrafish, having been utilized as an animal model for a shorter period, still requires the development of such standardized practices. Although different texts specialized in this model provide useful guidelines for the laboratory husbandry, care, and breeding of zebrafish, 12 , 13 the differences in animal maintenance, feeding, anesthesia, or euthanasia protocols, as well as sample collection across zebrafish research centers or experimental studies, affect the reproducibility and reliability of results. 14 , 15 Regarding sampling, it is still a common practice to collect the whole zebrafish larvae for many types of studies. 16 , 17 This practice results in greater difficulty when interpreting the data obtained from molecular studies such as gene expression analysis or microbiome profiling. Different studies demonstrate the differential expression of the same gene in different organs of the same zebrafish. 16 This phenomenon, known as tissue-specific expression, is crucial for the proper functioning and specialization of various biological systems. 18 Therefore, by using whole specimens in molecular studies, the results obtained will reflect a mixture of markers from various tissues, making it difficult to accurately attribute findings to particular organs or biological processes, which can obscure the interpretation of molecular patterns and hinder precise conclusions. Also, in the majority of histological studies on adult zebrafish, fixation and processing are performed using whole specimens. Different sampling protocols for sample collection have been reported in larvae, juveniles, and adult zebrafish.19–24 However, there is a notable lack of studies providing a standardized and comprehensive protocol for the individual sampling of nearly all zebrafish organs. 14
In the case of zebrafish dissection, several challenges may arise due to their small size (approximately 3 cm in adult specimens), variability among individuals, the fragility of their organs (which increases the risk of tissue damage during dissection), and researchers limited hands-on skills on this model. Everything mentioned above can contribute to sampling errors that affect the accuracy of subsequent analyses. 25
Knowledge of the species’ anatomy and physiology, as well as the technical requirements of the studies to be developed, is of great importance when planning sample collection. Although zebrafish share a similar anatomy with other fish species of commercial or research interest, there are notable differences that are noteworthy. For instance, zebrafish lack of glandular stomach, which is replaced instead by the intestinal bulb, 26 and present pharyngeal teeth, which develop from the 5th ceratobranchial toward the ventral surface of the pharynx. These teeth meet the mucosal pad, which is composed of a stratified keratinized epithelium that protects the dorsal pharyngeal mucosa in the chewing region and allows for grinding of food. 27 , 28 In a similar fashion, as for other fish species, the faster onset of autolysis and heterolysis when compared to mammalian models is an important factor to consider for histological and molecular studies due to the rapid degradation of tissue. 29
Another critical factor that may compromise the interpretation of results is the presence of clinical or subclinical infections, as well as parasitic infestations, which are commonly observed in most research facilities working with zebrafish. Proper sample collection and subsequent histopathological evaluation are essential to assess the health status of the animals, a prerequisite for ensuring the suitability of control specimens used in studies. 30
This article aims to optimize a complete sample collection for histopathological and molecular studies in adult zebrafish by establishing a comprehensive, organized, and systematic dissection protocol, considering the sampling and anatomical specificities of this experimental model.
Materials and Methods
Fish
The zebrafish used in this protocol were supplied by a research institution officially authorized for the production and distribution of this experimental model. All procedures have been reviewed and approved by the University of Santiago de Compostela Research Ethics Committee and adhere to the guidelines established by the committee for human/animal research. Experimental conditions have been designed to minimize pain and suffering, and all possible measures have been taken to ensure animal welfare. A total of 235 zebrafish were dissected following the protocol described below to obtain samples from individual organs, aimed to perform DNA/RNA sequencing analysis, as well as histopathological evaluations, in the context of an H2020 project. In addition, 20 specimens were used to compare the protocol described below with other fixation and sample collection techniques in this species.
Materials
Ethyl 3-aminobenzoate methanesulfonate (Tricaine Pharmaq 1000 mg/g).
Sodium bicarbonate (NaHCO3).
EtOH 70%.
Petri dish, 140 × 20 mm.
Magnifier Lens, w/90 Leds, 5 diopter.
Entomological needles number 000.
Micro-Adson Forceps.
Micro scissors.
RNAlater (Thermo Fisher Scientific, AM7021).
Bouin’s fluid (Picric Acid: PanReac AppliChem, 0002392475; Acetic acid glacial: Carlo Erba, P2H014082H).
Dissection protocol
Obtaining a cold work surface
Cut a sponge about 1.5 cm thick in the shape of a semicircle and place one of the halves on a Petri dish (14 cm diameter) (Fig. 1). Before use, disinfect the sponge with 70% ethanol, soak it in water, and freeze it. This will create a cold, firm, yet flexible surface to position the fish and help lower its temperature during dissection. Subsequently, place the Petri dish on a cold base to avoid rapid thawing of the sponge and cooling of the empty half of the Petri dish.
Euthanasia
Prepare a tricaine methanesulfonate solution at euthanasia concentration (0.8%), adding bicarbonate in a 1:2 ratio to avoid pH variations. Euthanize the zebrafish by anesthetic overdose by immersion in the tricaine solution. Keep fish in the solution for at least several minutes after the loss of reflexes to ensure euthanasia. After euthanasia, place the animal in a right lateral recumbent position (left lateral recumbent position if left-handed) in the center of the straight edge of the sponge; subsequently, perform the external inspection of the animal (Fig. 1 and Fig. 2A). Insert one blade of the scissors into the depression just caudal to the skull and dorsal to the caudal edge of the operculum. Close the scissors, severing the spinal cord (Fig. 2B, C). Confirming the euthanasia by spinal cord section ensures the absence of any type of suffering during the dissecting process.
Opening of opercular, pericardial, and abdominal cavities
Fix the fish to the sponge using two entomological needles. Insert one needle in the middle portion of the caudal peduncle, next to the caudal fin. Place the other needle in the anterior portion of the eye socket (displacing the eyeball so as not to cause damage to the organ), exerting some traction cranially to keep the fish stretched. Place a magnifying glass (4X or higher) over the fish before starting the dissection. Gills are located in the opercular cavity (covered by the operculum) and conformed by four gill arches on each side. Each arch is composed of gill rakers, oriented to the oropharynx, and two caudally orientated hemibranchs, conformed by filaments and lamellae. After cutting the operculum (Fig. 3A), the gill arches could be dissected by cutting their dorsal and ventral insertions with scissors, trying not to cause damage to the delicate gill filaments. Due to the rapid development of autolysis in this organ, it is recommended to dissect and fix them as soon as possible after the death of the animal to avoid deterioration. To expose the internal organs, including the heart, open a lateral flap in the abdominal wall by making four incisions. The first incision will extend from the anus, along the midline, to the ventro-rostral insertion of the operculum. The second incision begins at the anus and ends dorsally below the lateral line. From the endpoint of the second incision, make a third incision ventrally and along the lateral line, finishing at the caudal edge of the operculum. Finally, the first and last cuts will be joined, bordering the gills (Fig. 3A). Whether microbiological samples are intended, the first incision could be made in the opposite direction to avoid contamination of the abdominal cavity with intestinal content.
Intestine dissection
Once the abdominal cavity is exposed, observe the disposition of the internal organs (Fig. 3B, C, D and E) and the two curvatures present in the digestive tract. The first curvature is located ventrally in the middle of the abdominal cavity, encircling the liver. Starting from this curvature, the midgut advances cranially toward the anterior third of the cavity, where the second curvature starts. This second curvature is positioned above the rest of the abdominal organs, adjacent to the left border of the liver. The intestine then continues caudally, following a straight trajectory towards the posterior intestine (Fig. 4A). To dissect the intestine, gently pull the second curvature in a ventral direction with forceps, removing its peritoneal connections (Fig. 4A). This will allow the observation of the full length of the intestine, maintaining its connections with the esophagus and the anus. Use two forceps to separate the peritoneum attachments between the intestinal segments (Fig. 4B), obtaining a “U” shape (Fig. 4C). Using scissors, carefully sever the connection between the esophagus and intestinal bulb, ensuring that the rest of the digestive tract remains connected to the anus, in order to facilitate the recognition of the different parts of the intestine (intestinal bulb, mid and posterior intestines) (Fig. 4C). Subsequently, cut the connection of the posterior intestine with the anus and extend the entire dissected digestive tract in the half of the Petri dish devoid of sponge.
Dissection of other abdominal organs.
The liver and the spleen will remain connected to the intestinal bulb and mid intestine (Fig. 3B and Fig. 4C), respectively, simplifying their dissection in the sponge-free section of the Petri dish. To extract the spleen, grasp the gastrosplenic ligament (that links the digestive tract and the spleen) and gently pull it using forceps, liberating the organ. For liver dissection, encircle the intestine with forceps near the liver terminus, then apply gentle pressure to slide them forward while stabilizing the posterior part of the intestine with another pair of forceps. This step can also be performed before the section of the esophagus. The ovaries are organs that may occupy a substantial portion of the abdominal cavity in adult specimens (Fig. 3B and C). To dissect the entire ovary, grip the central part of the organ with forceps and, after applying soft traction ventrally to isolate it from surrounding organs, sever the mesovarium dorsally and the oviduct, located in the caudal-most part of the abdominal cavity. Repeat this process for the other ovary. In males, testicles are located in the same position but have a smaller volume. The dissection method for the testes mirrors the one mentioned previously for the ovaries. The swim bladder is compounded by two chambers and is located in the dorsal-most portion of the abdominal cavity, just beneath the retroperitoneal kidney (Fig. 3B). By grasping the cranial chamber with the forceps, it can be easily removed by dragging it in a ventro-caudal direction.
Dissection of the heart
The heart is located ventro-caudal to the gills, within the pericardiac cavity, separated from the abdominal cavity by the transverse septum. To extract the heart, it is necessary to section the junctions of the bulbus arteriosus with the ventral aorta and the venous sinus with the hepatic and common cardinal vein.
31
Taking the ventricle (the only macroscopically visible part of the heart) as a reference, make two incisions with scissors cranially and caudally (Fig. 5). Subsequently, using forceps, remove the entire organ, exerting gentle pressure to avoid tissue rupture. Alternative protocols advocate for pre-dissection fixation to enhance the preservation of heart shape and structure.
23
Dissection of the kidney
The complete resection of the zebrafish kidney presents significant challenges due to its small size, anatomical complexity (divided into head and posterior kidney) (Fig. 3B and Fig. 6A), its friable consistency, and its retroperitoneal position in the dorsalmost part of the abdominal cavity. For a more accessible approach to the kidney, all the abdominal organs should be removed. Grab the remaining left part of the abdominal wall with forceps for better support during the process. To remove the organ, use a scalpel blade. First, place the blade on the left edge of the kidney and, applying pressure on the dorsal surface of the cavity, slide the blade toward the right edge of the organ (Fig. 6B). Once extracted, it can be easily removed with forceps. While this method allows the rapid removal of a substantial portion of the kidney, it may compromise its structural integrity. Alternative protocols are specialized in kidney dissection.
21
Dissection of the brain
To access the brain, place the animal in a prone position, securing it with forceps by applying lateral pressure. With the animal properly positioned, use scissors to make two incisions along the sides of the skull, starting from the euthanasia confirmation cut (located at the projection of the posterior edge of the operculum) to the dorsal edge of the ipsilateral eye socket. Then proceed to lift the dorsal part of the skull by lifting from caudal to cranial (Fig. 7A, B). Then, sever the optic nerves located in the anterior portion of the organ, which maintain the connection to the eyes by means of the optical chiasm. Finally, insert closed curved scissors beneath the brain to liberate it.
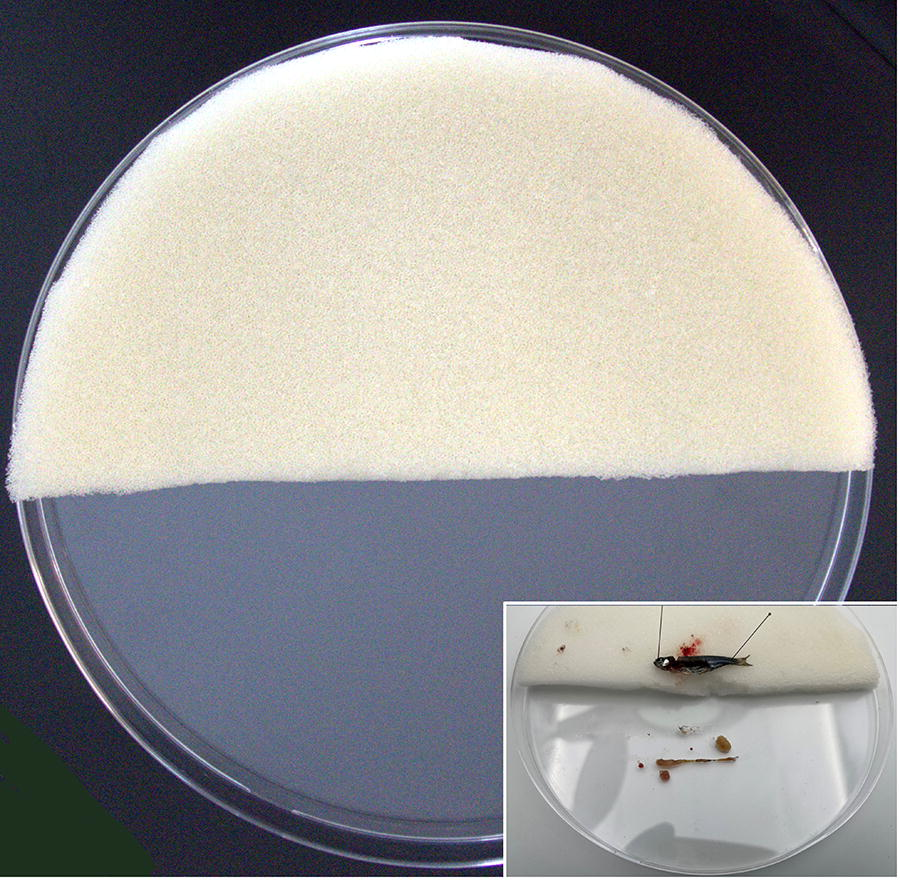
Platform on which the dissection will be performed, consisting of a Petri dish containing a semilunar sponge approximately 1.5 cm in thickness, which has been previously disinfected and frozen. Inset: Position of zebrafish during dissection.
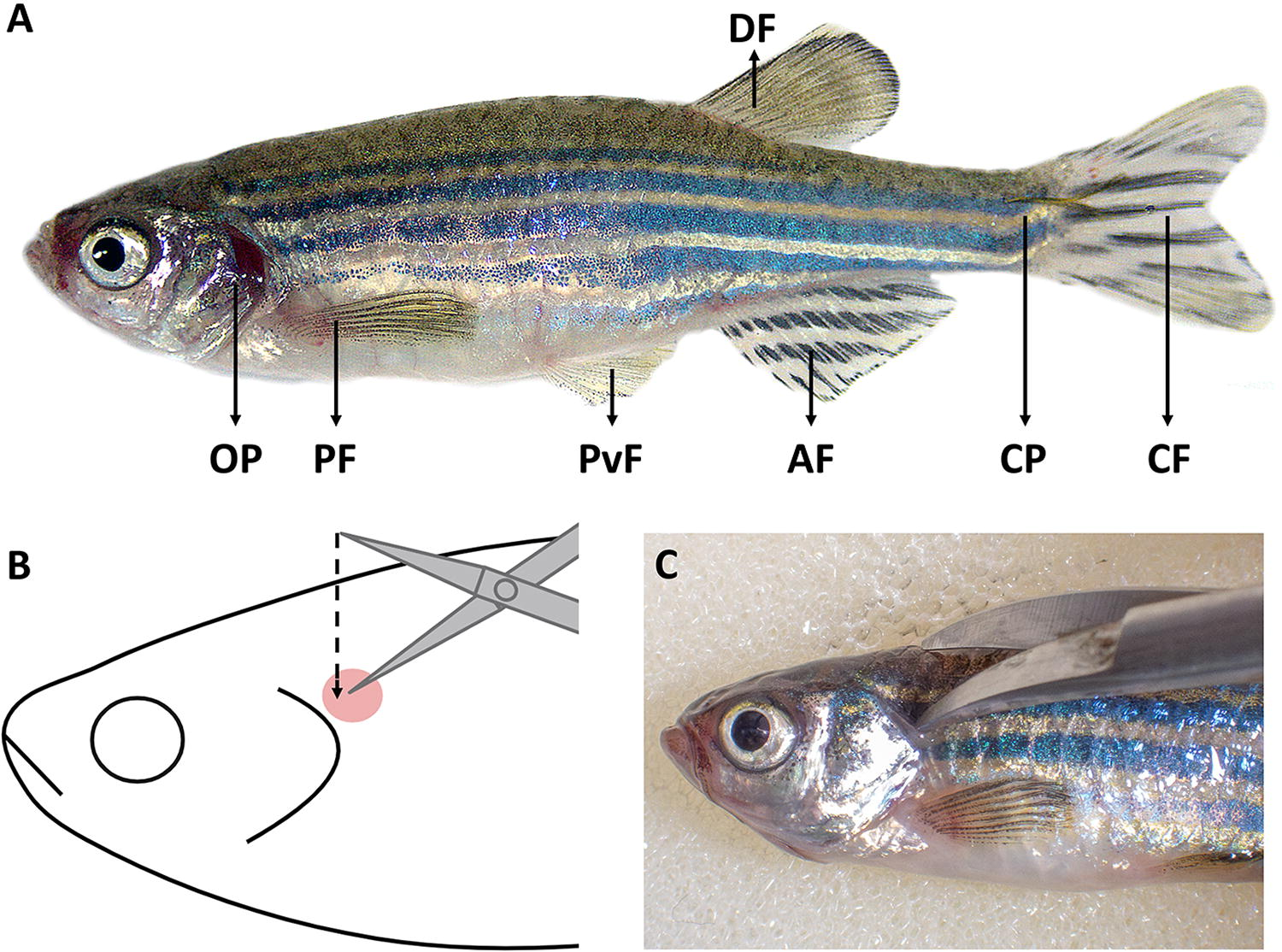
Zebrafish external anatomy and confirmation of death.

Opening of the opercular, pericardial, and abdominal cavities.
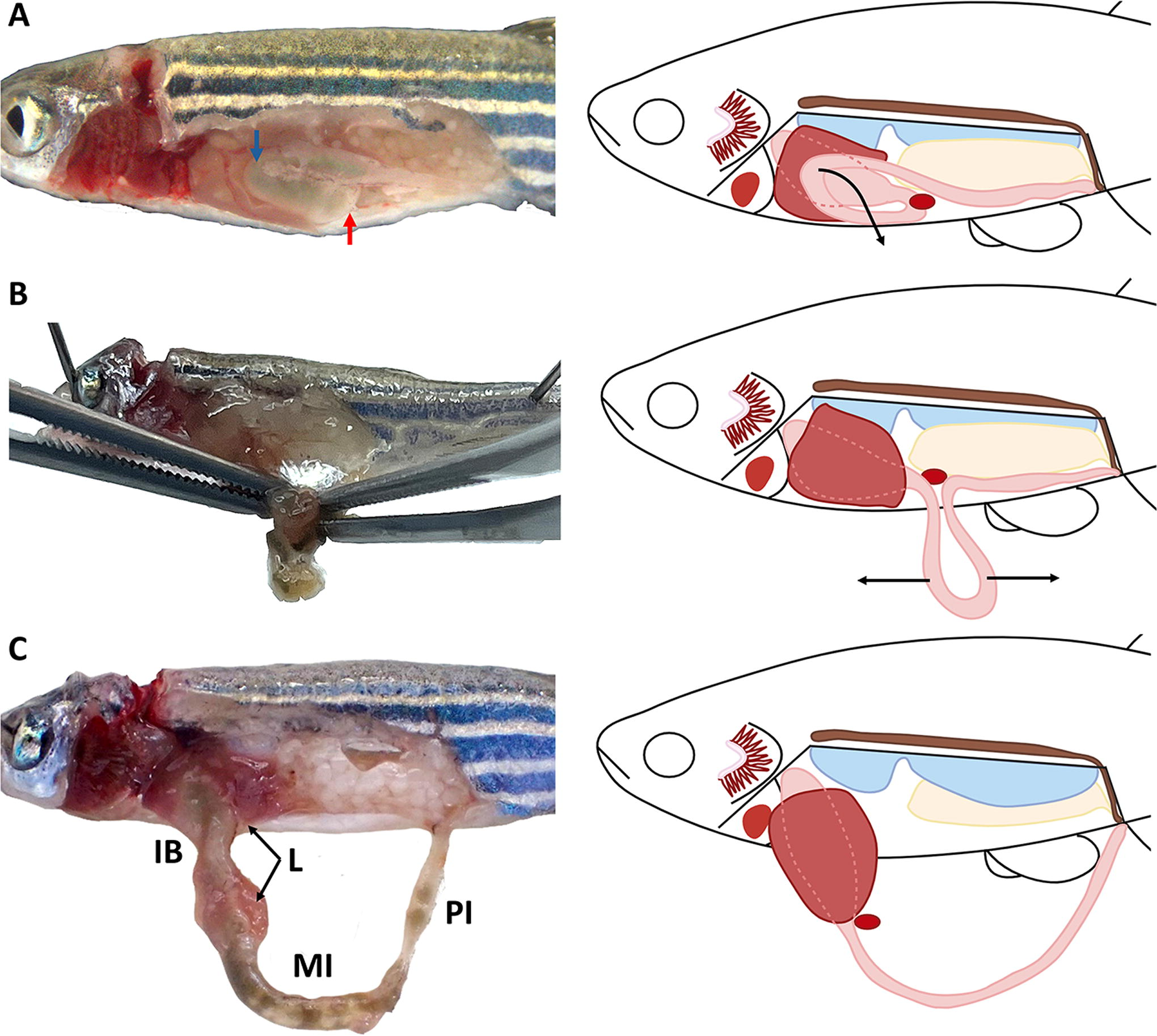
Photographic and schematic illustration of the intestine dissection in adult zebrafish.
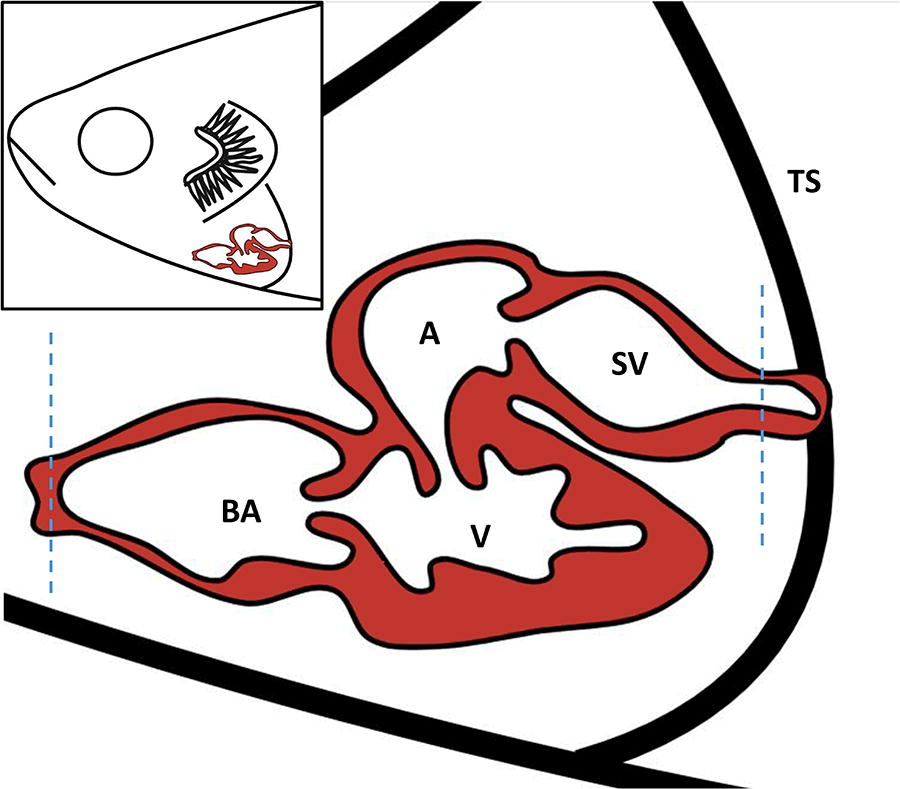
Scheme of heart resection in adult zebrafish through two incisions (blue dotted line) caudal to the venous sinus and cranial to the bulbus arteriosus. Bulbus arteriosus (BA), ventricle (V), atrium (A), sinus venosus (SV), and transverse septum (TS). Inset: location of the heart in adult zebrafish.

Kidney dissection in adult zebrafish.

Brain resection in adult zebrafish.
Comparison among different sampling procedures
Twenty additional zebrafish were used to evaluate the effects of individualized organ dissection, the use of cold plates, and elapsed sampling time on DNA/RNA integrity and preservation of the tissue architecture. To this end, eight of these animals were fixed whole in Bouin’s fluid following opercular resection and the creation of a lateral flap (an incision in the left abdominal wall) to enhance fixative penetration. This approach was aimed to evaluate the intestinal architecture obtained through this processing method in comparison to that obtained using the previously established protocol.
The remaining fish were used to evaluate the effects of using cold plates during dissection, as well as the impact of elapsed sampling time on RNA and DNA integrity and the development of tissue alterations in various organs of zebrafish samples. To this end, six fish were dissected on a frozen sponge and cold plate, allowing the procedure to be performed at a lower temperature, whereas the remaining six fish were dissected at room temperature without the use of cold surfaces. Moreover, to assess the effect of timing on tissue preservation, half of the animals from each condition were fixed in RNA later immediately after organ dissection, while the other half were fixed five minutes after the completion of dissection.
Histology
All samples obtained during dissection were fixed in Bouin’s fluid for 12 h at 4°C with an ethanolic post-fixation (70% ethanol) until further histopathological processing. Posteriorly, samples were dehydrated through a graded series of alcohol to xylol and embedded in paraffin wax. Sections of 2–3 µm thick from paraffin wax blocks were cut and stained with hematoxylin-eosin and periodic acid-Schiff for histopathological analyses. Slides were observed and photographed using an Olympus® BX51 light microscope equipped with a DP72 or EP50 digital camera.
RNA and DNA extraction and bioanalyzer
A total of 235 mid and posterior intestine samples from a larger study, along with 12 mid intestine samples from the trial aimed to evaluate the effect of temperature on sample preservation after dissection, were fixed in RNA later and conserved at room temperature for 24 h and, subsequently, at −20 degrees until processing. Total RNA and DNA were extracted using the E.Z.N.A.® DNA/RNA Isolation Kit, according to the manufacturer’s instructions, and RNA quality was evaluated with the 2100 Bioanalyzer system (Agilent).
Results
This protocol was designed for the sampling of different organs in adult zebrafish, ensuring an efficient and standardized method for high-quality sample collection for subsequent molecular and histological analysis.
Regarding the preservation of the tissue morphology between the dissection protocol here described and those specimens that were fixed whole, the only evident histological difference was found at the intestinal level. The tissue architecture was better preserved in isolated-fixed samples, allowing to see the whole mucosa and preventing the appearance of artifacts, such as epithelial detachment and autolysis of the tips of the intestinal folds (Fig. 8A and B).

Differences in the histology of the intestine between whole fixed specimens and the dissected ones.
Regarding the histological evaluation of samples obtained with or without cold surfaces, no evident histological differences were observed in the intestine or in the other examined tissues.
With regard to the integrity of nucleic acids, the double extraction of RNA/DNA conducted using 235 samples of mid and posterior intestine showed an average RIN (RNA Integrity Number) value of 7.6, with a range from 1 to 10. Of these 235, 85 samples (36%) had RIN values above 8; 99 samples (42%) were in the 7 to 8 range; and 51 samples (22%) fell within the 4.4 to 7 range.
The assay designed to evaluate the potential benefits of using cold plates during dissection showed that samples obtained from fish dissected on cold surfaces had higher RIN values and DNA concentrations. The mean RIN values of samples fixed immediately after organ dissection were similar between conditions (7.73 with cold surfaces vs. 7.00 without). However, greater differences were observed in samples fixed 5 min after complete dissection, where samples exposed to cold surfaces had a mean RIN value of 7.13, compared to 5.53 in those dissected at room temperature.
Regarding DNA concentration and quality, samples obtained on cold surfaces had values of 107.45 ng/µL when fixed immediately after intestine dissection and 107.94 ng/µL when fixed 5 min after complete dissection, with average purity (260/280) ratios of 1.80 and 1.94, respectively. In contrast, samples dissected at room temperature showed lower DNA concentrations of 80.30 ng/µL and 80.74 ng/µL when fixed during and after dissection, respectively, with purity ratios of 1.78 and 1.98.
Briefly, this protocol demonstrated efficiency both in speed, with an average dissection time of 5 min per fish, and in sample preservation. Regarding histopathological studies, it has proven highly effective in preventing tissue degradation (autolysis) and preserving the architecture of the brain, liver, spleen, gonads, kidney, and intestine. Despite the advantages of this protocol, several issues were encountered during the procedure, such as intestinal rupture, mainly in the posterior portion, during traction of the second curvature (Fig. 4A). In addition, small fragments of the liver often remained adhered to the intestinal bulb (Fig. 8C), complicating the separation process and risking cross-contamination between tissues.
Discussion
The dissection protocol described was designed to obtain suitable samples for both histopathological and molecular studies in zebrafish. This technique provides a comprehensive sampling protocol designed to enable the precise individual removal of most zebrafish organs while ensuring optimal tissue preservation for both histopathological and molecular studies.
Zebrafish, like other fish species, lack epidermal keratinization and exhibit greater laxity in their connective tissue compared with mammalian models. 29 This greater cutaneous permeability allows microorganisms to permeate more easily once the animal has died. This fact, combined with the elevated enzymatic activity that persists after death, especially in the gastrointestinal tract, results in rapid tissue decomposition, which can hinder sample collection and compromise the accuracy of results in various studies. 32 , 33 Due to the small size of this model, it is recommendable to perform the dissection under a magnifying glass and adapt the dissection material to the size of the fish, being advisable the use of ophthalmological surgical material, or similar, for enhancing accuracy during the technique. Precision achieved by dissecting each organ individually is particularly important in molecular studies, where samples from individual organs allow for a more faithful evaluation of gene expression of each organ, reducing the potential for distortion caused by tissue mixing. 18 Despite the advantages of this dissection technique, challenges such as the difficulty of separating the liver from the intestine can cause tissue contamination. To avoid this situation, it is necessary to carefully dissect the organ and use forceps to remove liver tissue that may remain attached to the intestinal bulb. Furthermore, the knowledge and experience of the technician or researcher are crucial to obtain accurate results during the technique.
One of the major benefits of this protocol is the ability to maintain the anatomical references of the intestine throughout the dissection process. This is critical in studies involving microbiome analysis, molecular research, and histological or histometric techniques, as it increases the precision of sample collection, yielding more reliable results. To mitigate the possibility of intestinal rupture due to the fragility of this tissue, it is helpful to reduce the depth of the cut with the scissors during the midline cut from the anus when cutting the lateral flap. By keeping the scissors as close as possible to the abdominal wall while cutting, the risk of damaging other organs in the abdominal cavity is significantly reduced. However, despite the rupture of the organ, if the position of the curvatures of the intestine is maintained, it is possible to continue the dissection in the same way as described in the protocol.
Regarding the tissue morphology at the histological level, no evident differences were observed in the samples obtained from zebrafish used to evaluate the effect of cold surfaces on tissue preservation. However, differences in the degree of autolysis were detected when comparing these samples with those from fish that were fixed whole after operculum resection and lateral flap creation. This may be due to the reduced tissues’ surface exposed to the fixative in the case of whole-fixed fish, which limits the fixative’s penetration ability, leading to artifact formation caused by rapid autolysis. 32 , 33
Concerning the stability of nucleic acids, lowering the temperature during dissection by using cold plates allowed for better results in terms of RIN values compared to those samples dissected at room temperature. In the case of samples that were fixed immediately after intestinal dissection, all samples showed a RIN value over 7, which indicates good RNA integrity, crucial to ensure reliable results in gene expression and sequencing analyses. The difference in mean RIN values between both conditions was only 0.7. However, it is important to consider the technician’s skill level and, consequently, the duration of the dissection. The average time required to complete all dissections in this study was five minutes (measured from the positioning of the animal on the sponge to the completion of brain dissection). This timeframe does not appear to be enough to cause significant damage to RNA integrity when performing the dissection at room temperature. The incorporation of additional procedures into the protocol, such as blood extraction following euthanasia, or a lack of technical experience are factors that can slow down sample collection and prolong the overall dissection time. In this study, as demonstrated in the conditions where samples were fixed 5 min post-dissection, cold plates had a positive effect on RNA and DNA preservation. Therefore, considering the possibility that dissection time may be extended due to external factors unrelated to the protocol itself, the use of cold surfaces during dissection is recommended to achieve better results in subsequent molecular analyses.
Note that, although in this protocol samples for histological and molecular analyses were collected separately using Bouin’s fluid and RNAlater, respectively, the tissue fixation and nucleic acid preservation can be performed simultaneously, depending on the sample type and intended downstream applications, by employing specialized molecular fixatives, such as Paxgene® or Histochoice®, 34 although the high cost of these fixatives should be considered.
In summary, this study provides a comprehensive adult zebrafish dissection protocol that enables the precise excision of individual organs while optimizing the preservation of nucleic acid integrity for molecular studies and tissue architecture for histological analyses. The ability to dissect individual organs without contamination is a major improvement over whole-specimen methods, particularly for juvenile zebrafish. While challenges like tissue fragility and organ separation may arise, careful execution minimizes these issues. The use of cold plates may enhance nucleic acid preservation, particularly in cases where less experienced personnel perform the dissection and/or when the sampling process extends over several minutes. Overall, this technique yields high-quality samples, leading to more reliable and precise research outcomes.
Footnotes
Acknowledgment
The authors would like to thank the excellent technical work of Sandra Maceiras and Rita Blanco.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research has received funding from the European Union’s Horizon 2020 research and innovation program under grant agreement No. 101036768 (NeoGiANT project).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.