
Abstract
Limiting the exposure to house dust mite allergens requires integrated approaches and suitable design, and bio-modified textile materials may be used as an element of the complex anti-dust mite protection. The aim of this work was to study the effects of nonwoven systems modified with triclosan on the development of Dermatophagoides farinae house dust mites in laboratory assay. Two four-layer nonwoven polypropylene systems were designed: A – two inner meltblown nonwovens and two outer spun-bonded nonwovens and B – an inner spun-bonded nonwoven, an inner needle-felt nonwoven, an inner meltblown nonwoven, and an outer spun-bonded nonwoven. Bioactive microspheres consisting of poly(L-lactide) (95 wt%) and triclosan (5 wt%) were used. Both systems were modified using dispersion with 2.5 wt% and 10 wt% of microspheres, which corresponds to the content of 0.26 wt% and 1.22 wt% of triclosan in nonwoven samples. The culture tests were performed for modified and unmodified nonwoven samples and the control sample. The anti-dust mite properties were assessed based on the mortality of mites. All modified nonwoven systems showed a strong suppressive effect on the tested Dermatophagoides farinae, compared to the control sample. A higher mortality was observed for nonwoven system B. It was found that a nearly five-fold increase in the triclosan concentration does not lead to a substantial increase in the decline rate of mite populations.
It is estimated that approximately 1–2% of the world human population is affected by an allergy to house dust mites (65–130 million), and the population of people who are allergic to their allergens is still increasing. 1 The results of the third National Health and Nutrition Examination Survey (NHANE S III) show that 27.5% of the US population aged 6–59 years had a positive skin test response to dust mite allergy. 2 Mite allergens are the cause of serious diseases such as bronchial asthma.3–9 In the years 1960–2000, the prevalence of asthma in Australia increased almost twice. House dust mites occur worldwide.1,4,5,10–16 In Europe, the interiors are mainly populated by Dermatophagoides pteronyssinus (DP) and Dermatophagoides farinae (DF), and – to a lesser extent – by Euroglyphus maynei (EM) (Astigmatina, Pyroglyphidae). The Polish flats are mostly inhabited by DF (67%), compared to 17.6% of DP and 1.6% of EM.14,17
Mites of the genus Dermatophagoides primarily inhabit beds and upholstered furniture, and secondarily the flooring and other places that provide them with food and favorable conditions for their development. Air temperature within the range 21–28℃ and relative humidity (RH) of 60–80% make the optimal conditions for the reproduction and development of house dust mites. Below 15℃ and above 35℃, as well as above RH of 80%, their development is slowed down. When the RH drops, the mites sparingly manage water or even use the water coming from their metabolism. Mites draw water from active sorption from the surrounding water vapor, regulating the consumption of water with the supracoxal glands filled with a solution of sodium chloride and potassium chloride.14,18,19 According to Colloff, 1 the water loss from the mites’ bodies reduces the colonization and population growth. The normal water content of DF is usually 71–76% by weight. 19 Under optimum conditions, the adult mites live 4–6 weeks, during which the females lay 40–80 eggs, and the development from an egg to an adult mite takes 3–4 weeks. 1
Limiting the exposure to house dust mite allergens requires integrated approaches. Indoors, dust mites are eliminated with the use of chemical methods,20–22 physical methods,23–28 and combined methods. 29 Chemical acaricides or denaturants (e.g. benzyl benzoate, permethrin derivatives, tannic acid, benzyl alcohol, tannic acid mixture, etc.) are commonly used against dust mites. The multicompounds of selected pesticides have also been tested. 30 In the case of textile materials, the research is focused on designing suitable structures and modification of raw materials and textile goods using different anti-dust mite agents.31–34 Mahakittikun et al. 32 investigated different textile materials and proposed potential parameters for evaluating the anti-mite efficacy of such encasings. It was found that textile floor coverings show anti-dust mite properties through the addition of bioactive agents to the coated backing and applying the same acaricides and anti-adhesive fluorocarbon dispersion on pile fibers. 33 Rahel et al. 34 describe polypropylene fibers modified with chitosan/Ag+ as highly toxic to synanthropic mites. Although many special furnishing materials with anti-mite properties are already produced, such as mattresses, bedding covers, and carpets, development of functional textiles is a promising solution in house dust mite control.
An important habitat for dust mites are devices used to maintain cleanliness, mainly vacuum cleaners, where allergens accumulate after being collected during vacuuming rooms and furnishings.
The aim of this work was to study the effects of bioactive nonwoven systems modified with triclosan on the development of DF house dust mites based on a culture test in laboratory assay.
Material and methods
Textile materials
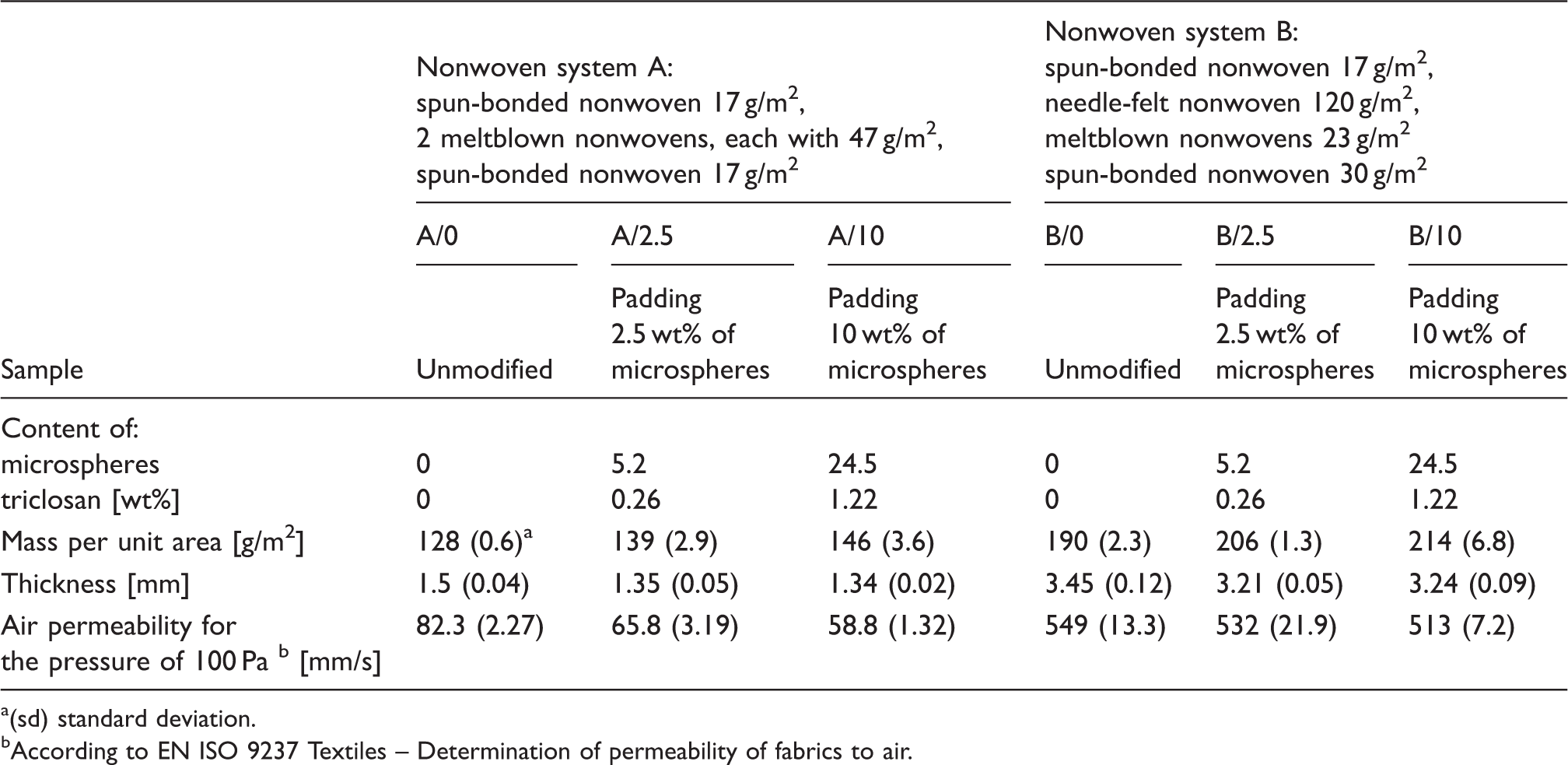
The study was conducted on two four-layer nonwoven polypropylene systems: A – consisting of two inner meltblown nonwovens and two outer spun-bonded nonwovens and B – consisting of an inner spun-bonded nonwoven, an inner needle-felt nonwoven, an inner meltblown nonwoven, and an outer spun-bonded nonwoven. Such types of nonwoven systems are used for vacuum cleaner bags. Polypropylene was used as the material for the nonwoven systems, which is characterized by hydrophobic properties. The normal humidity of polypropylene fibers (determined at 20℃ and 65% RH) is contained within the range 0.01–0.05%, which is associated with virtually zero water retention and lack of sorption swelling.
As a control sample a plain, tightly woven fabric with the mass per unit area of 100 g/m2 was used.
A bioactive modification was carried out for both systems using microspheres prepared by the Center of Molecular and Macromolecular Studies, Polish Academy of Sciences from poly(L-lactide) (95 wt%) with a molecular weight of 10,000 and triclosan (5-chloro-2 -(2,4–dichlorophenoxy)-phenol) (5 wt%) (Figure 1(a)). The bio-modifications of the nonwovens were performed using water dispersion consisting of 2.5 wt% or 10 wt% microspheres and formaldehyde free cross-linking agents (BASF, Germany): Helizarin Binder TX 4738 (10 wt%) and Helizarin Fixing Agent TX 4737 (2 wt%). A two-roll padder (Benz, Switzerland) with roller pressures of 25–35 kg/cm was used. The roller pressures were selected for both types of nonwoven systems so as to keep the same content of microspheres/triclosan. Drying was carried out in a tenter frame oven at a temperature of 90℃ for 4 minutes. The characteristics of the nonwoven systems are given in Table 1 and their microscope images are presented in Figures 1(b)–(d).
Microscopic pictures of bioactive materials used in the study: (a) bioactive microspheres; (b) nonwoven with microspheres; (c) cross-section of sample A: 1, 4 – spun-bonded nonwovens, 2,3 – meltblown nonwovens; (d) cross-section of sample B: 1, 4 – spun-bonded nonwovens, 2 – needle-felt nonwoven, 3 – meltblown nonwoven. Characteristics of the nonwoven systems under the study (sd) standard deviation. According to EN ISO 9237 Textiles – Determination of permeability of fabrics to air.
The culture test
In the experiment, a DF culture was used (Figure 2), which is the most common species of house dust mites in Poland.
35
The test culture had been bred since 2002 in the Department of Parasitology, the Medical University of Silesia in Katowice, as proposed by Arlian et al.
36
(the originator of the test culture is Dr Nigel Hill, PhD, London School of Hygiene and Tropical Medicine, Department of Medical Parasitology, Keppel London, UK). The culture tests were performed according to the heat escape method described by Cieslak et al.
33
on a population kept in the conditions ensuring optimal temperature and RH. The tests were carried out for samples taken from modified and unmodified nonwoven systems (Table 1) and for the control sample – plain tightly fabric. Each sample was placed on a Petri dish with a diameter of 9 cm equipped with a ventilation system. Two hundred mites at different stages of development were settled on each of the 21 samples used. Three repetitions were used for each of the six tested versions of nonwovens and the control fabric. The experimental cultures were kept in chambers stabilized with a saturated sodium chloride solution, at 75% RH and 25℃. The mites were fed with multi-ingredient food “Tropical” Super-Vit Basic Fish Flake Food (Tropical, Chorzow, Poland). The food was replenished whenever its distinct shortage on the dishes was noticed, but always before its complete exhaustion. The overall population growth was observed by identifying the presence of different developmental stages without the assessment of the age structure and live mites in each sample were counted. The experiment was continued until the extinction of the live mites on the modified samples. A minimum of 38 and a maximum of 61 (depending on the sample version) counts of the number of mites for each individual sample were done.
Microscopic documentation of the culture test: (a) Dermatophagoides farinae adult, 190.93 µm × 275.86 µm, 1 – mouth, 2 – supracoxal glands, 3 – lateral opisthosomal gland; (b) Dermatophagoides farinae adult male; (c) egg, 65.00 µm × 43.47 µm; (d) Dermatophagoides farinae protonymph, 1 – one pair of genital papillae; (e) anus, 1 – anal suckers; (f) fecal pellets, 2.86–3.16 µm; (g) dust mite trapped inside the tested sample; (h) Dermatophagoides farinae dehydrated.
Microscopic analysis
The number of live mites in each sample was evaluated daily using a SZ 60 stereomicroscope (Olympus, Japan), while simultaneously making an assessment of the differentiation of developmental stages and eggs, according to Solarz. 37 Larvae were identified (tiny, six-legged), nymphs (smaller than eight-legged adults, without the sexual organs around the hips of the third and fourth pair of legs), and adult individuals (males smaller, mainly heteromorphic with larger legs of the first pair, all with a broad, conical aedeagus (penis) at the ventral idiosomy between the hips of the fourth pair; larger females with smaller first pair of legs, all with a vulva at the ventral idiosomy between the hips of the third and fourth pair of legs). After the completion of the experiment involving the cultures, a scanning electron microscope (SEM) analysis was carried out using an electron microscope VEGA 3 (TESCAN, the Czech Republic) with a specific secondary electron detector LVSTD (Low Vacuum Secondary Tescan Detector). The incident beam energy was 10 keV at the pressure of 20 Pa and the samples were tested without sputtering. To obtain an image of an adult mite in the dry state, a secondary electron detector SE, the incident beam energy 20 keV, and a high-vacuum operation mode 2 × 10−2 Pa were used. The surface of the mites was coated with gold, using a Quorum Technologies Ltd vacuum sputtering system.
Statistical analysis
After the experiment, all daily counted numbers of live mites for each version of the samples were summed and divided by the number of measurements, in order to calculate average values and standard deviations for a mites group bred on each version of the samples. For each group also the median was determined. A Student’s t-test was performed to define statistically significant differences between the mean values of the compared groups. The results were also analyzed with a stepwise regression model. Linear correlation coefficients were calculated according to Pearson’s formula. The value of p < 0.05 was considered to indicate statistical significance.
Results
The microscopic documentation of the culture test is shown in Figures 2(a)–(h). Figures 2(a), (b), and (e) illustrates a male adult that was cultivated at the temperature of 25℃ and the humidity of 75%. The images confirm that optimal culture conditions were provided and the male mite is well hydrated. For comparison, a dehydrated mite in high-vacuum conditions is shown in Figure 2(h). Figures 2(c) and (d) show the presence of developmental stages, respectively eggs and a protonymph, and 2(f) shows fecal pellets, which are the evidence of food intake during the culture test. Dead mites trapped in the spaces between fibers are shown in Figure 2(h).
The population of mites in unmodified samples A0 and B0 developed and there a large number of juvenile stages (larvae, protonymphs, tritonymphs) and eggs was observed. The average number of live mites was the highest in sample A0 and amounted to 273.13; a slightly lower number was observed in sample B0 255.3 (median 300.001 and 260.0, respectively). For the control sample N, the average number of live mites was 318.89 (median 358.0). The lowest number was observed for sample B/10, 52.79 (median 12), followed by sample A/10, 61.1 (median 26.5), sample B/2.5, 69.4 (median 45.0), and A/2.5, 87.73 (median 70.0). In sample B/10, the most marked reduction in the number of mites was noted, including the juvenile stages, and despite the presence of males and females, there was noticeable limitation of reproduction and natural aging of the population. In the case of samples A0 and B0, the flake food was administered four times, and in sample B/10 it was supplemented only once. In general, all samples of nonwoven systems had an adverse effect on the development of the mite population compared to the control sample N. The differences were statistically significant in all cases (p ≤ 0.0000001, the test values (t), and degrees of freedom (df) for each pair compared are as follows: A/2.5 versus N, t = −28.97, df = 328; B/2.5 versus N, t = −30.17, df = 328; A/10 versus N, t = −31.04, df = 328; B/10 versus N, t = −32.46, df = 328; A/0 versus N, t = −5.58, df = 301; B/0 versus N, t = −8.40, df = 301). The negative impact was most pronounced in the case of sample B/10. It was only slightly lower for sample A/10, and this difference was not statistically significant (p = 0.22, t = 1.22, df = 370). The suppressive effect was much lower for both samples with a lower content of triclosan, B/2.5 and A/2.5, and these differences were statistically significant (B/10 versus B/2.5, p = 0.015, t = 2.45; A/10 versus A/2.5, p = 0.0001, t = −4.03; B/10 versus A/2.5, p = 0.0000001, t = −5.37, and df = 370 in all cases), except for the difference between samples B/2.5 and A/10 (p = 0.23, t = 1.20, df = 370). Sample B/2.5 was clearly more effective in reducing the mite population than sample A/2.5, and the difference was statistically significant (p = 0.005, t = −2.79, df = 370). For unmodified A0 and B0 samples, the negative effect was very weak, and the differences between these samples and all modified samples were highly statistically significant (p ≤ 0.0000001, A/0 versus A/2.5, t = −29.06; B/0 versus A/2.5, t = −29.15; A/0 versus B/2.5, t = −30.39; B/0 versus B/2.5, t = −30.44; A/0 versus A/10, t = −31.43; B/0 versus A/10, t = −31.57; A/0 versus B/10, t = −33.26; B/0 versus B/10, t = −33.64, and df = 343 in all cases). A more effective sample in inhibiting the growth of the population was the unmodified sample B0, and this difference, although it does not seem to be high when taking into account the comparison of the average values, was statistically significant (p = 0.0018, t = −3.15, df = 316). The mite populations became extinct first on sample B/10, and finally on sample A/2.5, after 38 and 61 days, respectively (Figure 3).
Changes in the number of live mites during the culture test for modified samples A/2.5, A/10, B/2.5, B/10, and control sample N.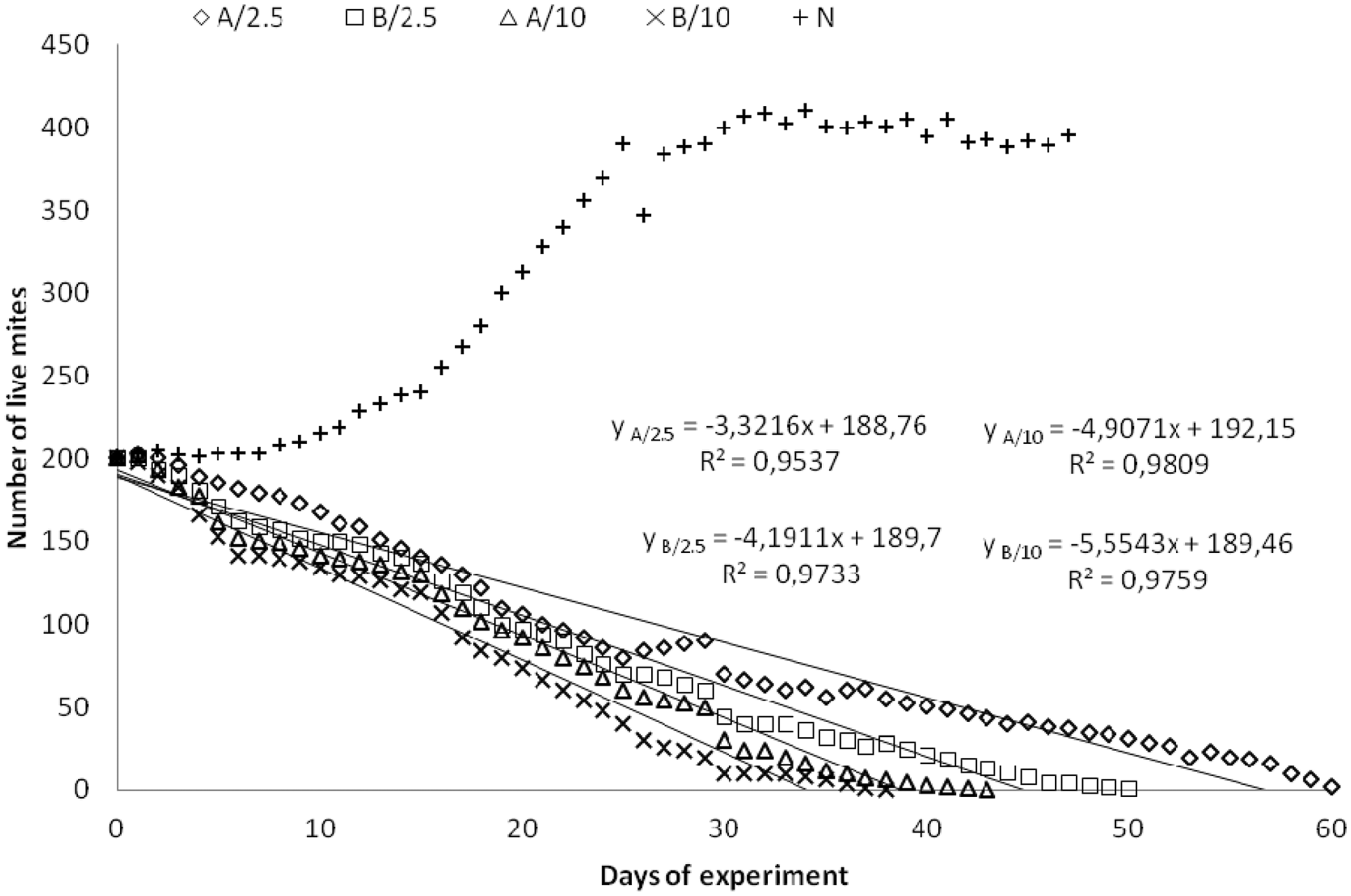
The determined linear correlations (Figure 3) show that the fastest decrease of live mite numbers was observed for sample B/10, followed by samples A/10, B/2.5, and A/2.5. The slopes of straight lines equal respectively −5.5543, −4.9071, −4.1911, and −3.3216. The calculated half-decline rate of the population is respectively 16.1, 18.8, 21.4, and 26.7 days. It was found that a nearly five-fold increase in the triclosan concentration (from 0.26 wt% to 1.22 wt%, Table 1) does not lead to a substantial increase in the mortality of mites. The ratios of the slopes of A/10 to A/2.5 and of B/10 to B/2.5 equal 1.47 and 1.33, respectively. Similarly, the ratios of half-decline rates for the above-mentioned pair of samples are very close to each other. It can be seen that among the samples with a higher microsphere content, the difference in the values of these coefficients is lower than that for samples with a lower content, equaling 2.67 and 5.32, respectively. This proves the predominant effect of high concentrations of triclosan in these samples. In samples with lower concentrations, there was a noticeable impact of the structure of nonwovens on the survival of mites. This is confirmed by the results of a culture test conducted for the unmodified samples A/0 and B/0 (Figure 4). After 30 days of the experiment, the number of mites on the control sample N stabilized (Figures 3 and 4(b)), whereas it was still decreasing on the unmodified samples. The constants of the population decline rate obtained from exponential formula (1), describing the kinetic curves of live mite decrease, are −0.005 and −0.013 day−1, respectively (Figure 4(b)):
Changes in the number of live mites during the culture test for unmodified samples A/0, B/0, and control sample N: (a) first 30 days; (b) after 30th day.

The structure of system A is tighter than that of system B. The values of mass per unit area to the thickness ratio are 0.085 and 0.055, and the air permeability values are 82.3 and 549 [mm/s], respectively. Mites penetrated system B more easily, and then – weakened by triclosan – were left trapped in the spaces between the fibers and died in them. This was confirmed by the microscopic analysis performed for both systems after the experiment and particularly relates to layer 2 of system B – the needle-felt nonwoven (Figure 2(g)). For this reason, using the lower concentration of triclosan and nonwoven system B seems to be the better and environmentally advantageous solution.
Discussion
Optimal conditions for the reproduction and development at all stages in the life cycle of DF include the availability of proper food, RH at the level of 75%, and temperature of 27℃. 38 It is widely recognized that in the natural conditions, the main food of house dust mites from the family Pyroglyphidae are human and animal skin scales. However, it was not proven sufficiently that it is the basis of the diet from which nutrients are absorbed. Skin scales consist mainly of keratin and lipids. They are also covered with a secretion of the sebum, bacteria, and yeasts, and they contain triglycerides and cholesterol. 1 Keratin itself does not constitute a balanced diet. As was demonstrated by De Saint Georges-Gridelet, 39 lack of vitamin B6 has a negative effect on the house dust mite population development. Studies also showed that the addition of yeasts significantly accelerates the growth of the population.40,41 The way in which mites assimilate nutrients is still under investigation. 1 It is believed that a big role in their diet can be played by fungi that pre-digest the food for them. Erban and Hubert 42 hypothesize that the efficient use of nutrients from the skin or hair is possible through the symbiotic interaction of mites with fungi, as well as keratinolytic bacteria and collagenolytic bacteria. The activity of proteolytic enzymes – keratinase and collagenase – of these microorganisms allows for the absorption of nutrients from the skin. The type of digestive enzymes the individual mites are equipped with depends on their food needs. DF contain enzymes with different biochemical functions, 1 which allow the mites to absorb various nutrients, providing them with a balanced diet and promoting proper development of the population. The enzymes catalyze the process of proteolysis, leading to hydrolytic degradation of the peptide bonds, and thus the breakdown of proteins into peptides and amino acids. In addition to this, they are capable of destroying the cell walls of fungi and bacteria, they are involved in the transport of cholesterol, etc. On the one hand, bacteria and fungi are engaged in facilitating the process in which mites assimilate nutrients; on the other hand, they themselves make the food the mites feed on. Therefore, the elimination of these microorganisms affects, directly and indirectly, the feeding process, and thus the development of populations of DF. The cultured mites had constant access to food, providing a balanced diet, which is confirmed by the results of the assessment of the population growing on the control sample, on which the population increased exponentially. The reduction of the population in the modified samples was induced by the presence of triclosan in the microspheres.
Triclosan is a chlorinated, aromatic chemical compound with two functional groups characteristic for ethers and phenols, which determine its antimicrobial activity. It is non-volatile at 20℃ and at the pressure of 4 × 10−6 mmHg, slightly soluble in water (0.01 g/dm3 at 20℃), and better soluble in strong alkali solutions, ethanol, and diethyl ether. Triclosan is characterized by a broad spectrum of antimicrobial activity, 43 and it is active against most Gram-positive and Gram-negative bacteria as well as certain fungi and protozoa. It shows, inter alia, high efficacy (MIC 0.01–0.1 mg/dm3) against bacteria from the genera of Staphylococcus, Streptococcus, and Mycobacterium. 44 It was found that triclosan manifests reducing activity against Staphylococcus aureus45,46 and Escherichia coli. 47 At low concentrations, it shows a bacteriostatic effect by blocking the synthesis of lipids necessary for the reproduction and construction of the cell membrane; at higher concentrations (several percent), it causes destabilization of the membranes resulting in a rapid bactericidal effect.43,48,49 Some authors believe that triclosan affects the food chain, mainly due to its lipophilic properties (log KOW = 4.8). 50 The use of triclosan, as well as other bioactive substances, should be controlled.51,52 Triclosan has been used worldwide in many consumer, medical, and cosmetics products. Despite numerous research works devoted to triclosan, its toxicological assessment and safety evaluation is not conclusive.53–55 Queckenberg et al. 54 evaluated the pharmacokinetics and safety of triclosan after dermal application of cream with 2% triclosan and concluded that it is safe for use in hydrophobic creams. One of the important factors is, inter alia, the selection of the process for its application and the minimum concentration ensuring the long-term biological activity of the modified material and suitable management of the post-consumer product. In the study, microspheres were used that are durably bonded to the fiber surface, and control released triclosan – due to the lack of solubility in water and the hydrophobic nature of the polypropylene fibers – exhibits activity against dust mites within the structure of the nonwovens. Using nonwoven systems containing 0.26 wt% of triclosan provides effective activity against the test mite population in the laboratory assay.
Conclusions
Two structurally different polypropylene nonwoven systems modified with bioactive microspheres consisting of 95 wt% of poly(L-lactide) and 5 wt% of triclosan were developed. Two contents of triclosan in both nonwoven systems were used. The optimal climatic and nutritional conditions were ensured for the test culture and the impact of the nonwoven systems on the survival and development of the population of house dust mites DF was assessed. All tested systems showed a negative impact on the development of the test mite populations, compared to the control sample. The adverse impact on the population was definitely stronger (statistically significant, p ≤ 0.0000001) in the case of modified nonwoven systems than in the case of the unmodified ones. System B, built with internal needle-felt and meltblown nonwovens and external spun-bonded nonwovens, modified with water dispersion containing 10 wt% microspheres (1.22 wt% of triclosan in the textile sample) reduced the test population most effectively. However, a nearly five-fold increase in the triclosan content (from 0.26 to 1.22 wt%) does not lead to a substantial increase in the decline rate of mites populations. Using the nonwoven system B modified with water dispersion containing 2.5 wt% microspheres (0.26 wt% triclosan in the textile sample) seems to be the adequate and environmentally advantageous solution. The developed nonwoven systems can be mainly used for vacuum cleaner bags, but also as underlayers in beds, carpets, and upholstered furniture as textile elements of the complex anti-dust mite protection.
Footnotes
Funding
This work was carried out within the Project – POIG.01.03.01-00-004/08 Functional nano- and micro textile materials – NANOMITEX co-financed by the European Union with the financial resources of the European Regional Development Fund and the National Centre for Research and Development, Poland.
Acknowledgements
The authors would like to thank Bogna Goetzendorf-Grabowska, Zenon Polus, and Bogdan Gajdzicki for their co-operation and help.