
Abstract
Studies indicate that the benefits of volunteering may extend to biological risk factors in disease development including chronic inflammation, though the pathway through which volunteer activity predicts chronic inflammation remains unclear. The current project focuses on the link between volunteering and C-reactive protein (CRP) as a measure of chronic inflammation, while paying a particular attention to sleep quality as a pathway. Using panel data from the National Social Life, Health, and Aging Project (N = 1,124), the present study examined whether sleep quality operates indirectly linking volunteer activity and CRP (indirect pathway), compensates for the lack of volunteerism (moderation-compensation), or regulates the benefits of volunteering on CRP (moderation-regulation). The findings suggest sleep quality as a compensatory pathway, in that sufficient sleep buffers the inflammatory effect of lack of volunteerism. The findings show that helping others may be beneficial for the helpers in terms of chronic inflammation and sleep quality as interconnected health outcomes.
Volunteerism in later life is extolled as salutary for the individuals, those served, and the community at large. Formal volunteering is defined as an unpaid activity that contributes to others beyond the volunteer’s own household (Morrow-Howell, 2010). At the individual level, volunteer activities affect a range of physical and mental health outcomes including psychological well-being (Glass et al., 2006; Matz-Costa et al., 2014), cardiovascular disease(CVD; Han et al., 2018), functional limitations (Okun et al., 2010), and all-cause mortality (Glass et al., 1999). The findings lend strong support for the successful aging model pioneered by Rowe and Kahn (1997) who suggested that the health implications of participating in social and voluntary activities are distinct from simply maintaining social relationships.
Recently, studies indicated that the benefits of volunteering may extend to important biological risk factors in disease development process including chronic inflammation (Han et al., 2018; Kim & Ferraro, 2014). Chronically elevated inflammation is linked to greater risks of heart attack, stroke, and mortality (Crimmins & Vasunilashorn, 2011) and is modifiable. It is shown that maintaining ties to a larger community is related to lower chronic inflammation (Loucks et al., 2006). Accordingly, inflammatory markers present an important way to explore links between how social engagement and volunteering, in particular, gets “under the skin” and affects physiological processes. Despite a growing body of research on volunteering and inflammation, the pathway through which volunteerism may influence inflammatory biomarkers remains unclear.
There may be multiple potential pathways linking volunteer activities and inflammatory process, including a stress response to environmental challenges, lower resting blood pressure, and sympathetic nervous system (SNS) activity (Uchino et al., 1996). One of the potential and understudied pathways is sleep quality. Quantity and quality of sleep is affected by social activities during the waking hours, including social interactions (Friedman et al., 2005) exercise, and volunteer activity (Basner et al., 2007). In both laboratory and large-scale epidemiological studies, poor sleep or sleep deprivation is shown to increase chronic inflammation (Cho et al., 2015; Irwin et al., 2010). A recent prospective study shows that 1-hr reduction in sleep is associated with 8.1% higher C-reactive protein (CRP) and 4.5% higher interleukin-6 (IL-6) levels 5 years later (Ferrie et al., 2013). The findings yield significant clinical implications for better sleep quality because even smaller fluctuations in chronic inflammation can be detrimental to CVD outcomes in older adults (Ridker et al., 2000). Capitalizing on sleep quality and inflammation as relatively modifiable health outcomes for older adults, the current project focuses on the relationship between volunteering and chronic inflammation, while paying a particular attention to sleep quality as a linking pathway. Two distinct pathways are hypothesized: The indirect pathway specifies that volunteerism has both direct and indirect effects on chronic inflammation through better sleep quality, and the moderating pathway stipulates that the link between voluntary activity and inflammation is contingent upon better sleep quality.
Volunteering, Sleep Quality, and Chronic Inflammation
The current project is informed by three interrelated lines of inquiry. First line of inquiry focuses on how volunteer activity affects sleep quality and inflammation. Volunteering involves both physical activity and social integration (i.e., giving and receiving support) that are linked to better health and sleep quality (Basner et al., 2007; Glass et al., 2006). In particular, research using American Time Use Survey shows that physically active lifestyle and volunteer activities influence greater sleep duration (Basner et al., 2007). Activities that are explicitly directed toward helping others in need offer unique health benefits. Models of compassionate neurobiology and caregiving system (Brown & Okun, 2013) shed theoretical light on why volunteerism may affect both sleep quality and inflammation. Humans have an evolved caregiving system that enables us to care and want to promote well-being of others. Such other-directed helping behaviors involve a cascade of biochemical events that may reduce stress and inflammation, from the activation of medial optic area of the hypothalamus (mPOA) to the release of oxytocin (OT) and progesterone (Preston, 2013). Both OT and progesterone are anti-inflammatory agents. At the same time, mPOA region is known to be responsible for sleep maintenance, bodily temperature, and CVD functional regulation during sleep (Mitchell & Weinshenker, 2010). Sleep, in this respect, is an innovative avenue for studying a range of helping behaviors and chronic inflammation. There has not been a study to our knowledge that examined volunteer activity, sleep quality, and chronic inflammation in one overarching model.
Second, converging evidence suggests that the anti-inflammatory effects of better sleep quality may be a mechanism linking volunteering and cardiovascular health. Sleep is a quintessential restorative behavior during which one detoxifies substances accumulated when awake and restores energy levels for the next day. Sleep quality is particularly germane for older adults since more than half of adults over 65 report at least one chronic sleep complaint (Benloucif et al., 2004; Strike & Steptoe, 2003). Laboratory studies show that sleep deprivation or poor sleep quality affects inflammatory messenger RNA (Irwin et al., 2006), CRP (Meier-Ewert et al., 2004; van Leeuwen et al., 2009), IL-6, and tumor necrosis factor-α (TNF-α; Irwin et al., 2010). In community-dwelling older adults, both subjective and objective measures of sleep quality were negatively associated with inflammatory proteins measured by CRP, IL-6, E-selectin, and TNF-α (Friedman, 2011; Prather et al., 2009; Strike & Steptoe, 2003). Taken together, the findings indicate sleep as a critical health behavior that is linked to the key inflammatory markers.
Finally, research establishes chronic inflammation as a potential predictor of CVD mortality (Crimmins & Vasunilashorn, 2011; Ridker et al., 2000), accountable for one in four deaths in the United States. The most common risk factor for developing CVD is atherosclerosis, and inflammatory proteins such as CRP play a key role in arterial plaque formation, thereby greatly increasing the susceptibility to myocardial ischemia and infarction (Bennett et al., 2013). In fact, CRP is an independent predictor of CVD even after adjusting for major risk factors such as total cholesterol level, body mass index (BMI), and diabetes. It has been also shown that chronic inflammation is related to stroke and cancer development (Bennett et al., 2013). Thus, chronic inflammation is a valuable biomarker for the current project.
Sleep as the Indirect Pathway
Helping behaviors (i.e., volunteering) improve older adults’ subjective sleep quality and regulate the sleep cycle in both experimental and population-based studies (Benloucif et al., 2004; Friedman, 2011; Rohleder et al., 2012; Uchino, 2006). Although the exact pathway underlying this link is not well-understood, there is evidence that activation of caregiving system through voluntary activity sets in motion a set of hormonal events that has restorative properties on the body and downregulates inflammation (Brown & Brown, 2015). The mPOA region activates when a person engages in helping behaviors, which may enhance the restorative effects of sleep. Several studies support this link, showing that individuals’ social environments contribute to better sleep quality such as intimate relationships (Kiecolt-Glaser et al., 2010), social engagement, and leisure activities (Basner et al., 2007; Lin et al., 2012). In addition, sleep quality has dramatic effects on inflammation. Laboratory studies show that 5–10 days of sleep deprivation (i.e., 4 hr of sleep per night) led to more than a 5-fold increase in CRP, IL-6, and TNF-α even during the recovery period (Ewert et al., 2004; van Leeuwen et al., 2009). On the contrary, those who had normal sleep patterns exhibited no changes in CRP. Taken together, scholars point to sleep quality as a potential restorative pathway through which volunteerism may influence circulating inflammatory markers (Rohleder et al., 2012). Studying this pathway also offers practical values for health practitioners and researchers to develop interventions for new and existing volunteers, targeting specific physical health outcomes. Thus, the current project hypothesizes that volunteerism may affect CRP both directly and indirectly through sleep quality.
The Moderating Pathway
According to the caregiving system model, repeated activation of caregiving system tends to downregulate stress and chronic inflammation, resulting in lower levels of inflammation (Brown & Brown, 2015). However, given the inflammatory effects of poor sleep, older volunteers reporting suboptimal sleep quality, such as sleep deprivation and lack of restfulness upon waking, may not experience health benefits of volunteer activities. On the other hand, if less active volunteers exhibit high quality of sleep, it may compensate for the lack of volunteer activity. In order to test this pathway, we propose two alternative moderation hypotheses. First, a compensation pathway postulates that lack of sleep (or poor sleep quality) can be compensated by active volunteering and vice versa. This hypothesis is supported by the research on positive social relationships, sleep quality, and inflammation (measured by IL-6 and E-selectin) such that better sleep quality can offset the lack of positive social relationships on inflammation (Friedman, 2011). Alternatively, sleep as a regulation hypothesizes that the benefits of volunteerism on CRP are contingent upon the quality of sleep. It may be that the positive effects of volunteering on chronic inflammation manifest only to the extent to which older adults report adequate sleep. Highly active volunteers who report poor sleep quality may not derive the health benefits from such activities.
Hypotheses
Guided by the caregiving system model and empirical evidence, the current project investigates whether sleep quality directly and indirectly affects CRP, compensates for the lack of volunteer activity and vice versa (moderation-compensation), or regulates the benefits of volunteering on CRP (moderation-regulation). Although inflammation and sleep study were considered as separate health outcomes in studies, considering both as interrelated is essential for understanding how volunteerism affects health in later life.
Method
The project used the first (W1) and second (W2) waves of data from the National Social Life, Health, and Aging Project (NSHAP), an ongoing nationally representative longitudinal study of 3,005 older adults residing in the United States. NSHAP W1 was collected in 2005–2006 and comprised of older adults aged 58 to 85. The initial response rate was 75%. During the interview, a random 83% sample (n = 2,494) was selected to undergo blood spot collection, of which 2,120 respondents agreed to provide sample (85% response rate). Williams and McDade (2009) showed that those who did not provide blood samples were comparable to those who agreed by age, gender, race, income, education, marital status, self-rated health, and number of doctor visits in the past year. Additional 181 cases were discarded due to reasons such as insufficient volume of blood or equipment malfunctions. As is standard in the literature, 209 cases with very high levels of CRP (>8.6 mg/L), indicative of inflammatory response to acute conditions such as flu or injury, were excluded (Herd et al., 2012). By 2010–2011, when W2 data were collected, 220 respondents had died; 88 were in very poor health or in a care facility, and an additional 94 respondents were not reinterviewed due to various reasons including refusal or lack of contact information. Of 1,328 eligible interviewees, 167 respondents who had very high levels of CRP (>8.6 mg/L) were excluded from the analyses. Finally, 37 cases had missing data on control variables; they were omitted from the sample. Thus, the final analytic sample size is 1,124.
Measures
Endogenous variables
CRP Concentration
In the NSHAP, during the face-to-face interview at both waves, a blood sample was collected via capillary finger stick and disposable lancet; up to five drops of blood were applied to filter paper for transport and storage. Blood-spot assays were completed at the Laboratory for Human Biology Research at Northwestern University (Williams & McDade, 2009). Given the highly positively skewed distribution of CRP (few individuals have higher levels of CRP), the values were natural log transformed.
Subjective sleep quality
Sleep quality is based on three self-reported items that were asked of all respondents at both waves. Sleep duration at W1 was assessed with a single item “How many hours do you usually sleep at night?” with the response options in whole hours. At W2, instead of reporting the hours of sleep in general, participants responded with clock time on the questions “What time do you usually go to bed and start trying to fall asleep?” and “What time do you usually wake up?” The questions were asked separately for weekdays (workdays) and weekends (days off). Average sleep duration was calculated from bedtime and wake time questions for weeknights and weekend (weighted 5/7 and 2/7). Sufficient sleep was defined as 6 or more hours of sleep on average, consistent with the literature (Dowd et al., 2011). Responses were coded as 0 (≥6 hr) and 1 (<6 hr). A sleep quality item was included in the scale for depressive symptoms, in which participants were asked whether their “sleep was restless” with four levels of response endorsement from rarely or none of the time (0) to most of the time (3) with higher number indicating more restless sleep. Overall sleep satisfaction was evaluated with the question “How often do you feel really rested when you wake up in the morning?” with the response categories ranging from 0 (never) to 3 (most of the time) with higher number indicating more rested sleep. Detailed information on sleep quality data collection and measures are available in Lauderdale et al. (2014). Although sleep quality was measured differently at W1 and W2, we treated them as relatively comparable based on prior studies conducted by the principal investigators (Chen et al., 2016).
Exogenous variable
Volunteer activity
Volunteer activity measures were available in the analytic sample at both W1 and W2. Respondents answered how often they volunteered for religious, charitable, political, health-related, and other organizations in the past 12 months. Reponses ranged from never (0) to several times a week (6).
Additional covariates
In addition to the variables of central interests, several covariates were considered because of their documented links with sleep quality and chronic inflammation (Herd et al., 2012; Kim & Ferraro, 2014). All covariates were measured at W1. Age is coded in years, and sex is dichotomized with 1 indicating female. Race was divided into a series of binary variables (White, Black, and Other race) with non-Hispanic White serving as a reference category. Other racial groups consisted of American Indian or Alaskan Native, Asian or Pacific Islander, and those who identified themselves as Other races. Marital status is a binary variable with 1 indicating married or cohabiting with a partner. In order to measure socioeconomic status, four categories of education (less than high school, high school graduate, some college, and bachelor’s degree or more) and low net worth were included in the analysis. For low net worth, respondents were asked to estimate their net worth including all of their investments, properties, and other financial assets minus debt. A binary variable was created distinguishing respondents in the lowest 20% of the household net worth from those in the top 80%.
For health-related life style factors considered to predict CRP, we used self-reported information. Tobacco use was defined by current consumption of cigarettes, pipes, cigars, or chewing tobacco. Physical activity was measured with an item probing respondents’ frequency of physical activities such as walking, dancing, or exercise (0 = never to 4 = 2 or 3 times per week). Binary variable for obesity was defined by calculating BMI with interviewer-measured height and weight (>30 kg/m2).
Additional analyses included other covariates (i.e., income, controlled and uncontrolled hypertension, former smoking, underweight, overweight, religious service attendance, network size, total volume of contact, self-reported physical/mental health, emphysema, asthma, arthritis, diabetes, center for epidemiologic studies depression scale (CES-D), and lipid lowering medication) and alternative coding of variables (i.e., continuous measure of net worth). These were omitted from the final analyses, however, because they were not significant in any of the multivariate specifications.
Analyses
The analyses used generalized structural equation models (GSEM) in order to include multiple paths in one model (MacKinnon et al., 2007). GSEM offers several advantages for testing the indirect effects in the current project because it allows for the estimation of the association between voluntary activity and sleep quality, comprised of multiple ordinal items, and the link between sleep quality and CRP is evaluated in the same model as opposed to the traditional multistep (Skrondal & Rabe-Hesketh, 2004). GSEM also allows the distribution of outcome variables to be dichotomous, ordinal (e.g., sleep quality), or log transformed (e.g., CRP). We estimated cross-lagged panel models in order to account for possible bidirectionality (Finkel, 1995). W1 volunteer activity was used to predict W2 CRP both directly and indirectly through W2 sleep quality, controlling for all other covariates as well as W1 CRP and sleep quality.
The current project employed multiple approaches in order to test the moderation effects. First, in order to test the multiplicative effects of latent sleep quality construct (categorical variable) and a categorical volunteer activity, the analysis used MIXTURE and KNOWNCLASS commands in Mplus 8.4 (Muthen & Muthen, 2012). Second, the analyses modeled the changes in CRP (by regressing W2 CRP on W1 CRP) as well as W1 predictors as lagged predictors with and without the multiplicative terms between volunteering and each sleep quality measure. The additive model (model with variables specified as independent predictors) and interaction models were compared using the log likelihood test. If the results for the log likelihood ratio test are statistically significant (p < .05), it indicates that the model with an interaction term is better than the model without an interaction term. Third, multigroup models were estimated with each sleep quality measure as grouping variables in order to determine whether the path between frequent volunteerism and CRP differs in multiple sleep quality groups. A fully constrained model where all paths were constrained to be identical for multiple sleep quality groups was estimated; then, the path between volunteer activity and CRP was allowed to be free between poor and adequate sleep quality groups. Nested model tests reveal whether the constrained or group-specific model is better.
Finally, in the NSHAP, reinterview rates were high in the follow-up study (i.e., 87.8% of the NSHAP W1 survivors reinterviewed) but sample attrition may nonetheless produce biased parameter estimates in longitudinal analyses. Since the majority of attrition was due to death, Heckman’s (1979) selection bias models were employed to adjust for the differential selectivity due to mortality. We first estimated a probit model to distinguish respondents who participated at the follow-up interview from those who died. Predictors of mortality in the probit model included age, female, and tobacco use along with several variables that were not in the substantive equation predicting CRP (i.e., difficulty in activities of daily living, underweight [BMI ≤ 18.5], and self-rated health). The selection instrument (λ) based on the inverse Mills ratio was subsequently estimated and included in all substantive models.
Result
Table 1 shows the descriptive statistics for study variables in the NSHAP. At W1, the average log CRP level was 0.12 (corresponding raw CRP value 1.82 mg/L). Respondents reported good sleep quality in general, with 88% reporting at least an adequate amount of sleep (6 hr or longer) and 12% reporting less than 6 hr of sleep. On average, the respondents rarely reported restless sleep (mean = 0.87, median = 1) and often reported that they felt rested in the morning (mean = 2.48, median = 3). The mean volunteering frequency was 2.37 (median = 2), between several times a year and once a month. At W2, the average inflammation level was 0.53 (raw CRP = 2.46 mg/L). In terms of sleep quality, 96% reported adequate amount of sleep (6 hr or longer), while 4% reported insufficient sleep. On average, sleep quality showed no drastic changes between waves. The mean for restless sleep was 0.83 (median = 1), and the mean for restful sleep was 2.55 (median = 3). The NSHAP respondents were still active in volunteering (mean = 2.33, median = 2).
Descriptive Statistics of Study Variables.

Note. N = 1,124. CRP = C-reactive protein.
Main Effects Model
Table 2 shows the results from cross-lagged GSEM predicting volunteering, sleep quality, and CRP at W2. Sleep quality as a latent variable consisted of insufficient sleep, the frequency of restless sleep, and the lack of restful sleep. Model 1 included the main effects of volunteering at W1 on sleep quality and CRP. This model yielded good model fit indices (root mean square error of approximation [RMSEA] = .02, comparative fit index [CFI] = 0.98, Tucker-Lewis index [TLI] = .96). Contrary to the hypotheses, volunteer activity did not have significant effects on sleep quality or CRP at W2. Sleep quality at W1 was significantly associated with CRP at W2 (β = −.07, p < .05).
Generalized Cross-Lagged Model Predicting Volunteerism, Sleep Quality, and CRP.
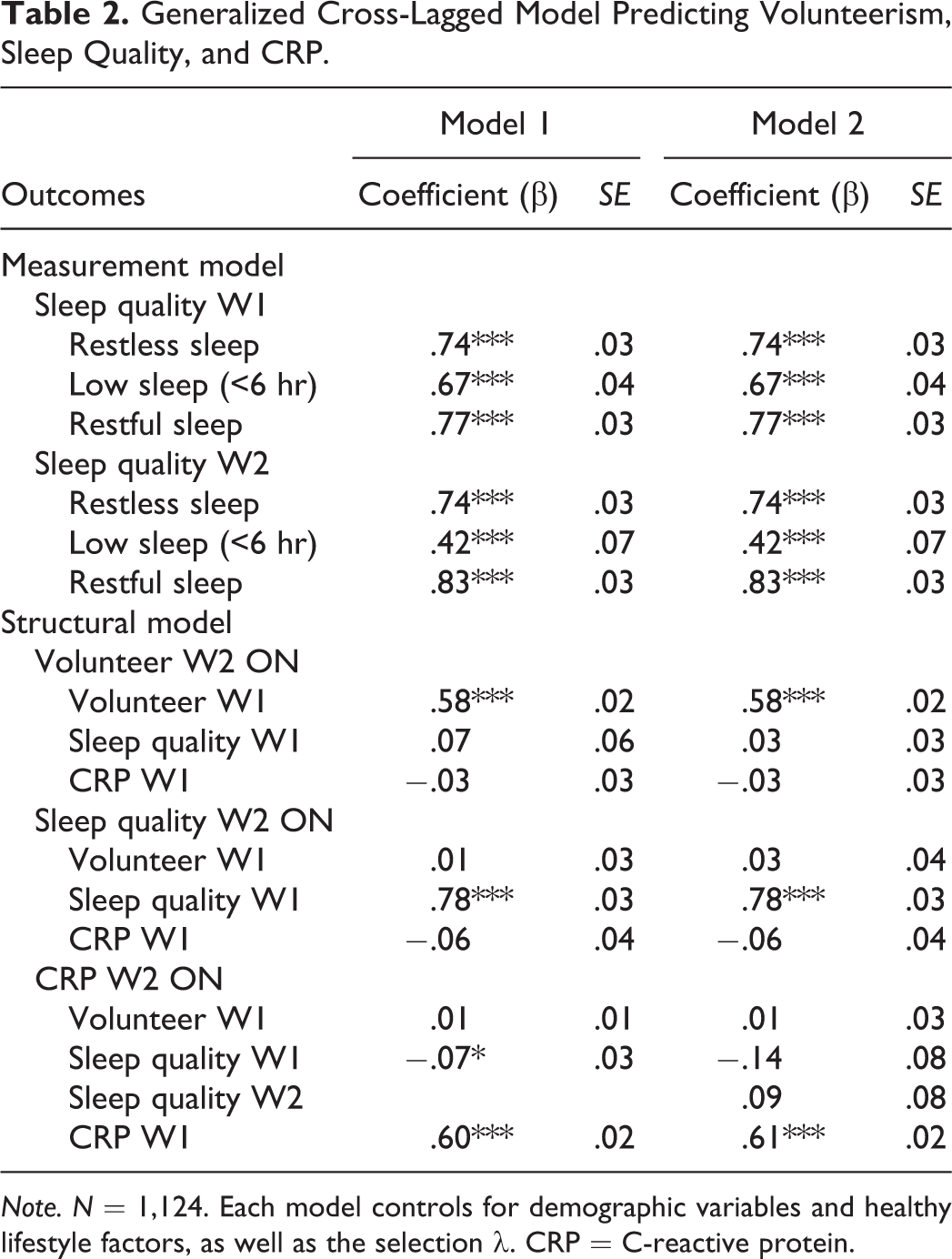
Note. N = 1,124. Each model controls for demographic variables and healthy lifestyle factors, as well as the selection λ. CRP = C-reactive protein.
Testing the Indirect Effect of Sleep Quality
Model 2 in Table 2 tested the hypothesis by adding a path between W2 sleep quality and W2 CRP. The model showed adequate fit indices (RMSEA = .03, CFI = 0.97, TLI = .95). The results showed, however, that sleep quality did not act as an indirect pathway linking voluntary activities and chronic inflammation. The standardized indirect effect was 0.002 and not statistically significant. In a supplementary analysis, an alternative indirect model was tested with W1 sleep quality (instead of W2). Volunteering at W1 was not significantly associated with sleep quality at W1 (β = .06, p = .08), but sleep quality at W1 was linked to CRP at W2 (β = .07, p < .05). The substantive conclusions remained unchanged.
Testing the Moderation Effect of Sleep Quality
The subsequent models tested the moderating effects of sleep quality. The first method, a multiplicative term between sleep quality as a latent variable and volunteer activity, was not statistically significant. In investigating this effect further, the next set of analyses tested combinations between each sleep quality measure and volunteer activity (e.g., Volunteering × Insufficient sleep, Volunteering × Rested sleep, Volunteering × Restless sleep) in separate models. The additive model (model with variables specified as independent predictors) and interaction models were compared using the log likelihood test. Results show that an interaction term between frequent volunteer activity and sufficient sleep (6 hr or more) was significant in predicting log CRP (b = .01, β = .02, p < .05). The log likelihood ratio test was significant in favor of the alternative model (d = 11.89, p < .01). The multiplicative model had an adequate fit (RMSEA = .01, CFI = 1.00, TLI = .98).
In corroborating this finding, group-specific analysis was conducted between insufficient and sufficient sleepers. There were 135 individuals who reported insufficient sleep. A fully constrained model where all paths were constrained to be identical for groups was estimated; then, the path between volunteering and log CRP was allowed to vary between low and adequate sleep groups. This model yielded adequate fit indices (RMSEA = .02, CFI = 0.95, TLI = .93). Group-specific analysis showed that frequent volunteer activity was a significant predictor of log CRP only for older adults who reported insufficient sleep (i.e., <6 hr; b = −.03, β = −.06, p < .05). Log likelihood ratio test revealed that a model with a group-specific path between volunteering and log CRP was better in terms of the model fit (d = 5.58, p < .05). The other combinations of voluntary activities and sleep quality measures were not significant. In order to aid the interpretation, the results from the analyses were plotted in Figure 1. The figure shows that those who report 6 or more hours of sleep on average do not really derive extra benefits from volunteering. In contrast, in support of the compensatory hypothesis, older adults who reported insufficient sleep (<6 hr) had a negative line along the categories of volunteering frequency. Adding the group-specific paths also explained 7% more variance in log CRP (0.384 vs. 0.391).
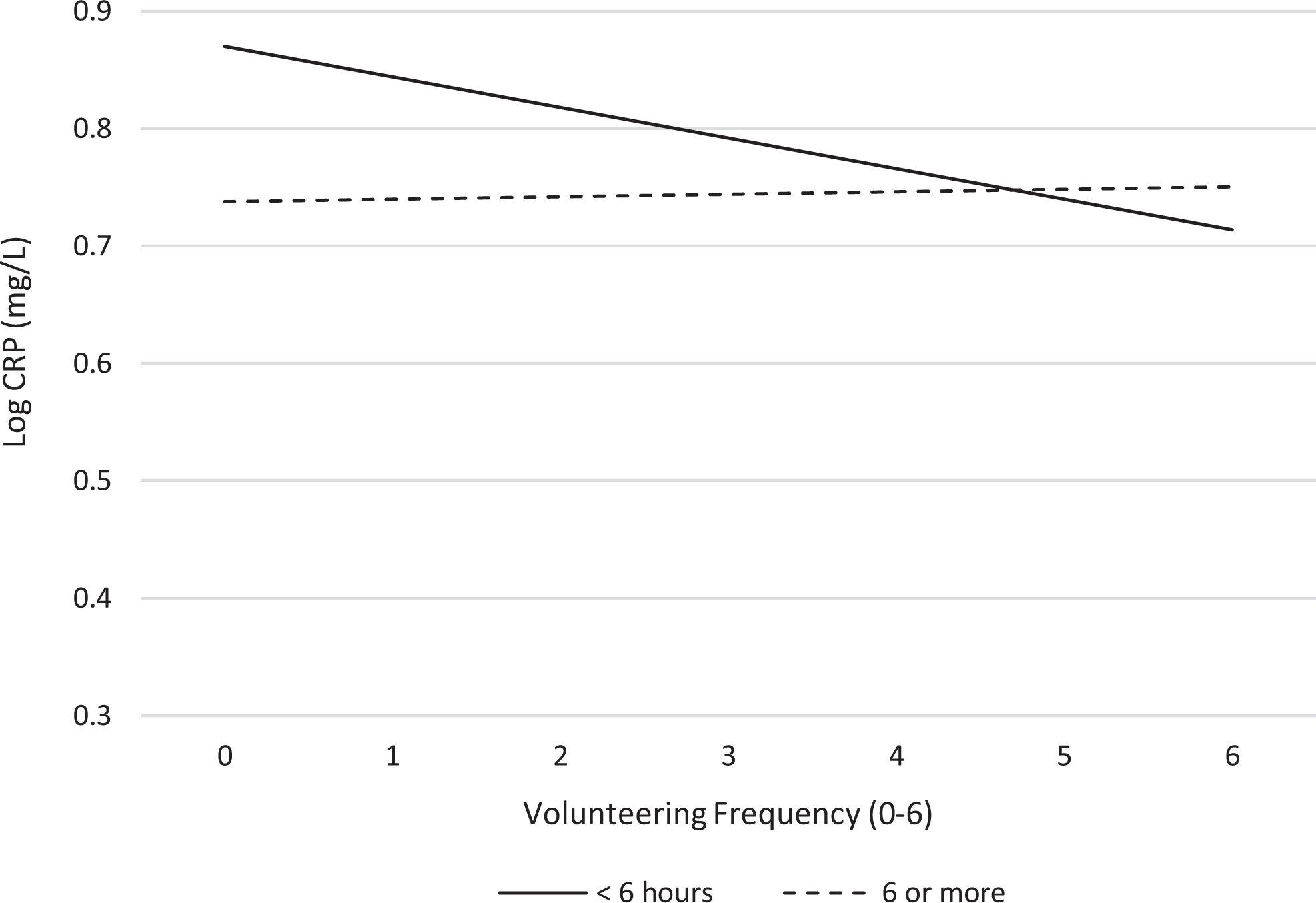
Volunteering and C-reactive protein (CRP), stratified by sleep duration. Note. Results adjusted for all covariates, nonrandom attrition effects, and W1 CRP.
Discussion
Using nationally representative sample of older adults in the United States, the current project extends the caregiving systems model by investigating sleep quality as a potential mechanism through which voluntary activity is linked to chronic inflammation. A number of studies report that voluntary activities and helping behaviors are associated with lower chronic inflammation (Kim & Ferraro, 2014) and better sleep quality (Benloucif et al., 2004; Rohleder et al., 2012), but no study to date investigated how voluntary activities and sleep quality conjointly influence chronic inflammation. The results support sleep quality as a compensatory pathway, revealing that frequency volunteer activity may be protective against CRP for older adults who report insufficient sleep. The lowest level of CRP was evident among older adults who reported both sufficient sleep and frequent volunteering, adjusting for covariates, baseline inflammation, and selection effects.
What are potential explanations for sleep as a compensatory pathway on chronic inflammation? The caregiving systems model posits that voluntary helping behaviors are likely anti-inflammatory through neurobiological benefits such as dampened stress response (Brown & Okun, 2013), decreases in SNS activity (Eisenberger & Cole, 2012), lower blood pressure (Inagaki & Eisenberger, 2016), and increases in OT (Brown & Brown, 2015). Given that sleep as a restorative behavior offers similar “quieting” effects on the central nervous system, sufficient sleep may compensate for lack of volunteering and vice versa in the context of chronic inflammation. This is not to argue that sufficient sleep or volunteering alone is a panacea for chronic inflammation, but rather to suggest that voluntary activities may offset some of the physiological damages from lack of sleep by activating caregiving systems. Further, the findings signify the importance to consider multiple related health outcomes (i.e., sleep quality and chronic inflammation) simultaneously rather than separately. Although not the core focus of the current study, additional psychological correlates of volunteering such as perceptions of feeling needed (Gruenewald et al., 2012), generativity (Moieni et al., 2018), and self-esteem (O’Donnell et al., 2008) would explain which psychological benefits of voluntary activity may exert the most anti-inflammatory effects. Motivations for volunteering are also important predictors of health outcomes, such as altruism, social connection (Konrath et al., 2012), and valuing others (Poulin, 2014). Future studies should also directly test which neural mechanisms are activated during different types of volunteering activity.
Sleep quality did not indirectly influence the relationship between voluntary activities and inflammation in older adults in the present analyses. The finding is in line with the extant literature that positive social relations and inflammation in older adults are not indirectly affected by sleep quality (Friedman et al., 2005). Interestingly, multiplicative terms between volunteering and other sleep measures were not significant in the model. It may be that sleep deprivation is the key contributor to the inflammatogenic process rather than feeling rested or restless upon sleep. One study demonstrates that habitually low sleep duration is associated with various adverse health outcomes including heightened inflammation, hypertension, and coronary heart disease (Troxel, 2010). Other studies explored the proinflammatory effects of more serious or pathological sleep outcomes such as insomnia (Cho et al., 2015) or obstructive sleep apnea (Vgontzas et al., 2009), indicating that moderate sleep issues (e.g., not feeling rested in the morning) might not be potent enough to fluctuate inflammation level in prospective data. Thus, a key to developing successful interventions is to target insufficient sleep as a health outcome among volunteers.
To date, there has been lack of longitudinal study testing whether older adults’ volunteer activity affects chronic inflammation, and how sleep quality might moderate or show indirect effects on this relationship. The cross-lagged model suggests that models including single measurement for CRP may overestimate the association between volunteerism and chronic inflammation. Thus, use of prospective panel data provides convincing argument on the health benefits of volunteering, as well as some information on the directionality, albeit using two waves. The findings highlight the significance of developing interventions to facilitate better sleep quality in older adults, and particularly among older volunteers. In particular, given psychological and physical benefits of participating in community volunteer activities, integrating volunteer activity-based components into interventions can be a possibly favorable and easily accessible means of preventing and treating sleep problems among older adults.
The findings should be interpreted with caution due to the following limitations. The assessment of sleep quality relied upon self-reports, and the measurements of sleep duration differed across the two waves. Even though the core variable of interest was sufficient sleep (6 hr or more), this discrepancy in the two sleep duration measures is a limitation. More objective measures of sleep quality are needed in order to further understand the role of sleep in an inflammatogenic process. Some objective sleep quality measures include sleep fragmentation (percentage of sleep spent waking and percentage less than 5 min of immobile sleep), sleep architecture (percentage of slow-wave sleep), or sleep efficiency (percentage of actual sleep by time spent in bed; Lauderdale et al., 2014). Second, though we utilized longitudinal data and adjusted for W1 CRP, nonrandom selection effects, healthy lifestyle factors, and chronic conditions that are associated with inflammation, there could be other health conditions that confound the relationship between volunteer activity, sleep quality, and inflammatory markers. Thus, it is impossible to completely parse out the issue of reserve causality. A causal findings of sleep quality and chronic inflammation is consistent with randomized controlled trials (Meier-Ewert et al., 2004) but could not be definitively confirmed from findings in the current project. Future studies may use three or more waves in order to formally test mediation, as well as the changes in volunteer activities, sleep quality, and chronic inflammation.
In conclusion, the current study extends the findings from cross-sectional data and provides the moderating effects of sleep quality on the link between volunteer activity and inflammation, measured by CRP. Given the role of inflammation and sleep deprivation in the disease development process and its modifiability, the findings suggest that voluntary activity may be a potential avenue for individuals to stay healthy and active in later life.
Footnotes
Author’s Note
Hyunwoo Yoon is now affiliated with School of Social Work, Portland State University, Portland.
Acknowledgments
The authors appreciate valuable comments from Sara Mustillo, Elliot Friedman, Kenneth Ferraro, and Patricia Thomas.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.