
Abstract
Endometrial carcinoma is the most common gynecologic malignancy is the United States and accounts for 6% of all cancers in women. The disease is classified as type I or type II based on clinicopathologic and molecular features. It is a multifactorial disease with a number of risk factors, including environmental exposures. How environmental exposures, such as flame retardants, may affect the incidence of endometrial cancer is a topic of current and ongoing interest. Tetrabromobisphenol A (TBBPA) is a widely used brominated flame retardant found in a variety of household products. A recent 2-year National Toxicology Program carcinogenicity study found that exposure to TBBPA was associated with a marked increase in the development of uterine tumors, specifically uterine carcinomas, in Wistar Han rats. Molecularly, TBBPA-induced uterine carcinomas in Wistar Han rats were characterized by a marked increase in tumor protein 53 mutation compared to spontaneous uterine carcinomas, as well as overexpression of human epidermal growth factor receptor 2. Similar to spontaneous carcinomas, tumors in TBBPA-exposed rats were estrogen receptor-alpha positive and progesterone receptor negative by immunohistochemistry. The morphologic and molecular features of uterine carcinomas in TBBPA-exposed rats resemble those of high-grade type I tumors in women, and these data suggest that exposure to TBBPA may pose an increased cancer risk.
Introduction
Endometrial cancer (EC) is one of the leading causes of cancer morbidity and mortality in women in the United States, with an estimated 49,560 new cases and 8,190 deaths in 2013, and even higher estimates for 2014 (American Cancer Society [ACS] 2013, 2014). EC in women is classified into 2 subtypes, type I and type II, based on clinicopathologic and molecular features. These 2 subtypes of EC have unique dualistic phenotypic and molecular features that correlate relatively well with the biologic behavior of these neoplasms (Horn et al. 2007; Lax 2004; Liu 2007; Llobet et al. 2009; Matias-Guiu et al. 2001; Ryan et al. 2005). Type I ECs account for 80 to 90% of cases and generally have a more favorable prognosis, while type II ECs, which represent approximately 10 to 20% of cases, are associated with a poor prognosis. While the features of the two EC subtypes may overlap to some extent, subtyping is of particular utility when determining clinical outcome, prognosis, or therapeutic options for these tumors.
Based on clinicopathologic features, type I tumors occur predominately in postmenopausal women, although they are also less frequently reported in premenopausal and perimenopausal women. Type I tumors are more often associated with nulliparity, are estrogen receptor-alpha (ERα) and progesterone receptor (PR) positive, and are slowly progressive with a lower risk of myometrial invasion (Mills and Longacre 2011). Alternatively, type II ECs occur in postmenopausal women, are associated with multiparity, and are high grade with an aggressive clinical course. Morphologically, type I tumors have endometrioid differentiation, characterized by irregularly shaped, variably sized glandular structures forming a merged, cribiform pattern or papillary proliferations, and associated with a desmoplastic stroma (Horn et al. 2007). Type I ECs are typically low grade and arise within a background of atypical hyperplasia, but high-grade type I tumors do occur that behave more aggressively. Type II ECs (nonendometrioid) are much more poorly differentiated, characterized by tubulocystic, papillary, or solid forms composed of small, cuboidal to “hobnail-” shaped cells with pleomorphic nuclei, frequent mitoses, and often bizarre nuclear forms. Necrosis may be common. Type II ECs are subdivided into a “clear cell” subtype based on cytoplasmic glycogen, or a “serous” subtype based on eosinophilic cytoplasm with apically located nuclei and focal cytoplasmic clearing. Features of both phenotypes may be present in the same tumor. Type II ECs arise within an estrogen-deficient, atrophic uterine epithelium and are associated with a precursor lesion of endometrial intraepithelial carcinoma. They are highly aggressive and infiltrative tumors, often spreading along the peritoneal cavity, and harbor a poor prognosis.
Aside from clinical and morphologic features, ECs in women can also be subclassified based on molecular phenotype (Horn et al. 2007; Lax 2004; Liu 2007; Llobet et al. 2009; Matias-Guiu et al. 2001; Ryan et al. 2005). Type I EC is associated with a higher incidence of phosphatase and tensin homolog (PTEN) inactivation, microsatellite instability, and mutation of the Kirsten rat sarcoma viral oncogene homolog (KRAS), Catenin (cadherin-associated protein), beta 1 (CTNNB1), and phosphatidylinositol-4,5-bisphosphate 3-kinase, catalytic subunit alpha (PIK3CA) genes. Conversely, Type II EC is associated with a very low incidence of the abovementioned molecular alterations, but rather has a very high incidence of tumor protein 53 (TP53) mutation, as well as overexpression of human epidermal growth factor receptor 2 (HER2), Cyclin D1 (CCND1), and Cyclins D1/E1 (CCND/E1); inactivation of cyclin-dependent kinase inhibitor 2A (P16); and alterations in E-cadherin (CDH1), expression.
While all type II ECs are considered high grade, an accepted and widely used histologic grading system has been developed for type I ECs to distinguish more aggressive, high-grade type I tumors from low-grade type I tumors. High-grade type I ECs have overlapping morphologic and molecular features of type I and type II tumors, including increased local invasion, loss of ERα and PR expression, increased KI67 protein expression, high rates of TP53 mutation, and a poor prognosis, relative to low-grade type I ECs (Liu 2007; McConechy et al. 2012; Zannoni et al. 2010). Table 1 lists some of the clinicopathologic and molecular features of the subtypes of ECs (Horn et al. 2007; Liu 2007; McConechy et al. 2012; Ryan et al. 2005; Zannoni et al. 2010).
Clinicopathologic and molecular features of human EC subtypes.
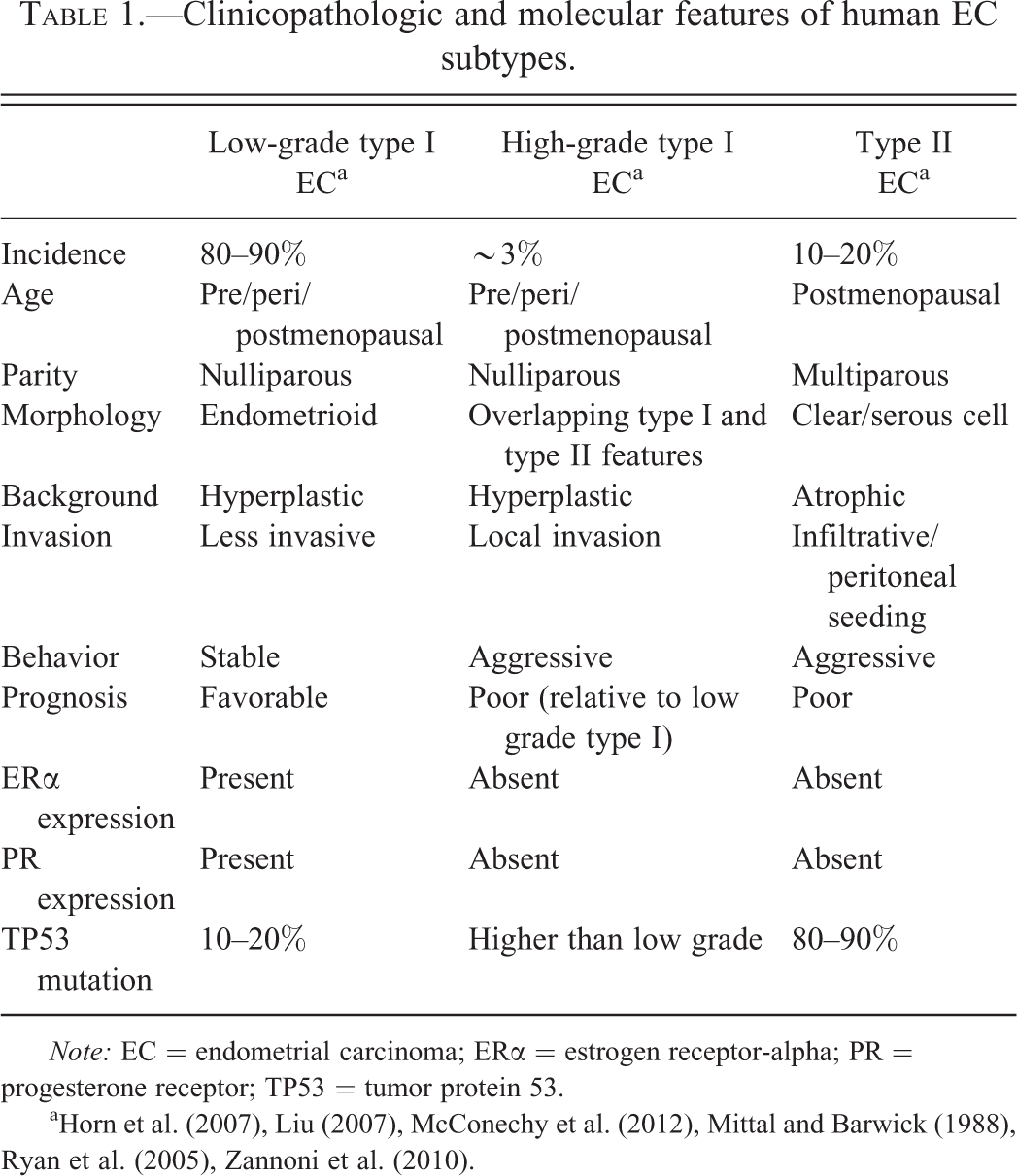
Note: EC = endometrial carcinoma; ERα = estrogen receptor-alpha; PR = progesterone receptor; TP53 = tumor protein 53.
A major risk factor for EC in women is hormone dysregulation. Many of the contributing factors to the development of EC influence estrogen levels, and increased estrogen is associated with EC development. Hormone (estrogen) therapy in menopausal women is associated with increased risk (Beral, Bull, and Reeves 2005), whereas the use of oral contraceptives and multiparity are protective (Modan et al. 2001). Early menarche and late menopause are risk factors, and nulliparity is a risk factor, since pregnancy shifts the balance of hormones from estrogen to progesterone (Emons et al. 2000). Other risk factors include family history, diabetes, obesity, tamoxifen therapy, and ovarian pathologies such as granulosa cell tumors and polycystic ovarian syndrome, which are associated with unabated estrogen secretion (Emons et al. 2000). Aside from family history, the majority, if not all of these risk factors, are associated with hormone imbalance. It is becoming evident that an increasing number of compounds that people are exposed to in their environment, whether they are environmental pollutants or occupational compounds, have the ability to influence hormone production and function.
Tetrabromobisphenol A (TBBPA), a compound structurally related to bisphenol A and formed by bromination of bisphenol A, is a widely used flame retardant in electrical equipment, plastics, paper, furniture, and carpeting. It is a high-volume production chemical in the United States and the most widely used brominated flame retardant in the world (Schauer et al. 2006). It has been identified in fish, shellfish, and drinking water. Like bisphenol A, TBBPA has been shown to have endocrine disrupting activity in vitro and in vivo, and in the uterotropic assay in ovariectomized mice, exposure to 500-mg/kg TBBPA resulted in increased uterus:body weight ratios (Kitamura et al. 2005). It is known that estrogens, estrogenic chemicals, and estrogen agonists can act as tumor promoters as well as carcinogens (Vollmer 2003). Previous studies have shown that TBBPA is possibly a very weak estrogenic agonist in vitro, but results have been equivocal (Hamers et al. 2006). It seems to stabilize and prevent turnover of estrogen levels through inhibition of estrogen sulfotransferase, a hormone metabolizing enzyme (Gosavi et al. 2013). However, there is little known about the long-term toxicologic and carcinogenic effects of TBBPA in humans. In a recent National Toxicology Program (NTP) 2-year carcinogenicity study, exposure of Wistar Han rats to TBBPA by corn-oil gavage was associated with a treatment-related increase in uterine carcinomas (NTP TR 587, 2013). Given the relevance of this compound to human exposures and the potential for endocrine disruption, investigation of the molecular mechanisms of uterine tumorigenesis due to TBBPA exposure in Wistar Han rats was warranted. The objective of this study was to characterize uterine carcinomas in Wistar Han rats exposed to TBBPA based on histopathologic and molecular criteria relevant to the human disease, to better understand the similarities and differences between human and rat EC, and to better understand the molecular events that occur in uterine carcinomas from TBBPA-treated Wistar Han rats.
Materials and Methods
Animals, Histology, and Immunohistochemistry
TBBPA-induced uterine carcinomas were obtained from Wistar Han [Crl:WI(Han)] rats in the 2-year NTP bioassay. The care of animals on this study was according to National Institutes of Health (NIH) procedures as described in the “The U.S. Public Health Service Policy on Humane Care and Use of Laboratory Animals,” available from the Office of Laboratory Animal Welfare, National Institutes of Health, Department of Health and Human Services, RKLI, Suite 360, MSC 7982, 6705 Rockledge Drive, Bethesda, MD 20892-7982 or online at http://grants.nih.gov/grants/olaw/olaw.htm#pol. The use of animals was approved by the National Institute of Environmental Health Sciences (NIEHS) animal care and use committee. Forty-one formalin-fixed, paraffin-embedded (FFPE) uterine carcinomas from TBBPA-exposed animals were used in this study (10 low dose, 15 mid dose, and 16 high dose). A total of 10 FFPE spontaneous carcinomas were available from control animals from various NTP studies. Due to the low incidence of spontaneously occurring uterine carcinomas in the current TBBPA carcinogenicity bioassay (2 spontaneous tumors in corn-oil vehicle gavage control group), additional FFPE spontaneous tumors were obtained from other available Wistar Han rat carcinogenicity bioassay studies: one from polybrominated diethyl ether mixture (DE-71-NTP TR 589, 2014d; corn-oil vehicle gavage), 2 from green tea extract (GTE-NTP TR 585, 2014c; deionized water vehicle), 2 from metal working fluids (MWF-NTP TR 586, 2014b; whole body inhalation), and 3 from antimony trioxide (AT-NTP TR 590, 2014a; whole body inhalation). Five FFPE normal uteri from corn-oil vehicle gavage control females in the concurrent TBBPA study were used as normal uterus controls. Four board-certified anatomic veterinary pathologists reviewed the histology of all uterine carcinomas using the standardized system of nomenclature and diagnostic criteria (SSNDC) guide for female reproductive proliferative lesions in rats to morphologically characterize the endometrial adenocarcinomas as well as differentiated or poorly differentiated tumors (Table 2). If a tumor had areas that were well differentiated and poorly differentiated, then the tumor was classified as poorly differentiated. Tumors available for molecular biology included those >0.5 cm at the time of necropsy. These tumors were sectioned in half, and one half was flash frozen in liquid nitrogen for molecular analysis and the other half fixed in 10% neutral buffered formalin (NBF) for histology and immunohistochemistry (IHC). Frozen tissue was available from 6 uterine carcinomas from TBBPA-exposed animals, and 9 vehicle control animals from the abovementioned studies (1 from TBBPA, 2 from GTE, 3 from AT, 2 from DE71, and 1 from MWF). The abovementioned frozen tissues and paired FFPE tissues were used for quantitative polymerase chain reaction (qPCR) and IHC analysis, respectively. For IHC, 5-μm paraffin-embedded sections were obtained and immunolabeled using the Vector ABC system with the following antibodies using routine methods: ERα (1:50, Beckman Coulter Inc., Fullerton, CA), PR (1:50, Beckman Coulter Inc., Fullerton CA), TP53 (1:500, Vector Laboratories, Burlingame, CA), and KI67 (1:100, Dakocytomation Corporation, Carpinteria, CA). The expression of TP53 protein in uterine carcinomas was graded by a board-certified pathologist based on a severity grade of 0 to 3 (0 = no immunoreactivity, 1+ = minimal, 2+ = moderate, 3+ = strong immunoreactivity). KI67 was graded based on a scale of 0 = no immunoreactivity, 1+ = <25%, 2+ = 25–50%, and 3+ = >50% of tumor cell immunoreactivity.
SSNDC classification of endometrial adenocarcinomas.

Source: Dixon et al. (2014).
Note: SSNDC = standardized system of nomenclature and diagnostic criteria.
DNA Isolation, Polymerase Chain Reaction (PCR) Amplification, and Autosequencing
DNA was isolated and extracted from 22 FFPE TBBPA-induced uterine carcinomas and 10 FFPE spontaneous uterine carcinomas from rat control animals with DNeasy Tissue Kit (Qiagen, Valencia, CA). Amplification reactions were carried out by semi-nested PCR using the designed primer sets (Table 3) for Kras exons 1 and 2 (codons 12, 13, 61), Ctnnb1 exon 2 (corresponds to exon 3 in human), Tp53 exons 5 to 8, and Pten exons 1 to 9 for 6 cDNA-treated samples. Controls lacking DNA were run with all sets of reactions. PCR products were purified using a QIAquick Gel Extraction Kit (Qiagen, Valencia, CA). The purified PCR products were cycled with Terminal Ready Reaction Mix-Big Dye (Perkin Elmer, Foster City, CA), and the extension products were purified using the DyeEx 2.0 Spin Kit (Qiagen, Valencia, CA). The lyophilized PCR products were sequenced with an automatic sequencer (Perkin-Elmer ABI Model 3100, Waltham, MA). An electropherogram was used for comparison between normal uterus from vehicle controls and uterine carcinomas from vehicle controls and TBBPA-exposed animals.
Primers used to amplify hotspot regions of rat Kras, Ctnnb1, Tp53, and Pten.
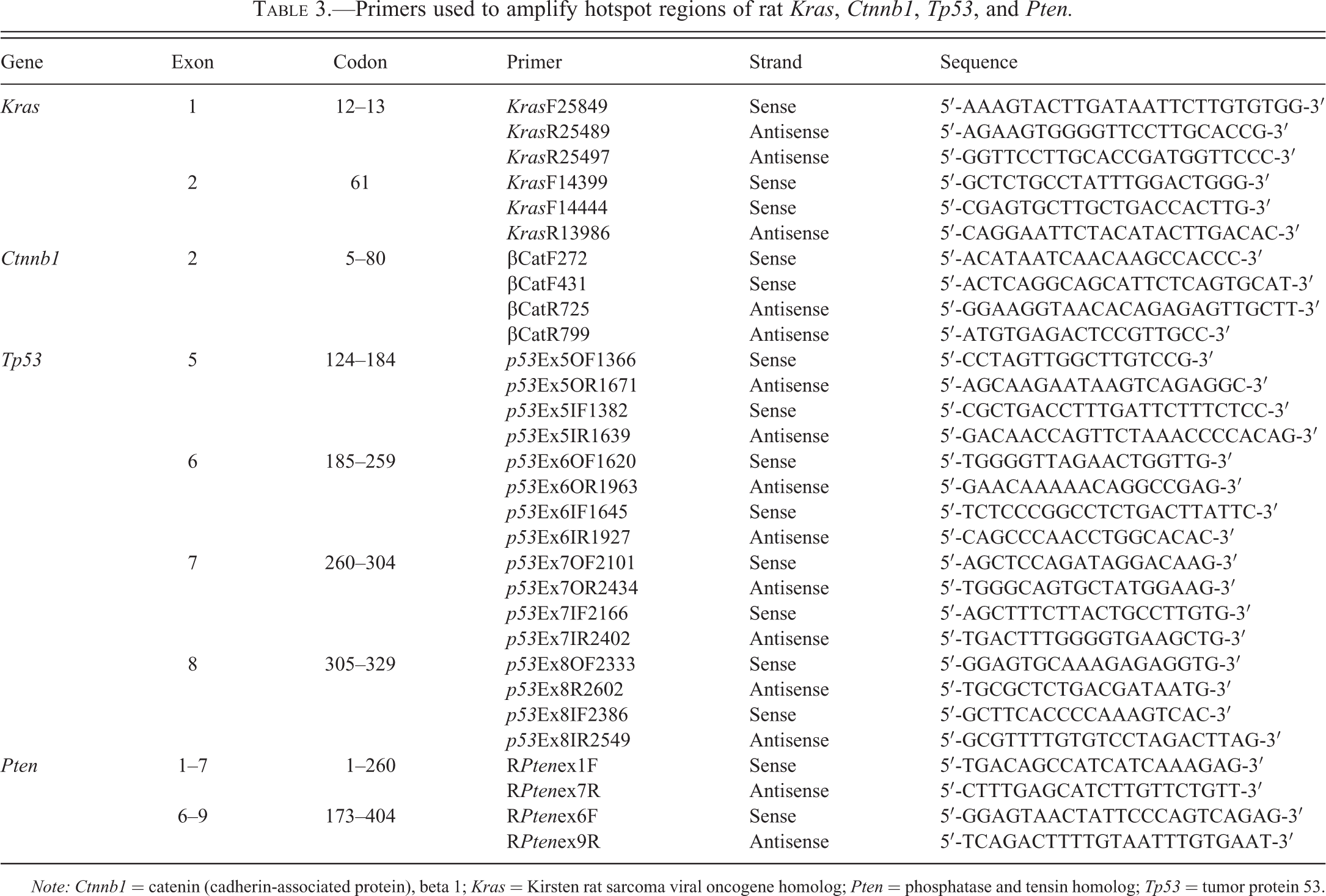
Note: Ctnnb1 = catenin (cadherin-associated protein), beta 1; Kras = Kirsten rat sarcoma viral oncogene homolog; Pten = phosphatase and tensin homolog; Tp53 = tumor protein 53.
qPCR
RNA was extracted and purified from frozen samples using the Invitrogen TRIzol Kit (Invitrogen, Carlsbad, CA). Fold increases and decreases in gene expression were determined by quantification of cDNA from target samples relative to a calibrator sample of controls. The 18s RNA gene was used as the endogenous control for normalization of initial RNA levels, and the expression of genes relevant for human EC (P16, Her2, Ccnd1, Cdh1, MutS protein homolog 2 [Msh2], MutS protein homolog 3[Msh3], MutL homolog 1 [Mlh1], interferon regulatory factor 1 [Irf1], Pten) was assessed.
Statistical Analysis
Incidences of neoplasms and nonneoplastic lesions were tested using one-sided poly-3 tests (Bailer and Portier 1988; Portier and Bailer 1989; Piegorsch and Bailer 1997). This method modifies the Cochran–Armitage linear trend test and Fisher’s exact test to take length of survival into account, by more closely approximating the total number of animal-years at risk. The poly-3 trend test assesses the significance of the linear trend of incidences with dose, while the pairwise poly-3 tests assesses the significance of pairwise differences in incidences of each dose group compared to the control group. Differences are considered statistically significant if the one-sided p value is less than .05.
Results
Exposure to TBBPA was associated with a treatment-related increase in uterine carcinomas in Wistar Han rats. The results of the 2-year NTP chronic bioassay indicated that TBBPA exposure resulted in a statistically significant (p < .05) increase in the incidence of uterine carcinomas in the top 2 dose levels of female Wistar Han rats (Table 4). Based on histomorphologic criteria (Table 2), both TBBPA-induced (28 of 41, 68%) and spontaneous (7 of 10, 70%) uterine carcinomas in Wistar Han rats were classified as poorly differentiated, high-grade carcinomas, characterized by deeply infiltrative lobules, cords, and nests of poorly differentiated epithelial cells with marked cellular pleomorphism, atypia, high mitotic rate, and large foci of necrosis (Figure 1A–B and Table 5). The remaining carcinomas were considered fairly well differentiated, as endometrial stromal invasion was limited to the superficial and mid-endometrium rather than replacing large areas of the deep stroma. Additionally, neoplastic glands exhibited milder nuclear atypia and pleomorphism as well as less necrosis (Figure 1C–D).
Incidence of epithelial uterine neoplasms in TBBPA-exposed Wistar Han rats in the 2-year NTP bioassay.
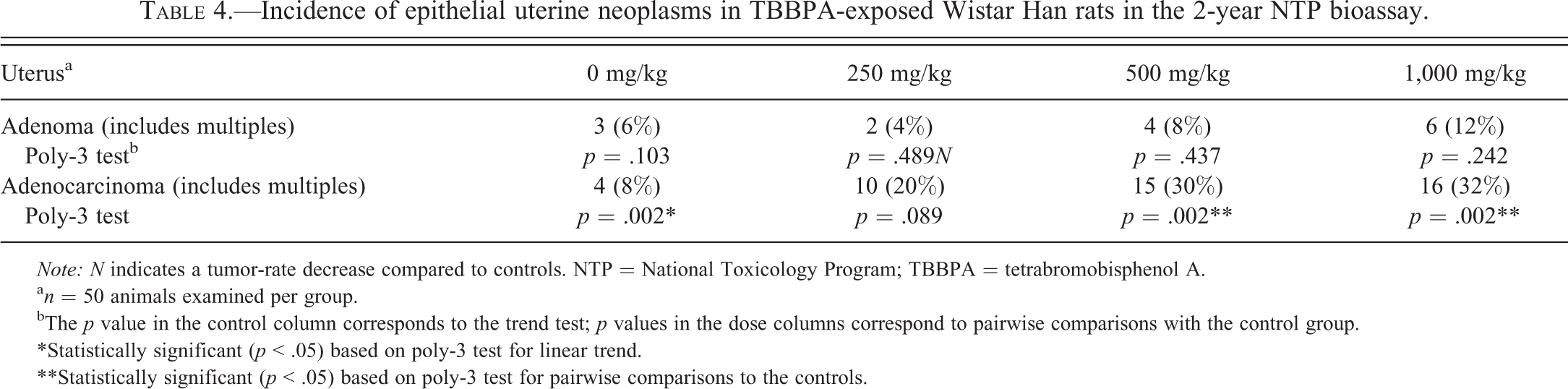
Note: N indicates a tumor-rate decrease compared to controls. NTP = National Toxicology Program; TBBPA = tetrabromobisphenol A.
an = 50 animals examined per group.
bThe p value in the control column corresponds to the trend test; p values in the dose columns correspond to pairwise comparisons with the control group.
*Statistically significant (p < .05) based on poly-3 test for linear trend.
**Statistically significant (p < .05) based on poly-3 test for pairwise comparisons to the controls.

Uterine carcinomas from tetrabromobisphenol A (TBBPA)-exposed Wistar Han rats. (A) Poorly differentiated uterine carcinoma characterized by marked invasion through the uterine wall into the peri-uterine tissues, accompanied by large zones of necrosis. (B) Poorly differentiated uterine carcinomas were characterized by solid clusters, cords, and nests of poorly differentiated epithelial cells with marked pleomorphism and cellular atypia, including large vesicular nuclei, multiple nucleoli, marked anisocytosis and anisokaryosis (inset), and mitotic figures. (C) Well-differentiated carcinomas were fairly well circumscribed with stromal invasion relatively limited to the superficial and mid-endometrium rather than replacing large areas of the deep stroma and a glandular phenotype. (D) Glands were generally lined by one or more layers of fairly well-differentiated cuboidal to columnar epithelial cells with minimal to mild pleomorphism and few mitotic figures.
Morphologic and immunohistochemical features of uterine carcinomas from control and TBBPA-exposed WH rats.

Note: TBBPA= tetrabromobisphenol A; TP53 = tumor protein 53; WH = Wistar Han.
aValues inside brackets represent average intensity scores. This score is an average of the immunoreactivity severity grade (0 = no immunoreactivity, 1+ = minimal, 2+ = moderate, 3+ = strong immunoreactivity) for each animal divided by the total number of animals in each cohort (n = 6 for TBBPA-treated rats; n = 9 for control rats).
Uterine carcinomas in TBBPA-exposed Wistar Han rats show a tendency toward loss of PR expression and increased TP53 protein expression compared to spontaneous uterine carcinomas. Since criteria for classifying human type I and II endometrial carcinoma relies partly on hormone (ERα/PR) expression, proliferation, and Tp53 mutations, uterine carcinomas from TBBPA-exposed animals and spontaneous uterine carcinomas from vehicle controls and controls were examined for expression of ERα, PR, KI67, and Tp53 by immunohistochemistry. All TBBPA-exposed (6 of 6) and spontaneous (9 of 9) uterine carcinomas in Wistar Han rats examined had diffuse positive nuclear immunoreactivity to antibodies against ERα. All spontaneous and TBBPA-exposed uterine carcinomas in animals had nuclear immunoreactivity in the endometrial stroma and myometrium, similar to the staining pattern seen in normal uteri. The neoplastic epithelial component in 2 of the 6 uterine carcinomas in TBBPA-exposed animals had focal nuclear immunoreactivity for PR (Figure 2B), whereas in control animals, there was diffuse nuclear immunoreactivity in 6 of the 9 tumors (Figure 2B inset). Poorly differentiated tumors from control and TBBPA-exposed animals had increased nuclear immunoreactivity to KI67 (Figure 2C), whereas well-differentiated tumors had little to no KI67 expression (Figure 2D). Finally, all examined uterine carcinomas (6 of 6) from TBBPA-exposed animals displayed diffuse and moderate to strong nuclear immunoreactivity to antibodies against TP53 (Figure 2E and Table 5). Five of the nine spontaneous uterine carcinomas displayed nuclear immunoreactivity, with an average intensity score of 1.0; 2 displayed strong intensity (3+) and were associated with poor differentiation. Three spontaneous tumors had weak immunoreactivity (1+), and 4 were negative (Figure 2F). The 2 well-differentiated spontaneous uterine carcinomas did not express TP53.
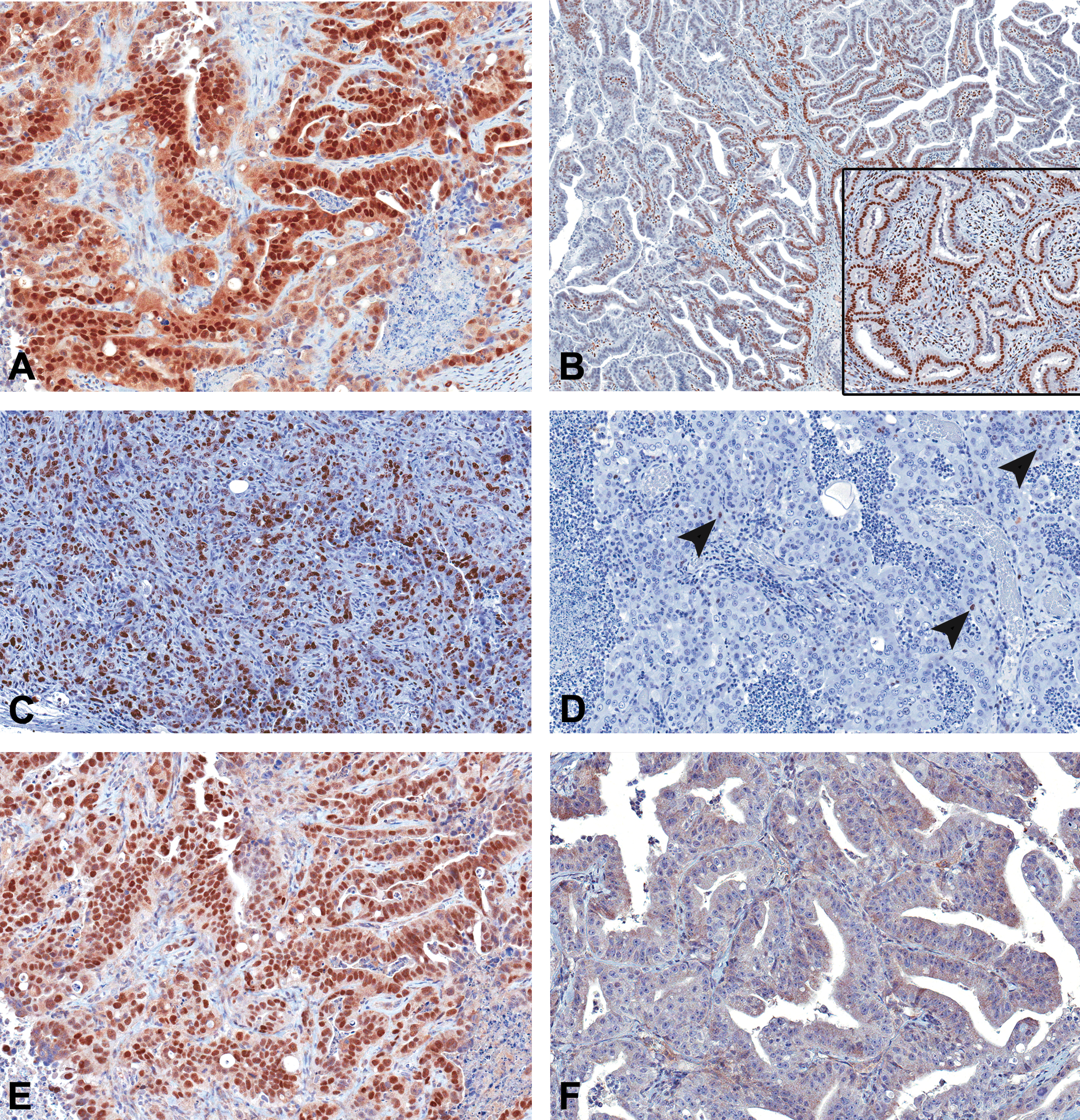
Immunohistochemical features of uterine carcinomas. (A) All spontaneous and tetrabromobisphenol A (TBBPA)-induced uterine adenocarcinomas had diffuse nuclear immunoreactivity to estrogen receptor-alpha (ERα). (B) Two uterine carcinomas from TBBPA-exposed animals had focal nuclear immunoreactivity to progesterone receptor (PR) antibody, and 1 spontaneous tumor showed diffuse nuclear immunoreactivity (inset). (C) Poorly differentiated uterine carcinomas showed diffuse KI67 expression, whereas well-differentiated tumors (D) had single cell (arrowheads) to no expression of KI67. (E) Uterine carcinomas from all TBBPA-exposed animals showed strong nuclear immunoreactivity to antibodies to Tp53, whereas 4/9 spontaneous tumors were negative (F).
Uterine carcinomas in TBBPA-exposed Wistar Han rats had increased incidence of Tp53 mutations compared to spontaneous uterine carcinomas. Since mutations of several cancer genes are associated with clinical outcomes in humans, we screened spontaneous and TBBPA-associated uterine carcinomas for mutations relevant for EC in women. FFPE uterine carcinomas from TBBPA-exposed animals (n = 22) and spontaneous uterine carcinomas from vehicle controls (n = 10) were examined for mutations in Kras, Ctnnb1, and Tp53 (Table 6). In TBBPA-exposed animals, 15 of 22 (68%) tumors harbored mutations in Tp53, often with multiple mutations per tumor (Table 7), whereas in spontaneous uterine carcinomas, 2 of the 10 (20%) tumors had a mutation in this gene (Table 6). Of the 22 uterine carcinomas from TBBPA-exposed animals that were assessed for mutations in Ctnnb1 and Kras, 3 harbored mutations in Ctnnb1, and 1 animal had a Kras mutation (Table 6). Although, neither Ctnnb1 nor Kras mutations were observed in spontaneous uterine carcinomas in this study (0 of 10; Table 6), the incidence in uterine carcinomas from TBBPA-exposed rats was very small, and the relative differences in the number examined (10 spontaneous vs. 22 TBBPA-exposed) relatively large; therefore, this difference is likely not statistically significant and may reflect variations in normal biology of these tumors in this strain.
Mutation frequency of Tp53, Ctnnb1, and Kras in uterine carcinomas from control and TBBPA-exposed WH rats.

Note: Ctnnb1 = catenin (cadherin-associated protein), beta 1; Kras = Kirsten rat sarcoma viral oncogene homolog; TBBPA = tetrabromobisphenol A; Tp53 = tumor protein 53; WH = Wistar Han.
Morphologic and molecular features in uterine carcinomas from humans and TBBPA-exposed WH rats.

Note: CDH1 = E-cadherin; CCND1 = cyclins D1; CTNNB1 = catenin (cadherin-associated protein), beta 1; HER2 = human epidermal growth factor; KRAS = Kirsten rat sarcoma viral oncogene homolog; ND = not determined; P16 = cyclin-dependent kinase inhibitor; PIK3CA = phosphatidylinositol-4,5-bisphosphate 3-kinase, catalytic subunit alpha; PTEN = phosphatase and tensin homolog; TP53 = tumor protein 53.
a Horn et al. (2007), Llobet et al. (2009), Lax (2004), Matias-Guiu et al. (2001), Ryan et al. (2005), Liu (2007).
bCurrent study.
Uterine carcinomas from TBBPA-exposed rats had increased HER2 expression compared to spontaneous uterine carcinomas. Since alterations in gene expression of a variety of growth factors, oncogenes, and tumor suppressor genes are also present in ECs in women, including HER2, CCND1, and CDH1, we evaluated uterine carcinomas for alterations in the expression of these genes (Figure 3 and Table 7). All uterine carcinomas in TBBPA-exposed animals and vehicle control animals overexpressed Her2; expression in uterine carcinomas from TBBPA-exposed animals averaged a 30-fold increase compared to a 2-fold increase in spontaneous carcinomas in vehicle control rats. Approximately 40 to 80% of type II tumors in humans overexpress HER2, as opposed to 10 to 30% of type I EC tumors. All uterine carcinomas examined overexpressed Ccnd1 and Cdh1, and the fold increase in expression was not significantly different between spontaneous uterine carcinomas and uterine carcinomas from TBBPA-exposed rats. In contrast to ECs in women, loss of Pten and P16 tumor suppressor gene expression was not observed in spontaneous or TBBPA-associated carcinomas. In addition, both spontaneous and TBBPA-associated uterine carcinomas had an increased expression of microsatellite markers (Msh2, Mlh1, and Msh3) instead of the characteristic decreased expression associated with microsatellite instability in ECs in women. Additionally, there was no difference in expression of Irf1, a tumor suppressor gene that has been shown to decrease with the grade of endometrial adenocarcinomas in women (Kuroboshi et al. 2003) between TBBPA-induced and spontaneous tumors.

Real-time quantitative PCR (qPCR) gene expression of mediators involved in human endometrial carcinogenesis. Uterine carcinomas in tetrabromobisphenol A (TBBPA)-exposed animals averaged a 30-fold overexpression of human epidermal growth factor 2 (Her2) compared to a 2-fold increase in spontaneous uterine carcinomas. Uterine carcinomas from control and TBBPA-exposed animals overexpressed cyclin D1 (Ccnd1) and E-cadherin (Cdh1), although the fold increase in expression was not significantly different between tumors.
Discussion
Long-term exposure of Wistar Han rats to TBBPA resulted in treatment-related increased frequencies in uterine carcinomas. The majority of uterine carcinomas in both TBBPA-exposed animals and spontaneous tumors were considered poorly differentiated. Uterine carcinomas in this study varied from a relatively more well-differentiated glandular and papillary form to poorly differentiated, infiltrative clusters, ribbons, or solid sheets of anaplastic glandular epithelial cells. A majority of poorly differentiated carcinomas also expressed high levels of KI67, in contrast to well-differentiated tumors. In women, KI67 expression is a prognostic factor correlated with higher grade, more aggressive ECs and a poor prognosis (Lax 2004; Oreskovic et al. 2004; Salvesen et al. 1999; Zannoni et al. 2010). In addition to similar morphologic features, for the most part alterations in gene expression and mutation spectra relevant for human EC were similar in Wistar Han rat uterine carcinomas regardless of TBBPA exposure. Uterine carcinomas in both groups had nuclear expression of ERα, increased expression of Her2, Pten, P16, Irf1, Ccnd1 and Cdh1, and microsatellite markers. Loss of Pten and P16 tumor suppressor gene expression are common events in type I and type II EC in women, respectively, and increased expression of Ccnd1 is a common feature of type II tumors. While EC in women is generally associated with microsatellite instability (loss of Msh2, Mlh1, and Msh3) and loss of Cdh1, P16, and Pten function, both spontaneous and TBBPA-associated uterine carcinomas had increased expression of these mediators.
The primary molecular differences observed in uterine carcinomas from TBBPA-exposed animals compared to spontaneous tumors included a marked increase in Tp53 mutation and increased Her2 gene expression, and possibly a reduction in PR expression. While all uterine carcinomas, irrespective of differentiation or TBBPA exposure, showed nuclear expression of ERα, PR expression was reduced or lost in a majority of uterine carcinomas from TBBPA-exposed animals and fewer spontaneous tumors. Expression of ERα and loss of PR expression in TBBPA-exposed Wistar Han rats may suggest that these tumors remain estrogen responsive, but have lost receptor functionality for progesterone. Interestingly, research has shown that PR status may be a more reliable prognostic factor than ERα status in EC in women and corresponds with a more aggressive, high-grade tumor (Oreskovic et al. 2004; Zannoni et al. 2010).
Spontaneous uterine carcinomas and carcinomas from TBBPA-exposed rats both shared morphologic and molecular features with type I and II EC in women. Features common to both spontaneous and TBBPA-exposed tumors that were shared with type I EC tumors in women included late onset, endometrioid differentiation, and ERα hormone receptor expression. Features shared between rat uterine carcinomas and human type II EC include poor differentiation and deep invasion, high mitotic rate and KI67 index, increased cellular pleomorphism and atypia, Her2 expression, and Tp53 mutation. While both spontaneous uterine carcinomas and those from TBBPA-exposed rats share features of type I and type II human EC, carcinomas from TBBPA-exposed rats have significant differences from spontaneous tumors. In addition to the abovementioned features, uterine carcinomas from TBBPA-exposed rats had significantly increased Tp53 mutation and Her2 expression compared to spontaneous tumors, features more often observed in type II EC in women. Taken together, the morphologic and molecular features of uterine carcinomas in TBBPA-exposed rats overlap those of type I and II EC in women, most consistent with high-grade type I EC (Table 7), suggesting that uterine carcinomas in TBBPA-exposed rats may closely resemble this type of EC. However, many human EC tumors also share features of both subtypes, and a clear distinction based on histomorphologic and molecular features is sometimes difficult.
By far, the most striking differences between uterine carcinomas in TBBPA-exposed animals and controls were the marked increase in Her2 gene expression and Tp53 mutation, 2 of the major genetic alterations in type II EC in women (Liu 2007). Her2 or Her2/neu is a receptor tyrosine kinase that is an upstream regulator of a number of important cell proliferation and anti-apoptosis pathways. It is highly upregulated in type II EC and has been linked to adverse prognostic outcome and decreased overall survival (Doll et al. 2008). The TP53 tumor suppressor gene is responsible for cell cycle checkpoint maintenance, apoptosis, and genomic stability (Blagosklonny 2000). Loss of TP53 tumor suppressor function is important in a wide variety of human cancers, including the development of aggressive endometrial cancers. TP53 mutation is associated with high-grade tumors, advanced disease, and a poor prognosis (Liu 2007). TP53 mutations are extremely common in type II EC (80–90%) and found in a subset (10–20%) of high-grade type I EC in women (Liu 2007; Llobet et al. 2009; Oreskovic et al. 2004; Zannoni et al. 2010); therefore, it is thought to possibly occur as a late event in the development of aggressive endometrial cancer. In this study, the high rate of Tp53 mutation in uterine carcinomas from TBBPA-exposed rats compared to spontaneous tumors suggests that the increased incidence of uterine carcinomas in treated animals may be driven through a Tp53-dependent mechanism. While some degree of TP53 protein expression was observed in both spontaneous and TBBPA-associated uterine carcinomas, those tumors in TBBPA-exposed animals showed a much higher degree of immunoreactivity, consistent with the increase in mutation rate. The presence of nuclear immunoreactivity did not always correspond with the presence of a mutation in Tp53 in this series of samples. However, these types of discordant results have been reported previously and may be a result of gain of function of the TP53 gene, accumulation of wild type protein due to DNA damage (Blagosklonny 2000), or a reflection of a low percentage of mutation-positive tumor cells (Bian et al. 2001; George et al. 2007; Kropveld et al. 1996). This latter explanation could also account for nuclear expression of TP53 in spontaneous samples, which was much lower than that of the TBBPA samples, in which there was a high rate of mutation. Alternatively, gene mutations may have occurred in unsequenced exons.
Other features of type I and II EC in women include Ctnnb1 and Kras mutation. Beta-catenin is the protein product of Ctnnb1 and is an essential part of the WNT signaling pathway, regulating cellular processes including proliferation, motility, and survival (Wu et al. 2007). CTNNB1 mutation in EC in women occurs in approximately 20 to 40% of type I tumors compared to 0 to 5% in type II tumors (Llobet et al. 2009). KRAS mutation, seen in 10 to 30% of type I EC in women (Llobet et al. 2009), is associated with constitutive activation, enhanced cellular proliferation, transformation, and cell survival, and has been associated with adverse clinical outcomes in women (Matias-Guiu et al. 2001). Ctnnb1 and Kras mutations were not observed to play a significant role in either spontaneous uterine carcinomas or those from TBBPA-exposed animals in this study; the general lack of these mutations suggests that these are not critical pathways for the generation of these tumors in the Wistar Han rat model.
The results of this study suggest that uterine carcinomas in TBBPA-exposed Wistar Han rats most closely resemble high-grade type I EC in women on the molecular level, which are typified in humans by conservation of ERα expression, a high rate of TP53 mutation, more aggressive clinical course, and a poor prognosis compared to other typical low-grade type I tumors (Liu 2007; McConechy et al. 2012; Zannoni et al. 2010). Interestingly, the Berlin-Druckrey II (BDII) Han rat model develops a high rate of spontaneous uterine carcinomas (∼90%), which, based on molecular features, have also been classified as similar to high-grade type I ECs in women (Adamovic et al. 2008; Behboudi et al. 2001; Nordlander et al. 2005; Samuelson et al. 2009). They have an early onset, endometrioid differentiation, and are estrogen responsive, but have high rates of Tp53 mutation, similar to the uterine carcinomas observed in TBBPA-exposed Wistar Han rats in this study. However, carcinomas in TBBPA-exposed rats have additional features of type II ECs, including overexpression of Her2.
In summary, uterine carcinomas were observed with a higher frequency in Wistar Han rats exposed to TBBPA, have higher rates of proliferation, an increased rate of Tp53 mutation, a tendency toward decreased PR expression, and increased HER2 expression compared to spontaneous uterine carcinomas in Wistar Han rats. These features overlap with those of type I and II EC in women and are most concordant with high-grade type I tumors, which are clinically more aggressive and harbor a poorer prognosis compared to other type I tumors. Furthermore, the presence of such a high rate of Tp53 mutations in TBBPA-exposed animals compared to controls suggests that uterine carcinogenesis in Wistar Han rats exposed to TBBPA is partially Tp53-dependent. Since alteration of TP53 is relevant for aggressive endometrial cancer in women, and plays a predominant role in the pathogenesis of a number of other cancers in humans, it is reasonable to hypothesize that TBBPA exposures may pose a cancer risk to humans through similar mechanisms. However, inherent species differences in the metabolism of TBBPA or sensitivity to its biologic effects on uterine pathophysiology may exist, and further functional studies on the carcinogenic effects of TBBPA on the female reproductive tract are warranted.
Footnotes
Acknowledgments
The authors would like to thank the CMPB Necropsy Core Laboratory personnel, Histology, Immunohistochemistry, and Sequencing Core Laboratories for their technical expertise; and Ms. Beth Mahler and Ms. Eli Ney (NIEHS) for images and figures.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institutes of Environmental Health Sciences (NIEHS), National Institutes of Health (NIH), and the Division of the National Toxicology Program (DNTP).
Author Contribution
Authors substantially contributed to conception or design (AP, MH), contributed to acquisition (JH, TO, H-HH, SB, T-VT, AP, TM, SP, MH), contributed to analysis (JH, TO, H-HH, SB, T-VT, AP, TM, JD, SP, SEMH), contributed to interpretation (JH, TO, AP, SP, SE, MH), and drafted the manuscript (JH, TO, AP, SP, MH). All authors critically revised the manuscript, gave final approval, and agreed to be accountable for all aspects of the work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Authors’ Note
Janice B. Harvey and Tanasa S. Osborne contributed equally to this work. This article may be the work product of an employee or group of employees of the National Institute of Environmental Health Sciences (NIEHS), National Institutes of Health (NIH); however, the statements, opinions, or conclusions contained therein do not necessarily represent the statements, opinions, or conclusions of the NIEHS, NIH, or the U.S. government.