
Abstract
Researchers have sought to understand the specialized processing of faces and bodies in isolation, but recently they have considered how face and body information interact within the context of the whole body. Although studies suggest that face and body information can be integrated, it remains an open question whether this integration is obligatory and whether contributions of face and body information are symmetrical. In a selective attention task with whole-body stimuli, we focused attention on either the face or body and tested whether variation in the irrelevant part could be ignored. We manipulated orientation to determine the extent to which inversion disrupted obligatory face and body processing. Obligatory processing was evidenced as performance changes in discrimination that depended on stimulus orientation when the irrelevant region varied. For upright but not inverted face discrimination, participants could not ignore body posture variation, even when it was not diagnostic to the task. However, participants could ignore face variation for upright body posture discrimination but not for inverted posture discrimination. The extent to which face and body information necessarily influence each other in whole-body contexts appears to depend on both domain-general attentional and face- or body-specific holistic processing mechanisms.
Keywords
Introduction
Information from both faces and bodies are important for social cognition. Both help us understand who people are, what they are feeling, and what they are doing so that we can anticipate what they are going to do next (Reed & McIntosh, 2008; Slaughter, Stone, & Reed, 2004). The social relevance of faces has been extensively documented showing specialized cognitive and neural mechanisms for processing isolated faces (e.g., Maurer, Le Grand, & Mondloch, 2002; Farah, Levinson, & Klein, 1995; Kanwisher, McDermott, & Chun, 1997). Specialized processing has also been found for isolated bodies (e.g., Minnebusch, Suchan, & Daum, 2009; Reed, Stone, Bozova, & Tanaka, 2003; Reed, Stone, Grubb, & McGoldrick, 2006, Robbins & Coltheart, 2012a, 2012b; Yovel, Pelc, & Lubetzky, 2010). In everyday life, however, we see and interact with people who have heads attached to their bodies. To understand this whole-person perception, researchers have turned to the question of how faces and bodies might be processed together and how their perception can influence one another (e.g., App, Reed, & McIntosh, 2012; Aviezer, Dudarev, Bentin, & Hassin, 2011; Aviezer et al., 2008, Aviezer, Trope, & Todorov, 2012a, 2012b; Bernstein, Oron Sadeh, & Yovel, 2014; Kaiser, Strand, Seidl, Kastner, & Peelen, 2014; Rice, Phillips, Natu, An, & O’Toole, 2013; Simhi & Yovel, 2016, 2017). In this study, we investigate whether face-body integration is obligatory and whether the contributions of face and body information are symmetrical.
Selectivity of Isolated Faces and Bodies
Researchers have studied the perception of faces and bodies extensively using paradigms designed to isolate processing to either faces or bodies. In behavioral studies of isolated face and body perception, evidence suggests that both face and body perception rely on specialized cognitive mechanisms such as holistic and configural processing (Collishaw & Hole, 2000; Farah, Tanaka, & Drain, 1995; Harris, Vyas, & Reed, 2016; Reed, McGoldrick, & Bukach, 2016; Reed, Stone, et al., 2006; Seitz, 2002). Both are susceptible to disruption of processing when images are inverted (e.g., faces: Farah et al., 1995; Yin, 1969; bodies: Reed et al., 2003; Reed, Stone, et al., 2006). Brain imaging studies of isolated faces and bodies have shown substantial evidence for selectivity of faces and bodies (Downing, Chan, Peelen, Dodds, & Kanwisher, 2006; Downing, Liu, & Kanwisher, 2001; Haxby & Gobbini, 2011; Kanwisher & Yovel, 2006; Peelen & Downing, 2005, 2007). For example, Peelen and Downing (2005) directly contrasted isolated face perception to isolated body perception identified face selective activation in the right occipital face area and a body-selective region bilaterally in the extrastriate cortex. Although there was overlapping activation in the right fusiform gyrus (FG), a direct contrast of faces and bodies revealed distinct peaks of activation for faces and bodies. Moreover, although face and body deficits can co-occur (Moro et al., 2012), selective deficits have been documented for both face perception (e.g., Farah, Levinson, et al., 1995; Kanwisher et al., 1997; Maurer et al., 2002) and body perception (Moro et al., 2012; see Haggard & Wolpert, 2005 for a review). Taken together, these studies suggest that although faces and bodies may rely on similar cognitive processing styles, they are represented, at least in part, by separate neural substrates.
Much has been learned about these two domains in isolation, but this approach has some important limitations. To isolate face-specific processing studies have often removed all alternative sources of variation typically available during person perception. Face studies often remove or hold constant the body, contour of the face, hair, and skin color so that only facial features are varied (see Bukach & Peissig, 2009 for review). Similarly, studies of body processing have reduced facial information so it is nondiagnostic for body posture recognition: Facial features in the whole-body stimuli have been rendered indistinct and unvarying across stimuli (Reed et al., 2003; Reed, Stone, et al., 2006), obstructed from view (Morris, Pelphrey, & McCarthy, 2006), or masked (Robbins & Coltheart, 2012b; Soria-Bauser, Suchan, & Daum, 2011; Stekelenburg & de Gelder, 2004; Yovel et al., 2010). Importantly, studies that present faces and bodies in isolation or hold one part constant cannot address the critical questions of whether and how information from face and body may interact during whole-person perception, under circumstances that include types of variability encountered in real-world tasks.
Integration of Face and Body in Whole-Body Studies
More recently researchers investigated how faces and bodies are processed together by studying them in the context of whole-body stimuli. The importance of using whole-body stimuli is evident in both behavioral and neural studies. These studies provide evidence that face and body perception may be integrated in some parts of the perceptual cognitive processing stream in a way that is not captured by isolated face and body studies. Both efficiency scores and N170 amplitude and latency measures reveal a strong body-inversion effect for whole-body stimuli with masked faces but not for headless bodies (Minnebusch et al., 2009). Further, several functional neuroimaging studies comparing whole body to isolated face and body stimuli have found differential neural activation for whole bodies (e.g., Berstein, Oron Sadeh, & Yovel, 2014; Brandman & Yovel, 2010; Harry, Umla-Runge, Lawrence, Graham, & Downing, 2016), indicating that faces and bodies may have overlapping but differential representations in face-selective areas such as the FG and anterior temporal lobe (ATL). In the FG, whole-body activation could be modeled by a linear combination of isolated face and body activation, suggesting the representations of faces and bodies are additive and may therefore be independent (Berstein et al., 2014). In contrast, faces and whole bodies appear to have shared neural coding in the right ATLs such that whole body and face responses were positively correlated on a voxel-by-voxel basis in the right ATL but not body-part and face responses (Harry et al., 2016). This evidence suggests that face and body information may be integrated in some neural regions but not others.
Evidence for integration of face and body information has also been found in behavioral tasks that require whole-body judgments. When body posture varies in meaningful, dynamic, and emotionally relevant configurations and the overall impressions of emotional intensity are evaluated, face and body information complement and interact with each other (e.g., App et al., 2012; Aviezer et al., 2008; Aviezer et al., 2011; Aviezer et al., 2012a, 2012b; Burton, Wilson, Cowan, & Bruce, 1999; Meeren, van Heijnsbergen, & deGelder, 2005; Rice et al., 2013; Roark, O’Toole, Abdi, & Barrett, 2006; Van den Stock & de Gelder, 2012, 2014; Simhi & Yovel, 2016, 2017). In many of these studies, whole-body stimuli are constructed so that the face and body configurations represent either congruent emotions (e.g., both face and body convey anger) or incongruent emotions (e.g., face conveys anger, and body conveys fear). Ratings of emotion intensity for the whole-body stimuli are reduced or affected in incongruent conditions compared with congruent conditions, indicating that information from the face and body is integrated for emotional processing. Judgments of whole-body person identification also show that face and body information can be combined to improve recognition over faces alone, particularly when the stimulus pairs are from different instances and are difficult to recognize from face information alone (Rice et al., 2013).
The nature of face-body integration during whole-person judgments and the relative salience of face and body information appears to depend on a number of factors, including task demands and stimulus properties that have an impact on the way that attention is distributed across the stimulus. For example, evidence suggests that different goals or task demands can allocate attention differentially; faces contribute more to whole-body emotion judgments, but that bodies contribute more to judgments of motion direction (App et al., 2012). Although it is true that when people are viewed up close, person recognition is based more on the face, bodies also contribute to this recognition process when people are viewed from a distance in which the whole person can be viewed at once (Hahn, O’Toole, & Phillips, 2016). Other studies have found that body information aided person recognition only after exposure to motion (Simhi & Yovel, 2016). Variability in body posture may be important to draw attention to the body during whole-body perception.
Thus, from the earlier studies, we know that faces and bodies can be processed together when viewers are provided with whole-body stimuli and the nature of the task predisposes processing to integrate information from both face and body. However, these studies do not directly address whether this coprocessing is obligatory in both face-body and body-face directions.
Obligatory Processing in Selective Attention Studies
One way that obligatory processing has been investigated is through selective attention paradigms that require participants to base their judgment on a single part of an object and ignore irrelevant information from the rest of the object. In these paradigms, integration is operationally defined as a failure of selective attention or the extent to which subjects cannot successfully ignore the irrelevant information (in this case, face in body context or body in face context). Thus, if the parts are processed in an integrated fashion, the irrelevant part should interfere with judgments of the target part. This established effect provides an index of obligatory processing of the irrelevant part and dependence between object parts. For example, Aviezer et al. (2011) instructed participants to focus on facial expressions and avoid using other contextual information from the body. Body configuration influenced the categorization of facial emotion with and without working memory load, suggesting that bodies automatically influence face processing. However, Aviezer et al. only focused on how the emotional body context affected facial emotion judgments. Importantly, they did not examine how face information might affect body processing for nonaffective face and body expression stimuli and for nonemotional, perceptual judgments.
Another selective attention paradigm commonly used in both face and body studies is the composite task. In the composite task, whole-body stimuli are divided in half, and participants decide whether the cued half of two images match (i.e., belong to the same object), while ignoring the noncued half of the stimulus pair. Holistic processing is indicated by the composite effect, that is, impaired performance for trials in which one half matches and the other half does not because the participant cannot selectively attend to a single part. This interference is reduced when images are misaligned (i.e., the edges of the tops and bottoms of objects are not matched) or inverted, manipulations designed to disrupt expert mechanisms such as holistic processing. By including both face and body target trials, the paradigm can address the relative salience of face and body information in the integrated representation.
However, composite whole-body tasks have yielded mixed results as to whether the body modulates face processing in whole-body stimuli. These conflicting results may be attributed to the presence or absence of variability in the whole-body postures: When posture varied or the body was divided via the vertical body axis, the composite effect was found indicating holistic body processing (Robbins & Coltheart, 2012b; Willems, Vrancken, Germeys, & Verfaillie, 2014); however, when posture was not varied and the body was divided along the horizontal axis, no composite effect was found (Soria-Bauser et al., 2011).
The importance of posture variability may be related to the same mechanisms underlying the role of motion in integrating face and body information. One possibility is that variability and motion both influence allocation of attention. Some research suggests that faces automatically draw attention even when they are competing with other objects (Palermo & Rhodes, 2007; Langton, Law, Burton, & Schweinberger, 2008; Ro, Friggel, & Lavie, 2007; Lavie, Ro, & Russell, 2003). Further, faces appear to capture attention even in whole-body stimuli, particularly for upright stimuli (e.g., Arizpe, McKean, Tsao, & Chan, 2017; Rice et al., 2013). However, eye-tracking studies have shown that despite a tendency to fixate on faces for upright whole-body stimuli, fixations move more centrally for inverted bodies. Thus, variability may be particularly important to show the influence of body information on face judgments as well as the influence of faces in body posture judgments even in nonstandard orientations (i.e., inversion). For example, variation of facial features could modulate the interaction between face and body processing most strongly in conditions where attention is not already drawn to the face (i.e., for inverted faces). Because real-world, whole-body perception includes variability in both face identity and body posture, it is important to study person perception using whole-body stimuli in experimental designs where information from both faces and bodies vary.
Current Study
Taken together, the literature earlier suggests that the degree to which the face and body are processed together may be related to how much they are allowed to vary within the context of the whole body. We know that face and body information can interact but we do not know if this integration is obligatory, especially when the irrelevant information is not diagnostic to task performance. What is missing from prior studies is a systematic manipulation of variation in both face and body parts that provides a more ecological, whole-body context to judge the relative salience of face and body information. In this study, we investigate the extent to which face and body posture processing is obligatory in whole-person perception and the relative salience of face and body information by comparing conditions in which faces and body posture are free to vary versus when they are held constant. By systematically manipulating variation in face and body in a selective attention task, we can assess both the influence of the face on body judgments, as well as the influence of the body on face judgments, and whether these effects are symmetrical.
In the real world, faces and bodies provide information relevant to a variety of social goals, including perception of identity, expression, and action intention. In our task, we do not specify a particular goal beyond determining whether the target parts are the same or different. This task is therefore a strong test of obligatory processing as the face and body posture information are likely differentially diagnostic for particular social goals. Further, we also include an inversion manipulation that is often used for the perception of faces and bodies and has been shown to disrupt a variety of orientation-specific processing mechanisms, including holistic, relational, and feature processing (McKone & Yovel, 2009; Rossion, 2009; Yovel, 2009). The current study provides a novel and strong test of the hypothesis that variability is an important factor that influences whole-body perception, thereby helping to explain conflicting results in the literature and providing better understanding of the attentional and perceptual processing involved in person perception.
Experiment 1: Selective Attention to the Face in Whole-Body Stimuli
In Experiment 1, we explore the obligatory processing of body posture information when participants are directed to selectively attend to the face. Specifically, we manipulated the variability of the nonfocal, irrelevant body region to determine if faces can be processed independently of the rest of the body (i.e., body posture). Using a sequential matching paradigm for whole-body stimuli, we assessed upright and inverted face discrimination under two variability conditions: A constant condition in which the body posture was the same for each face trial comparison (e.g., body posture of Stimulus 1 matched the body posture of Stimulus 2), and a varied condition in which body postures never matched for successive stimuli. To the extent that body posture variability directs attention to the whole body, task performance for faces should be reduced when body posture varies compared with when body posture is held constant. To the extent that this integration of face and body rely on orientation-specific mechanisms, the effect of body posture variation should be greater for upright than inverted trials.
Method
Participants
Fifty-one participants (64% female; mean age = 19.69, SD = 1.07) volunteered for partial course credit. In this study, we equated sample sizes across all three experiments, and different participants were used in each experiment. We used slightly larger sample sizes than is often used in the literature because we manipulated stimulus variability, a factor known to influence effect size. We wanted to ensure we had enough power to detect potential bidirectional effects of face variation on body posture discrimination and vice versa and not make erroneously conclusions about potential null effects.
Because three participants’ performance was at chance in at least two conditions, their data were excluded from analyses, such that the data from 48 participants were included in the reported analyses.
Stimuli
For use as experimental and practice stimuli in all three experiments, a large set of 204 whole-body stimuli were constructed by combining body postures and faces that were previously used in Reed, Stone, et al. (2006). Body postures were three-dimensional, black-and-white figures created with Poser 2.0 (Curious Labs, Santa Cruz, CA). Each posture was a physically possible, novel, nonmeaningful pose that was visually distinguishable from the others and could not be easily labeled. The poses were asymmetrical with respect to both vertical and horizontal axes. Distractors for each posture were constructed by altering the position or angle of the arms, legs, and head angle of the original stimulus.
The faces were male, black-and-white images created using FACES 3.0 (Thomas Investigative Publications, Austin, TX) software. All faces shared the same facial outline (hair hairline, ears, and chin) so that only the internal features (eyes, nose, and mouth) differed between faces. Face distractors were constructed by altering all three internal features.
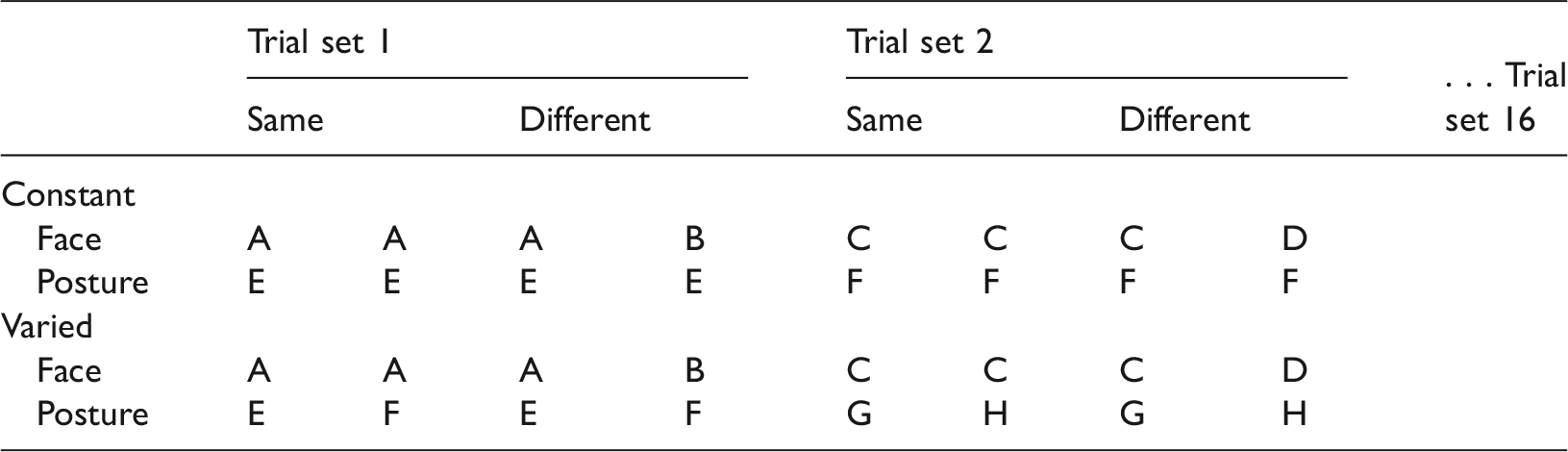
Using Photoshop CS3 (Adobe Systems, Inc., San Jose, CA), the whole-body stimuli were created by aligning the bodies and the faces and adjusting their relative size so that they fit proportionately to create 8 cm × 8 cm upright orientation stimuli (Figure 2). From this set of whole-body stimuli, different sets and pairs of stimuli were assembled by recombining faces and body postures to meet the condition criteria for each experiment (Figure 1). Inverted versions of all the stimuli were created by rotating the images 180°. Pilot testing confirmed that faces and body postures in the whole-body stimuli were highly discriminable, although body postures were more discriminable than face stimuli in the whole-body context because of their relative size and spatial frequency of their features.
Example of stimulus pairs: (top row) body discrimination with a constant face, (middle row) body discrimination with a constant body posture, and (bottom row) face or body posture discrimination with varied face or body posture. Sequential matching paradigm: two stimuli with the same orientation are judged to be same or different based on either their focal regions—either their faces or their body postures. An inverted trial is depicted.

Experiment 1—Face Discrimination Task: Whole-Body Stimulus Construction Across Trials and Conditions.
Procedure
Participants completed a sequential matching task in which they discriminated the faces of whole-body stimuli. Stimulus 1 was presented for 250 ms, followed by blank screen for 1000 ms and a second stimulus that remained on the screen until response; a central fixation cross was presented for a 1000 ms inter-trial interval (ITI) (Figure 2). Using their index fingers, participants pressed the S-labeled key (either the “s” or the “l” key on the keyboard) to indicate that the two stimuli were the same, or the D-labeled key (either the “s” or the “l” key) to indicate that the stimuli were different; key assignment was counterbalanced. Participants were instructed to respond as fast and accurately as possible.
To maximize the power of the manipulations, we used a category-block design in which experimental trials were blocked by the variability of the nonfocal region (constant and varied) first and then by orientation (upright and inverted), yielding four blocks in total. Before each block of trials, participants received instructions and four practice trials that included an example of each condition using practice-only stimuli. Block order was counterbalanced across participants such that constant and varied blocks were counterbalanced and then upright and inverted blocks were counterbalanced within the constant and varied blocks. Within each block of 32 trials, half of the trials were same, and half were different. There was a total of 128 experimental trials. Response time (RT) and accuracy were recorded.
Results and Discussion
For each participant, we calculated visual sensitivity (d-prime, d′: z(hit rate) – z(false alarm rate) with correction for hit and false alarm rates of 0 or 1 to eliminate infinite z values [Macmillan & Creelman, 1991]) and mean RTs for correct trials for each condition. Trials outside of a RT window of 150 ms to 3000 ms were excluded from analyses as “jump the gun” and inattention errors (less than 1% of trials). Block order produced no main effects or interactions (p > .05) for either measure so it was not considered further.
Sensitivity and RT data were submitted to a 2 × 2 repeated measures analysis of variance (ANOVA) with nonfocal region variability (constant and varied) and orientation (upright and inverted) as within-subjects factors. Sensitivity data are illustrated in Figure 3(a). Body posture variation affects face discrimination, indicating that face processing is not independent of body posture processing. When body posture varied, face discrimination performance declined and the effect was larger for upright than inverted trials. Results indicate a main effect of variability with superior face discrimination when body posture was held constant (constant = 2.45 [SD = .14] vs. varied = 1.98 [SD = .14]; F(1, 47) = 9.61, p = .003, ŋp2 = .17) and a main effect of orientation with superior face discrimination for upright trials (upright = 2.58 [SD = .15] vs. inverted = 1.85 [SD = .15]; F(1, 47) = 19.17, p < .0001, ŋp2 = .29). The Variability × Orientation interaction was significant, F(1, 47) = 4.15, p < .05, ŋp2 = .08. Simple effects analysis revealed that body variability affected upright face discrimination, F(1, 47) = 10.74, p = .002, ŋp2 = .19, but not the inverted condition, F(1, 47) < 1, p = .42, ŋp2 = .01. RTs did not indicate speed-accuracy trade-offs and showed the same pattern as the d′ results reported earlier. Constant conditions produced significantly faster responses than varied conditions (940 ms vs. 884 ms; F(1, 47) = 24.69, p < .0001, ŋp2 = 0.34), upright conditions produced faster responses than inverted conditions (878 ms vs. 845 ms; F(1, 47) = 12.95, p < .0001, ŋp2 = 0.22). Follow-up simple effects analyses on the marginal interaction, F(1, 47) = 3.06, p = .09, ŋp2 = .06, showed that variability affected the upright condition (844 ms vs. 915 ms; F(1, 47) = 14.38, p < .0001, ŋp2 = 0.23) but not the inverted condition (926 ms vs. 953 ms; F(1, 47) = 3.56, p > .07, ŋp2 = 0.07).
Results from Experiments 1 and 2. Discrimination sensitivity for face (Experiment 1) and body discrimination (Experiment 2); error bars represent standard error. For constant conditions, the nonfocal region was the same for both stimuli in the trial. For varied conditions, the nonfocal region was never the same.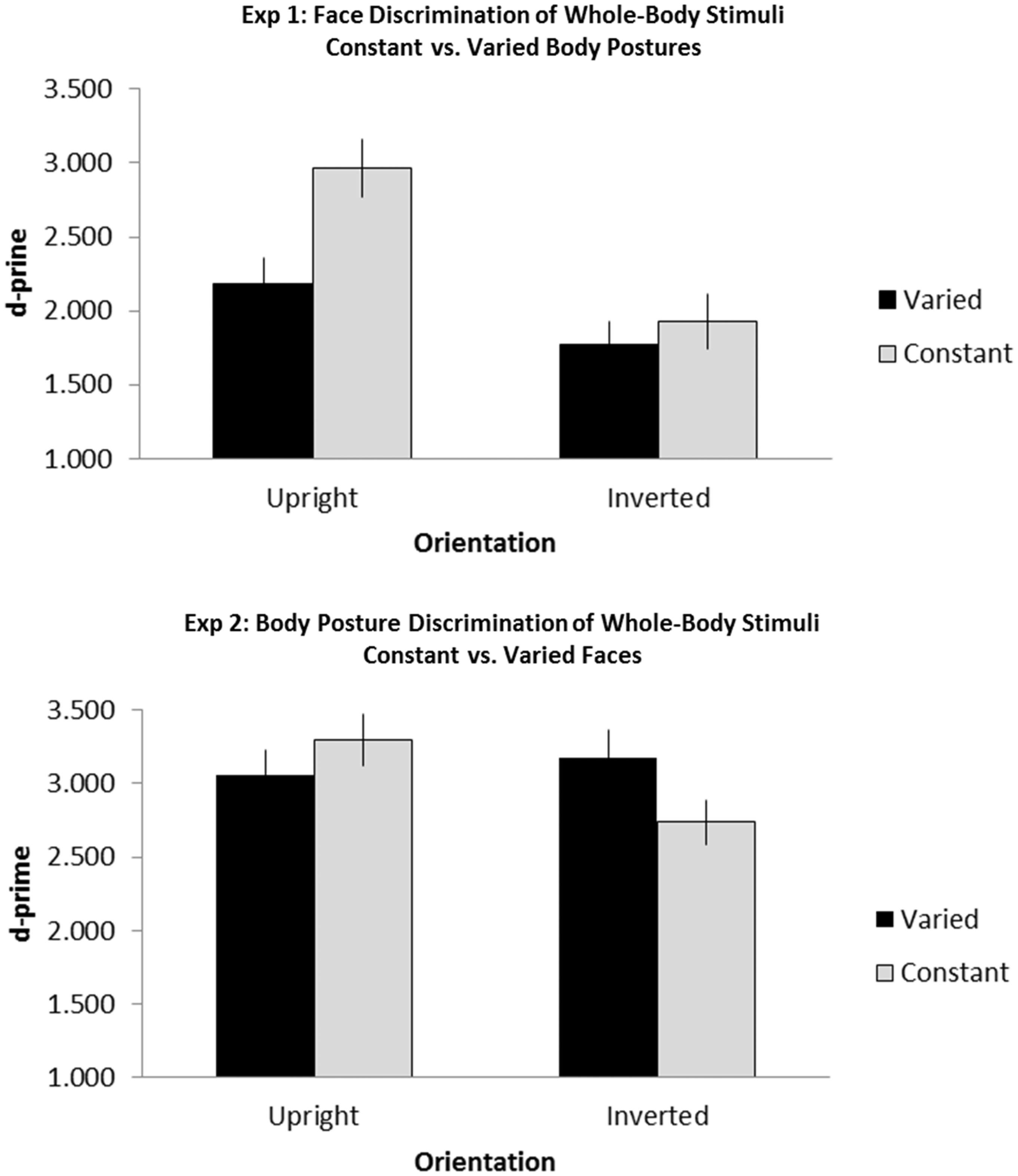
In sum, variation of body postures modulates upright face processing at a viewing distance at which one can see the whole body. Visual sensitivity of upright bodies was reduced when body postures varied, suggesting that changes in body posture information cannot be ignored even when attempting to selectively attend to face information. Further, this interference from variation in body posture was greater for upright than inverted conditions, suggesting that integration of face and body may rely on orientation-specific mechanisms, similar to other specialized processes identified in isolated face and body studies. This finding provides strong support that when body posture is free to vary, as it does in the real world, attention is focused on the whole person and face information is not processed independently of body information.
Experiment 2: Selective Attention to Body Posture in Whole-Body Stimuli
In Experiment 2, we explore how variability of faces affects body posture discrimination. Although research has shown faces enjoy attentional prioritization despite other competing stimuli (Langton et al., 2008; Palermo & Rhodes, 2007; Ro et al., 2007), eye-tracking studies suggest that this automatic attentional capture for faces is greater for upright than inverted whole bodies (Arizpe et al., 2017). Thus, the effect of face variability on body posture discrimination may depend on orientation. We hypothesize that variability has the greatest potential to have an impact when attention is not automatic. That is, if attention is already on the face, then adding variability in the postures should not change performance. However, when attention is not automatically on the face when stimuli are inverted, as Arizpe et al.’s findings suggest, then adding variability might change the way attention is distributed and variability should affect inverted more than upright stimuli in the body posture task.
If obligatory attention to the face is orientation dependent, we may find greater effects of face variability on posture discrimination for upright orientations, but if faces capture attention regardless of orientation, we may find influences of face variation for inverted orientations as well. We assessed upright and inverted body posture discrimination under two variability conditions: A constant condition in which the face was the same for each body posture trial comparison (e.g., face of Stimulus 1 matched the face of Stimulus 2), and a varied condition in which faces never matched for successive stimuli. This paradigm allowed us to isolate the effects of differential variability of faces on body posture processing.
Method
Participants
Fifty-six participants (60% female; mean age = 19.54 years [SD = 1.40]) volunteered for partial course credit.
Stimuli and Procedure
Similar stimuli and procedures were used as in Experiment 1, with two exceptions. First, the task was to discriminate the body postures of whole-body stimuli, with faces varying as the irrelevant part. Second, stimuli were selected such that 16 body postures and 16 distractor postures were paired in the same manner with irrelevant faces.
Results and Discussion
Data were calculated as in Experiment 1. A Variability (constant and varied) × Stimulus Orientation (upright and inverted) repeated-measure ANOVA was conducted. Sensitivity data are illustrated in Figure 3(b). Body posture processing is not independent of face processing, but the nature of their interaction is different in the body task than it was in the face task. Contrary to the face discrimination results, face variability had no effect on upright body posture discrimination but increased visual sensitivity for inverted stimuli. Statistical analyses indicate a significant interaction between variability and orientation, F(1, 55) = 5.20, p = .03, ŋp2 = .09. Simple effects confirmed that face variability did not affect the discrimination of upright body postures, F(1, 55) = 1.11, p = .30, ŋp2 = .02, but it improved the discrimination of inverted body postures, F(1, 55) = 5.46, p = .02, ŋp2 = .09. No main effects of variability (constant = 3.02 [SD = .12] vs. varied = 3.11 [SD = .15]; F(1, 55) < 1, p = .80, ŋp2 = .01) or orientation (upright = 3.18 [SD = .13] vs. inverted = 2.95 [SD = .15]; F(1, 55) = 2.06, p = .16, ŋp2 = .04) were found. RT data indicated no significant main effects or interactions (all ps > .12).
In sum, variation of faces modulated body posture discrimination, even though the faces were less discriminable than the postures. Surprisingly, face variation did not influence the discrimination of upright body posture but increased discrimination of inverted body posture. This data pattern may occur, in part, because of differences in where attention is directed for upright and inverted bodies during body posture discrimination. Arizpe et al. (2017) found that participant fixations were distributed across both head and body of upright whole bodies during body posture matching in which faces were masked, but fixations shifted away from the head toward the pelvis area when bodies were inverted. Therefore, varying face information may not influence upright body posture discrimination because attention is already directed to the face in upright whole bodies, and thus face and body information may be integrated during upright body posture discrimination in both constant and varied conditions. In contrast, face variation may influence inverted trials because in constant conditions, we suggest that attention is drawn toward the pelvis and varying face information draws attention away from the pelvis area more to the upper body. This attention shift results in improved discrimination because the upper body is more diagnostic than the lower body, as demonstrated by increased sensitivity when attention is explicitly directed to the upper body region (Arizpe et al., 2017; Reed, Stone, et al., 2006).
Experiment 3: Within-Participant Face and Body Discrimination
Experiment 3 replicated findings of Experiments 1 and 2 with a within-subject design and extended the visual variation differences between constant and varied conditions. In the constant condition of Experiments 1 and 2, the irrelevant part was held constant within trial but varied between trials. In Experiment 3, we designed the constant condition to be more similar to prior studies that have kept faces or bodies completely constant both within and across trials (e.g., Reed et al., 2003; Reed, Stone, et al., 2006). That is, a single body posture was used for all trials in the constant condition of face discrimination blocks, and a single face stimulus was used for all trials in the constant condition of body posture discrimination blocks.
Method
Participants
Fifty-three participants (62% female; mean age = 19.22 years [SD = 1.43]) with normal or corrected-to-normal vision completed the experiment for partial course credit.
Procedure
Similar stimuli and procedure were used as in Experiment 1, with two exceptions. First, participants performed blocks of both face discrimination and body posture discrimination trials. Second, the constant conditions used stimuli that had a single, stock face or body posture for the nonfocal part of the stimulus; in other words, there was no variation on the nonfocal region even across trials. A single face was selected at random from the posture stimulus pool to be paired with the all faces for the face discrimination task, and a single posture was selected at random form the face pool to be paired with all the postures for the posture discrimination task. Again, for each task, half of the trials were the same, and half were different. For the varied conditions, the focal part of the stimulus differed and nonfocal, irrelevant part of the stimulus was never replicated within and across trials; that is, there was complete variation on the nonfocal region.
To reduce participant confusion about which part (face or body) was relevant on a given trial, we used an ABBA design in which experimental trials were blocked by body region (face and body), the variability of the nonfocal region (constant and varied), and orientation (upright and inverted). Each participant performed eight blocks of 64 trials, for a total of 512 experimental trials.
Results and Discussion
Data were calculated as previous experiments. The sensitivity data illustrated in Figure 4 confirm again that face and body processing are not independent of each other. The omnibus 2 (Task: face and body posture) × 2 (Variability: constant and varied) × 2 (Orientation: upright and inverted) repeated-measures ANOVA was conducted first, followed by separate ANOVAs for each task so that the results of Experiments 1 and 2 could be directly compared with Experiment 3. Significant findings from the overall ANOVA indicated first that faces were less discriminable overall than body postures (faces = 1.82, SD = .08; body postures = 2.50, SD = .10; F(1, 52) = 58.04, p < 0.0001, ŋp2 = 0.53). Replicating Experiments 1 and 2, upright stimuli (2.34, SD = .10) were more discriminable than inverted stimuli (1.98, SD = .08; F(1, 52) = 25. 18, p < .0001, ŋp2 = 0.33.) There was also a Task × Orientation interaction, F(1, 52) = 4.17, p = .05, ŋp2 = 0.07, with larger orientation effects for faces than bodies.
Results from Experiment 3. Discrimination sensitivity for face and body discrimination; error bars represent standard error. For constant conditions, the nonfocal dimension never changed (i.e., a single face or body posture was used for the whole experiment). For varied conditions, the nonfocal region was never the same, as in Experiments 1 and 2.
Importantly, variability interacted with task conditions. A Variability × Orientation interaction (F(1, 52) = 9.71, p = 0.003, ŋp2 = 0.16) indicated greater orientation effects for constant (F(1, 52) = 26.05, p = .0001,
To directly compare this study’s findings with those of Experiments 1 and 2, we conducted separate face and body discrimination analyses. For face discrimination, the Variability × Orientation repeated measures ANOVA indicated a main effect of orientation (F(1, 52) = 22.55, p < .0001,
RT produced similar patterns of performance: Discrimination was faster for bodies than faces (788 ms vs. 909 ms; F(1, 52) = 43.58, p < .0001,
In sum, the results of Experiment 3 were impressively similar to those of Experiments 1 and 2 combined. For face discrimination, body posture variation decreased visual sensitivity for upright faces and decreased the magnitude of the inversion effect. Thus, body posture cannot be ignored even when the task is only to discriminate faces. In contrast, when faces varied, visual sensitivity improved when body postures were inverted and the body inversion effect was eliminated. The differences between conditions were slightly reduced in Experiment 3, possibly due to carry-over effects from switching between face and body discrimination tasks which has been found in other studies (Richler, Bukach, & Gauthier, 2009). Despite the reduction in effect sizes, the similarity in data patterns found in Experiments 1 and 2 with those found in Experiment 3 suggest that the effect of variability on whole-body perception is robust.
General Discussion
When we view people in the real world, we often recognize them at a distance at which we can see their whole body with changing postures and views of their faces. In conversation, both facial expressions and body language are important sources of information. We pay attention to the whole person because not only are faces diagnostic to person recognition but also people’s postures are as well. Thus, to understand person perception, it is important to understand the processes underlying the interaction of face and body information when that information varies. Although research has shown that face and body information can interact, we investigated whether this integration was obligatory and whether the contributions of face and body information were symmetrical. By systematically varying variability in the irrelevant body part (i.e., changes that are nondiagnostic to task performance), we used a selective attention task with whole-body stimuli to test the degree to which participants could focus on the relevant part and ignore the irrelevant part. We also compared upright and inverted conditions to determine whether face or body integration relied on orientation-specific processing for the face or body.
In three experiments, we found evidence of obligatory but asymmetric processing of faces and body postures. Face discrimination was influenced by variation in body posture, despite instructions to only attend to the face. Importantly, the effect of variability was orientation specific: For upright but not inverted face discrimination, participants could not ignore body posture variation, even when it was not diagnostic to the task. In contrast, although body posture discrimination was also affected by face variation, participants could ignore face variation for upright body posture discrimination but not for inverted posture discrimination. Further, face variation improved sensitivity for inverted postures. Neither finding could be fully attributed to differences in discrimination difficulty between faces and body postures given the relative size of facial features compared with limbs because performance in upright and inverted discrimination conditions was not equally affected by variation.
Domain-General and Domain-Specific Mechanisms Integrate the Face and Body
The extent to which face and body information necessarily influence each other in whole-body contexts appears to depend on both domain-general attentional and face- or body-specific holistic processing mechanisms. Domain-general object recognition and attentional mechanisms allow whole-body stimuli to be represented as multiple parts (Biederman, 1987; Marr, 1982). Attention can be concurrently focused on parts and cohesive wholes, but it is stronger within a part (Vecera, Behrmann, & McGoldrick, 2000). Observers are biased to process the tops of most objects because they tend to provide important information about functionality or intention (Chambers, McBeath, Shiano, & Metz, 1999). In this study, our results suggest that participants generally focused their attention on the upper parts of the stimuli in constant conditions. However, variation in irrelevant parts revealed changes in processing by directing attention to or away from relevant regions for task performance.
For face discrimination, we found that participants could not just selectively attend to the face: Compared with constant conditions, irrelevant body posture variation reduced face discrimination performance. Domain-specific holistic processing is strongest for faces when the stimuli are upright (Robbins & McKone, 2003). When faces are part of whole-body stimuli, automatic face processing and attentional face precedence (Langton et al., 2008; Palermo & Rhodes, 2007; Rice et al., 2013; Ro et al., 2007), as well as the general upper region bias, focuses attention on facial features. However, suppose that like faces, bodies are processed more holistically when upright than when they are inverted (Harris et al., 2016; Mondloch & Maurer, 2008; Reed et al., 2003). Variation in body postures draws attention to the whole body via competing attentional body precedence and larger scale visual changes. This may lead viewers to engage a broader window of attention to incorporate both the head and body, potentially reducing attentional resources available to bind facial features into a configural whole in upright stimuli (e.g., Triesman & Gelade, 1980).
Interference from varying body postures on face discrimination is reduced when stimuli are inverted because both faces and bodies are processed less holistically, allowing better selective attention to the face which still retains attentional precedence because of task goals. Some researchers have suggested that this orientation specificity in holistic processing can be attributed to inversion producing a qualitative shift in processing to more feature or parts-based mechanisms (Maurer et al., 2002), a quantitative shift in efficiency (Richler et al., 2009), or a smaller spatial window over which holistic processing might be applied (Bukach, Bub, Gauthier, & Tarr, 2006).
For body posture discrimination, both Experiments 2 and 3 replicated the surprising finding that variation of faces influenced performance for inverted stimuli but had little effect on performance for upright stimuli. One possible explanation for this is that domain-general processes bias attention to the face or head and upper parts of upright whole bodies. Domain-specific mechanisms also favor attention to the face over the body (Downing, Bray, Rogers, & Childs, 2004; Rice et al., 2013). Thus for upright stimuli, attention is already directed to the upper parts of bodies that have the most diagnostic part information (Reed, Stone, et al., 2006), so that variation in facial features has little effect on performance. The stronger influence of face precedence is instead observed in the effects of face variation on inverted posture discrimination. Eye-tracking studies show that attention is directed to the upper portion of inverted bodies, or the leg region, when face information is constant (Arizpe et al., 2017). We propose that face variation draws attention down to the upper trunk, arm, and head region. Although one might predict that attention to the face might interfere with body posture perception, we observed that this modulation of attention improved posture judgments. This occurs even though changes to facial features are relatively smaller than changes in limb position. This counter-intuitive finding might be explained by the greater diagnostic value of the upper region of our stimuli (see Reed, Stone, et al., 2006 showing greater d′ for upper parts of postures; Chambers et al., 1999). We note that this explanation is post hoc and can be further tested by explicitly manipulating the diagnosticity of upper versus lower body parts in this type of paradigm.
An additional factor to consider is the relative location of body and parts in the visual field. Prior research suggests that there is a general bias to attend to tops of objects that is not face specific (e.g., Chambers et al., 1999), and this bias may be in operation here. Nevertheless, this factor alone cannot explain the full pattern of our results. First, a bias to attend to the upper location of objects alone would result in better selective attention to targets when they are in the upper location, making it easier to ignore variation in distractors when they are in the lower location. However, we find the opposite pattern when parts vary: Participants could not ignore variation in bodies below faces in the face recognition task (i.e., upright stimuli) nor could they ignore variation in faces below bodies in the body recognition task (inverted stimuli). Moreover, the results cannot simply be due to part location because the pattern depended on variation of the parts. Finally, the location explanation alone cannot explain why variation in the irrelevant part improved discrimination in one case and decreased discrimination in the other case. Nonetheless, to fully understand why face variation may improve inverted posture discrimination, additional experiments are needed to systematically disentangle the relative contributions from the attentional capture of faces and object processing biases to body posture discrimination.
A remaining open question is the extent to which our findings are specific to person perception, or the perception of faces and postures within the context of the whole body. Although our study was not designed to address this question specifically, and therefore did not include a nonface or nonbody control condition, there are two reasons to suggest our results may be specific to person perception. First, irrelevant part variability interacted with orientation-specific processing that is more strongly characteristic of recognition performance for faces and body postures than most other objects (Maurer et al., 2002). If the results were due merely to domain general attentional mechanisms, we would not expect such a strong interaction with orientation. Second, whole bodies appear to direct attention differently from nonbody objects (e.g., Gervais, Reed, Beall, & Roberts, 2010; Reed, Grubb, & Steele, 2006). Gervais et al. (2010) found that static body postures conveying an implied action shifted covert attention in the direction of the action’s motion or consequence, despite gaze and face direction, but block figures sharing identical parts, size dimensions, and angles did not differentially shift attention.
Conclusion
Understanding interactions between the face and body has implications for person perception. This study demonstrates that variation has a powerful impact on how body and face information are processed and the extent to which their processing is obligatory. In other words, the strength of face-body integration depends on variation. We propose that lack of integration in some experiments may be due to unnatural constancy variation. To answer the question of whether faces and bodies are necessarily integrated during person perception, we can say yes—but how we attend, and therefore process, faces and bodies depend on whether body posture is free to vary. Our results also suggest asymmetries in the obligatory processing of faces and body. Whether it varies or not, face information may be difficult to ignore. However, it is only when body posture varies that it cannot be ignored. Researchers should consider the visual and attentional mechanisms in operation when faces and bodies are viewed together and their characteristics are allowed to vary in ecologically valid ways. When viewing people at a distance in the real world, different body types and movement characteristics may help disambiguate who they are, especially if they have similar faces.
Footnotes
Acknowledgements
The authors thank Kathryn Mgrublian, Rebecca Manzella, Libby Friede, Michael Strom, Madison Lodge, Adrienne Jo, and Emilia Hagen for their help in collecting data.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded in part from the James S. McDonnell Fellowship awarded to the second author.