
Abstract
Procedural memory consolidation has been shown to enhance a variety of perceptual and motor skills during sleep, but only recently has this effect been investigated in trained musicians performing music. I tested the extent to which expected music performance skill improves over a night of sleep and to what extent the new skill memory may be fragile following initial learning. Musicians (N = 60) learned a 13-note piano melody in individual evening training sessions under one of four practice conditions. Performance of the target melody was tested the next morning following a night of sleep. In line with previous research findings, subjects showed evidence of overnight gains in performance speed and accuracy. However, these results reveal, for the first time with musicians, that additional practice on a second, similar melody seems to inhibit these overnight gains, perhaps indicating that the overnight consolidation of new skill memories is susceptible to interference from similar tasks.
The content of practice sessions and the strategies employed to develop and maintain skills have been topics of research both in and outside of music. Yet much remains to be understood about the neural mechanisms that lead over time to the mastery of intricately complex behavior like music performance. Of course it is possible to learn, practice, teach, and perform music well without a great deal of insight into these underlying mechanisms, as musicians have done for centuries. But a deeper understanding of the fundamental machinery of motor learning may not only clarify the nature of developing skills, but also may suggest strategies for maximizing the effectiveness of practice and pedagogy.
Over the past several decades, a substantive body of research into the nature of skill learning has emerged outside of music, predominantly in the fields of kinesiology, psychology, and neuroscience (for reviews see McGaugh, 2000; Walker & Stickgold, 2006). The formation and refinement of procedural memories, and specifically of memories for motor skills, has more recently received considerable attention, though the field of music is only beginning to integrate the findings from this research into our understanding of music performance, practice, and teaching (Cash, 2009; Duke, Allen, Cash, & Simmons, 2009; Duke & Davis, 2006; Simmons, 2012; Simmons & Duke, 2006).
The time course of procedural memory formation is not well characterized, nor are the factors that may interfere with memory storage and retrieval. It is clear, however, that active practice of a new skill, like playing an instrument, typically yields observable improvements while practice is ongoing. Subsequently, during the minutes and hours following practice, new memories undergo a covert process of consolidation that increases their stability and resistance to interference and forgetting (e.g., Brashers-Krug, Shadmehr, & Bizzi, 1996; Fischer, Hallschmid, Elsner, & Born, 2002; Muellbacher et al., 2002; Walker, Brakefield, Hobson, & Stickgold, 2003; Walker, Brakefield, Morgan, Hobson, & Stickgold, 2002). Neuroimaging techniques have revealed that the portions of the brain associated with skill learning remain quite active following practice, independent of conscious awareness (Albert, Robertson, & Miall, 2009; Albouy et al., 2008; Fischer et al., 2002; Korman, Raz, Flash, & Karni, 2003; Muellbacher et al., 2002; Peigneux et al., 2006).
The creation of new proteins is essential to neural plasticity following learning, and the role of protein synthesis in memory formation has been studied through the use of chemical protein synthesis inhibitors (known to prevent the creation of new synaptic terminals) in rats. Protein synthesis inhibition immediately following the learning of a shock-avoidance task, for example, has been shown to virtually erase the effects of learning; the administration of the same inhibitors four to six hours after the learning trials produce no decrements in performance of the learned task (Milekic & Alberini, 2002; Trepel & Racine, 1999).
Similar effects have been observed in human learning as well. Repetitive transcranial magnetic stimulation (rTMS), when applied over human primary motor cortex immediately following practice on a serial motor task, can also interfere with procedural memories for newly learned skills. And, as with the experiments with rats, rTMS applied six hours following practice creates no detectable interference and no observable effects on learners’ task performance (Kantak, Sullivan, Fisher, Knowlton, & Winstein, 2010; Muellbacher et al., 2002).
These and similar findings show that memory formation typically continues long after active practice ends, and that the period of overnight sleep following the learning of new motor skills facilitates further stabilization and refinement of the memories’ physical substrate, changes that are reflected in enhanced performance following sleep (Brashers-Krug et al., 1996; Duke & Davis, 2006; Korman et al., 2003; Simmons & Duke, 2006; Walker et al., 2002). Walker (2005) described two distinct phases of memory consolidation: stabilization, which occurs during waking hours following practice, and enhancement which, for explicit procedural skills, typically occurs during sleep (Walker, 2005). These processes have been shown to be independent of circadian effects (Walker et al., 2002, 2003).
Memory consolidation has also been explored with skilled musicians performing musical tasks. Simmons and Duke (2006) demonstrated that musicians learning brief melodies showed evidence of sleep-based enhancements in the accuracy of their performances. Other investigations in music have further illuminated the variables that influence the formation and refinement of skill memories following practice, such as tempo variations, learner experience, and timing of brief rest breaks during practice (Cash, 2009; Duke et al., 2009; Simmons, 2012).
A number of studies have demonstrated that, when learners practice two novel tasks in the same session, learning the second task may interfere with the consolidation of the task learned first and may reduce or eliminate the offline improvements in the first task that would typically be observed following sleep (Balas, Roitenberg, Giladi, & Karni, 2007; Brashers-Krug et al., 1996; Dorfberger, Adi-Japha, & Karni, 2007; Fischer, Nitschke, Melchert, Erdmann, & Born, 2005; Korman et al., 2003; Shadmehr & Brashers-Krug, 1997; Walker et al., 2003). This hypothesized competition for consolidation-related resources has been examined in several studies involving two similar tasks learned in juxtaposition. Offline gains in a primary task were evident in Shadmehr and Brashers-Krug’s (1997) study when participants learned a second task 5.5 hours after practice on the primary task, although not when the second task was learned only 5 minutes, 30 minutes, or 2.5 hours after learning the primary task. Walker et al. (2003), measuring participants’ speed and accuracy on a five-element sequential key-press task, found that, following a night of sleep, the participants performed more quickly and accurately than they had at the end of the training session. A second group of participants, who practiced a second sequence immediately after learning the first, showed no evidence of post-sleep performance enhancements in the sequence learned first. Many of Walker et al.’s results were replicated by Duke and Davis (2006) whose study participants practiced five-element sequences on a piano keyboard. Interestingly, Balas et al. (2007) found that even practicing a second unrelated, but familiar, complex task (in this case, a handwriting task) immediately following training interfered with subsequent consolidation of the task learned first.
As yet, no study has investigated the potential effects of interference created when musicians learn multiple novel tasks in a single practice session. Of course, this is something that musicians do all the time. My aim in this study was to examine factors related to memory formation and stabilization in musicians. Specifically, I measured the effect of sleep on musicians’ performance of a novel melody, and tested whether potential sleep-based enhancements in a target melody would be inhibited by learning two unfamiliar melodies in a single training session.
Method
Participants
Participants were 60 music majors enrolled in undergraduate and graduate degree programs at The University of Texas at Austin. All had completed no more than four semesters of undergraduate group piano instruction and had taken no more than three years of private piano instruction. All participants gave informed consent and received monetary compensation for their participation.
Design
I created four groups to test the effects of learning a second melody on the skill memory of the first (target) melody (see Figure 1). One group (A-sleep-a; n = 15) practiced only melody A (see Figure 2) for 12 30-second blocks (signified by the capital letter A in the group name) in the evening and then returned, following sleep, for a brief test (three blocks) on melody A (signified by the lower-case letter a).
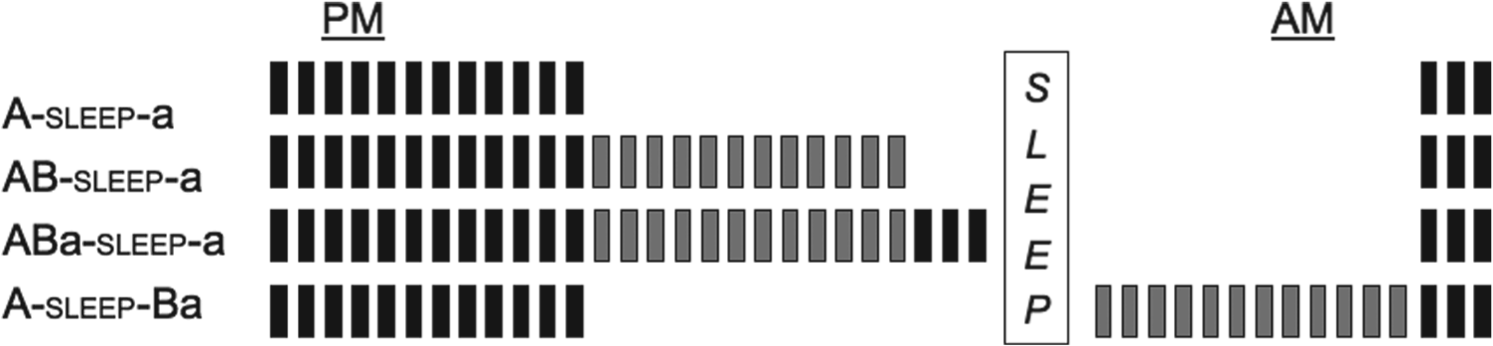
Design of the four experimental groups. Each rectangle represents a 30-second practice/re-test block (black = melody A, grey = melody B), with 30 seconds rest between each block. PM and AM sessions were approximately 12 hours apart.

Written notation and fingerings for melody A.
Another group (AB-sleep-a; n = 15) practiced melody A and then a second melody (melody B, see Figure 4) during the evening training session. They returned the following morning and performed a three-block test on melody A.
Participants in a third group (ABa-sleep-a; n = 15) also practiced melody A for 12 30-second blocks, practiced melody B for 12 blocks, and then were briefly tested on melody A at the end of the evening session. This group was also re-tested on melody A following sleep.
In order to determine the extent to which a consolidated memory would be resistant to interference from extended practice on a different melody, I included a fourth group (A-sleep-Ba; n = 15) who practiced melody A in the evening session, slept, and then returned in the morning to practice melody B before the test on melody A.
Materials
Musical instrument digital interface (MIDI) data for the piano performances were recorded using computer software written specifically for this experiment. In analyzing the data, I measured performance of the primary task (melody A) in terms of the number of correct key presses per 30-second block (CKP/B), a single measure that reflects both speed and accuracy. The CKP/B scores for each participant were then averaged over consecutive three-block triplets (see Figure 3), a method used previously in other research (e.g., Cash, 2009).
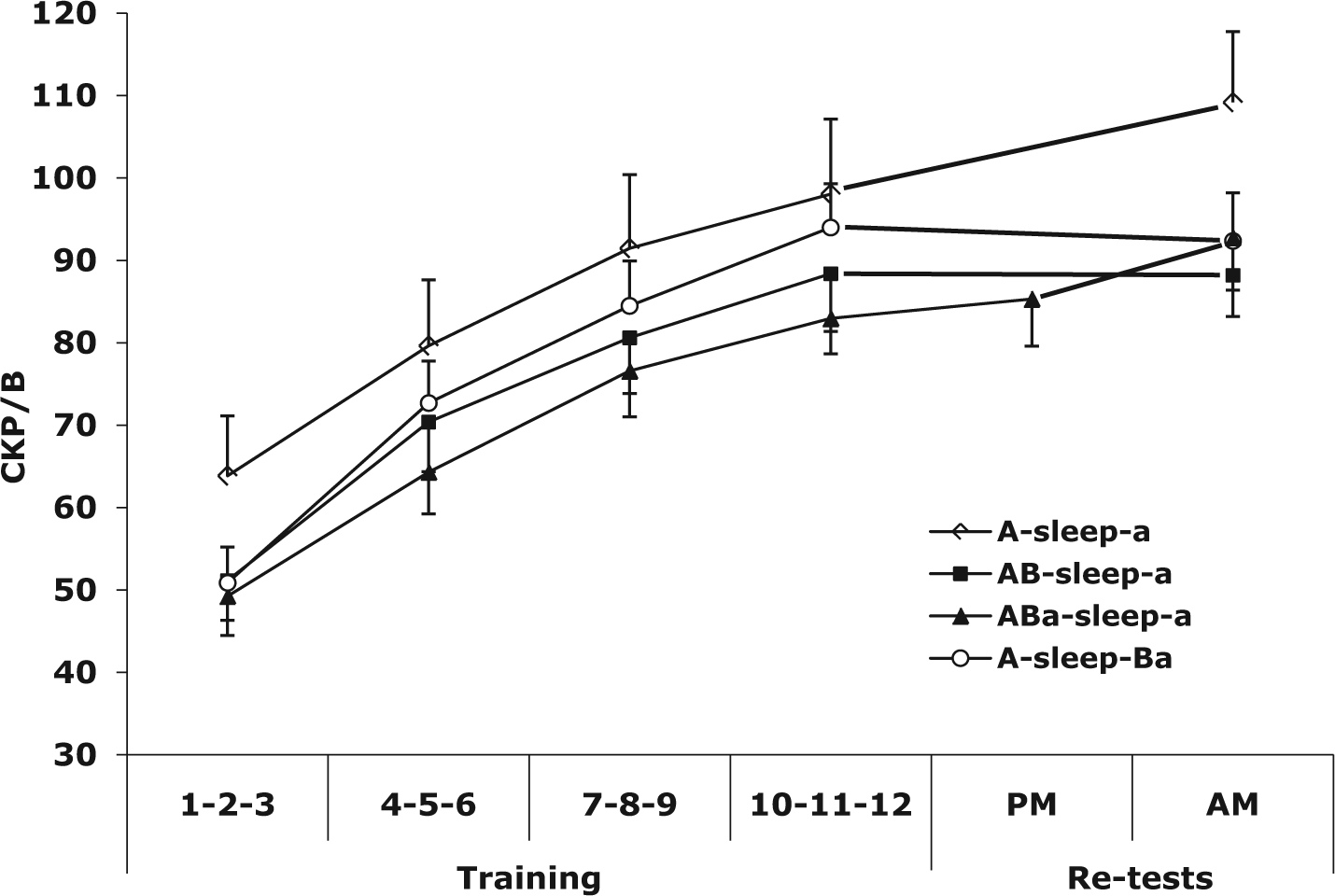
Group CKP/B means for training and re

Written notation and fingerings for melody B.
Procedure
In individual appointments with me, participants sat at a Roland KR-375 digital piano with a 12-inch laptop screen visible in front of them. They practiced the melodies with their left (nondominant) hand on the piano using fingerings indicated on the screen during 12 30-second practice blocks separated by 30-second rest intervals. Participants were instructed to repeat each melody from beginning to end ‘as quickly and accurately as possible’ throughout each 30-second block. The target melody (Figure 2) was adapted from a previous study (Simmons & Duke, 2006). Music notation and fingerings were visible on the computer screen at all times. All participants reported for two sessions approximately 12 hours apart, one in the evening and one on the subsequent morning, following overnight sleep. Participants were instructed to get a full night’s sleep following practice, and to avoid practicing any instrument before re-test.
Results
Exploratory data analysis indicated that five participants represented outliers within their respective groups. I defined outliers as scores that were either (1) more than 1.5 times the group interquartile range above the 75th percentile or (2) more than 1.5 times the group interquartile range below the 25th percentile. Inclusion of these outliers in the data set understandably resulted in heterogeneity of variances according to Levene’s test. Thus in all subsequent analyses of CKP/B, I omitted two participants from A-sleep-a (resulting n = 13), one participant from AB-sleep-a (resulting n = 14), and two participants from A-sleep-Ba (resulting n = 13).
Subjects rated their feeling of alertness at all sessions using the Stanford Sleepiness Scale (Hoddes, Zarcone, Smythe, Phillips, & Dement, 1973), and there were no significant differences in sleepiness among groups at the beginning of training, F(3,51) = 0.76, p = .52, or at test, F(3,51) = 0.80, p = .50. There were no significant correlations between reported alertness at training and performance at the beginning of training, r(53) = −0.06, p = .66, or between reported alertness at test and performance at the beginning of test, r(53) = 0.03, p = .83. There were also no significant correlations between reported alertness and performance gains during training, r(53) = 0.06, p = .66, or gains between training and test sessions, r(53) = 0.01, p = .94.
I analyzed participants’ performances at the end of training and at test using a two-way mixed analysis of variance (ANOVA): group × time point (the mean of the last three blocks of training and mean of the three morning test blocks, with time point as the repeated measure). Results indicated a significant overall increase in CKP/B between the end of training and test, F(1, 51) = 6.83, p = .012; no significant difference among groups, F(3, 51) = 1.30, p = .28; and a significant interaction between group and time point, F(3, 51) = 3.25, p = .029, indicating differential effects of learning conditions on overnight consolidation. 1
In line with existing memory consolidation research, I observed evidence of delayed offline performance gains in the A-sleep-a group, who learned a single melody (melody A) in the evening and were tested the following morning, after overnight sleep. This group exhibited significant improvement in CKP/B from the end of training to test, improving 11.4%, from a group mean of 98.04 CKP/B (SE = 9.09) to 109.17 CKP/B (SE = 8.60), t(12) = 3.77, p = .003.
In contrast, the AB-sleep-a group, who learned a second melody (melody B) immediately after learning the target melody (melody A) showed no overnight enhancement in melody A, performing very similarly at the end of training and at test (training = 88.4 CKP/B, SE = 7.00; test = 88.2 CKP/B, SE = 5.02), t(13) = 0.047, p = .96.
The purpose of the ABa-sleep-a group was to test the extent to which the interference from learning melody B was immediate (i.e., observable during the training session) or delayed (i.e., during overnight consolidation). This group was tested briefly on melody A after having learned melody B, and showed no decrement in their performance of melody A. The mean CKP/B for melody A at the end of the 12 blocks of training was 83.0 CKP/B (SE = 4.32); after learning melody B, the mean CKP/B for melody A was 85.3 CKP/B (SE = 5.72) t(14) = −0.86 p = .40. Following sleep, however, the mean CKP/B for melody A increased by 11.8% from the end of training (training = 83.0 CKP/B (SE = 4.32): morning re-test = 92.8 CKP/B, SE = 6.40), t(14) = 2.39; p = .031. Thus the brief test of melody A (three blocks) following the learning of melody B at the end of the evening training session seems to have reactivated the memory of melody A in a way that inhibited the interfering effects of learning melody B that were observed in the AB-sleep-a group. This was the most surprising, and perhaps the most important, result of the study: that the recall of melody A after extended practice on melody B resulted in overnight enhancement in melody A similar to that observed in the A-sleep-a group.
The A-sleep-Ba group learned melody A in the evening training session, and then in the morning session, prior to the test of melody A, learned melody B. The learning of melody B prior to the test of melody A obscured any enhancements in melody A that may have occurred as a result of overnight consolidation. At the end of the evening training session, the mean CKP/B for melody A was 94.0 (SE =3.47); in the morning test session, following 12 blocks of practice on melody B, the mean CKP/B for melody A was 92.4 (SE = 4.68), t(12) = 0.52, p = .61.
Discussion
The present study is one of a growing line of investigations that systematically examine procedural memory consolidation, specifically the evolution of consolidation-based stabilization and the subsequent consolidation-based enhancement in musicians learning musical tasks. Like the findings obtained in similar research, the results of the current study show that memories for newly learned music skills are enhanced during post-training sleep. The participants who learned only one melody in the evening training session and returned for a brief test of the same melody following overnight sleep (A-sleep-a) clearly demonstrated improvements in their performance (11.4% increase in CKP/B), consistent with offline, overnight gains observed in similar sequence-learning tasks (Balas et al., 2007; Dorfberger et al., 2007; Duke et al., 2009; Duke & Davis, 2006; Kuriyama, Stickgold, & Walker, 2004; Simmons & Duke, 2006).
The continuation of learning offline, after practice on a new task has ended, is clearly relevant to music teachers and learners. Although musicians may often notice in their own practicing that a difficult passage feels easier after time away from active practice, there has been little discussion in music of the mechanisms that facilitate the refinement of new skill memories offline. Systematic research in this area, however, is beginning to provide insight into the way music is learned, refined, stored, and recalled.
The current study also demonstrates in a limited context that the overnight enhancement of a newly learned musical task can be inhibited by learning a second, competing task during the same practice session, an effect that has been reliably demonstrated in contexts outside of music. The group of participants in this study who learned two melodies in juxtaposition during the evening training session (AB-sleep-a) made no offline performance gains in the first melody at the morning test, evidence that learning the second melody (melody B) inhibited subsequent offline gains that would have been expected in the morning test of melody A. My results are consistent with the findings reported in a growing number of studies examining the relative stability of new skill memories following practice (Balas, et al., 2007; Dorfberger et al., 2007; Fischer et al., 2005).
It is believed that similar tasks learned in close temporal proximity may compete for the same neural resources required for consolidation following practice. Of course, since we encounter so many new stimuli in a day, the brain must selectively consolidate those memories that are the most meaningful, important, and potentially beneficial.
It has been shown that music skills show evidence of sleep-based enhancements attributable to consolidation (Cash, 2009; Simmons, 2012; Simmons & Duke, 2006) but prior to the current study the potential for multiple musical tasks to interfere with consolidation was unknown. Of course, musicians typically practice multiple skills in a single practice session, and the results of the present study indicate that new skill memories are susceptible to interference following practice, such that practice on similar tasks in a single training session may interfere with subsequent consolidation-based enhancement of the task learned first.
Originally intended as a control group, the ABa-sleep-a group showed, as predicted, that learning a second melody prior to sleep did not immediately interfere with the memory of melody A. Somewhat surprising was the result that recalling melody A immediately after having learned melody B during training led to consolidation-based enhancements in melody A overnight, enhancements that were not observed when melody A was not recalled at the end of training (as in the AB-sleep-a group). In other words, the recall of melody A in this experiment may have triggered overnight consolidation-based enhancement that might otherwise have been blocked by practice on melody B, as was observed in the AB-sleep-a group.
Walker and his colleagues used a similar procedure in their study of consolidation (2003), in which participants learned two sequences in a single training session and were then immediately tested on both. Learners showed no change in performance from the end of training to test on either sequence. Unlike the present study, though, Walker et al. did not re-test this group again after sleep. The group in Walker’s study was designed as a control to demonstrate that the interference effects of learning the second sequence did not immediately affect the memory of the first sequence in any way, but rather affected the overnight enhancement of the first sequence (as Walker et al. observed in a different experimental group). No study to date has examined the effect on overnight consolidation following a brief recall of a learned task (e.g., melody A in the current study) after having learned a second, similar task. Further research is certainly needed to explore this finding further.
The A-sleep-Ba group did not show performance improvements on melody A upon morning test; they were the only group, however, to recall melody A immediately after extended practice on melody B in the morning session. It seems as though the proximity to learning melody B, and the concentration that it required in the morning session, made the immediate recall of melody A difficult for this group.
Several dimensions of this study add to the existing literature on motor memory consolidation. First, I examined the process of skill acquisition and consolidation in the context of experienced learners. Although researchers have more recently begun to examine differences in the learning process attributed to differences in task familiarity (Balas et al., 2007), task complexity (Kuriyama et al., 2004), and differences in age (Dorfberger et al., 2007), little is yet known about motor skill acquisition and consolidation in skilled learners.
Second, I examined the process of skill acquisition and consolidation for complex tasks. The task in this study was challenging and complicated for learners, not only because of the sequence length (13 key-presses rather than the five key-presses often used in other studies), but also because of the added auditory processing demands. Participants had not only to read the music notation and fingerings on the screen in bass clef (as using finger numbers alone was not sufficient – each finger was used for more than one pitch), but also to respond to the sounds they were hearing (e.g., wrong notes, unevenness, hesitations). This offers insight not only into motor skill learning with musicians and musical tasks in particular, but also into skill learning that involves performing more complex tasks. Motor skill task complexity has been examined primarily in kinesiology research (e.g., Wulf, McNevin, & Shea, 2001; Wulf, Shea, & Matschiner, 1998), with few studies investigating learning and memory consolidation.
Finally, the present study replicates the findings of Simmons and Duke (2006), Duke et al. (2009), and Simmons (2012), which found consolidation-based enhancement in musicians who practiced novel motor skills, and adds to this existing research by introducing additional practice on a second task. It is important to note that all of the participants in the current investigation slept between training and test, and thus the variable of sleep was not isolated from the mere passage of time. Although many investigations, including studies in music (cited earlier), have shown that sleep is necessary for consolidation-based enhancement of motor skills, it is impossible to dissociate sleep from time in the current data.
It is as yet too early to offer prescriptions for music practice and pedagogy based on current findings. Such generalizations are unwarranted at this time and will require much additional, systematic research. How exactly procedural memories are encoded, how they are integrated into a learner’s memory store, and how they are later retrieved, refined, and then reintegrated into existing memory are complex processes that remain relatively unclear. The present findings do, however, add to our current understanding of the intricate and multi-sensory activity that comprises music skill learning.
Footnotes
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.