
Abstract
It has been argued that historical biogeography, the study of how processes that occur over long periods of time influence the distribution of life forms, is in the midst of a scientific revolution. The aim of this paper is to analyze the evolution of historical biogeography during the first decade of the 21st century and to identify major trends for the near future. We constructed a database containing all articles which dealt with historical biogeography published in the Journal of Biogeography during 1998–2010. The database included 610 contributions. Our results indicated that historical biogeography is going through a growth period. The papers analyzed were written by 2018 authors, with a mean of 3.3 authors per paper. Authors from 62 countries were involved, and most of them worked in Europe or North America. The Palearctic was the most analyzed region. Most contributions dealt with terrestrial habitats and were devoted to animal (especially Chordata) and plant taxa. Phylogeography was the most used approach (35%), followed by biota similarity and PAE (13%) and molecular biogeography (12%), with cladistic biogeography and event-based methods at 6% each. Some of the future challenges that historical biogeography faces are summarized: (1) to increase the study of taxa which are underrepresented according to the segment of biodiversity they represent; (2) to balance the amount of work devoted to different biogeographical regions; (3) to increase biogeographical knowledge of aquatic habitats; (4) to maintain the diversity of approaches, preventing the reduction of time, spatial, and taxonomic scales addressed by the discipline; and (5) to continue integrating historical biogeography along with other sources of information from other disciplines (e.g. ecology, paleontology, geology, isotope chemistry, remote sensing) into a richer context for explaining past, present, and future patterns of biodiversity on Earth.
Keywords
I Introduction
Historical biogeography studies how those processes that occur over long periods of time, for example tectonics or evolution, influence distributional patterns of life forms (Cox and Moore, 1993). Crisci (2001) argues that historical biogeography is in the midst of a scientific revolution (sensu Kuhn, 1970) and that it is a rapidly evolving discipline. The scientific revolution that historical biogeography experienced in the second half of the 20th century and the first years of the 21st century is shown in the great number of approaches to the subject (and their corresponding methodologies) that arose in the most recent decades (see Crisci et al., 2003; Posadas et al., 2006). Furthermore, several attempts have been made to characterize historical biogeographical approaches and methodologies by defining different ‘taxonomies of the methods’ (Andersson, 1996; Crisci, 2001; Humphries, 2000; Lieberman, 2000; Morrone, 2005, and literature cited therein). The aforementioned context drives us to the question: what is the present state of historical biogeography? Posadas and Donato (2007) presented a preliminary attempt to unravel this question based on the papers published in the Journal of Biogeography during 2005–2006. Here, we propose to answer this question using the same journal, but for a broader time interval.
Since the Journal of Biogeography (JB) began publication in 1974, it has become one of the most prestigious scientific publications on biogeography. In fact, Morrone and Guerrero (2008) have identified the JB as the principal core journal for biogeographical research. The editorial policy of this journal is that it seeks to be representative of the discipline of biogeography, to be global in scope, and to be inclusive of major traditions and viewpoints within the discipline (Journal of Biogeography, 2011). Therefore, we assumed that by analyzing the papers published in the JB in 1998–2010 we would obtain a good picture of: (1) the state of knowledge of historical biogeography; (2) the evolution of this discipline in the considered span; and (3) its major trends in the near future.
The state of the art of historical biogeography we present here goes further than a scientometric-bibliometric analysis, because it concerns not only major publication trends in the field (e.g. authorship trends) but also internal trends of the discipline (e.g. methodologies applied, geographic and taxonomic groups considered, use of molecular clocks).
II Data and methods
Data for the analysis were compiled from the JB. All articles that dealt with historical biogeography published in this journal in 1998–2010 were considered. The database included 610 published articles for which a series of factors were taken into account (e.g. number of authors, place of residence of the first three authors, location at the regional scale, historical biogeographical approach and technique, taxon considered, habitat, use of molecular clocks). The database includes a variety of articles (e.g. guest editorials, research papers, books reviews, comments) on a wide range of topics in the field (e.g. theoretical or empirical studies, reviews, methodological discussions). Thus, some of the items considered were not applicable to all the papers in the database – for example, taxon or area analyzed. Each result presented was accompanied by the number of papers examined for each variable. Although authorship analysis indicated a total of 2018 authors, place of residence of authors was determined only for the first three authors of each contribution; so the universe was restricted to 1431 authors (70% of the total).
Paper categorization as related to historical biogeographical approaches was based on Crisci (2001; see also Crisci et al., 2003). The taxonomy of historical biogeography presented by Crisci et al. (2003) included nine basic approaches, eight of which were found in the analyzed data set: (1) dispersalism; (2) phylogenetic biogeography (in the sense of Brundin, 1966); (3) ancestral areas; (4) panbiogeography; (5) cladistic biogeography; (6) biota similarity and PAE (parsimony analysis of endemicity); (7) event-based methods; and (8) phylogeography. It is important to note that under the label ‘biota similarity and PAE’ we included all those papers in which biota similarity was analyzed, including those analyses that were carried out using algorithms other than parsimony (e.g. Unweighted Pair Group Method with Arithmetic Mean; UPGMA). Also, the endemicity analysis (Szumik et al., 2002), the cladistic analysis of distribution and endemism (CADE; Porzecanski and Cracraft, 2005), the corrected weighted endemism technique (CWE; Crisp et al., 2001), and the method for the identification of areas of endemism using species co-occurrences (Giokas and Sfenthourakis, 2008) were included under the biota similarity and PAE approaches. Additionally, we considered here three new categories as independent approaches: (9) molecular biogeography; (10) paleoenvironmental reconstruction; and (11) combined approaches. Also, we included a category labelled as (12) ‘others’ in order to include 35 papers which corresponded to historical biogeography but did not fit in any one of the previous categories.
Molecular biogeography was defined by Lavin et al. (2000) as the reconstruction of the biogeographical history of one taxon on the basis of its cladogram obtained from molecular data. This definition is somewhat vague, because it does not point out how to reconstruct the biogeographical history of the taxon, nor that molecular-based cladograms could be applied to any approach to historical biogeography which requires phylogenies as input data (e.g. ancestral areas, dispersal-vicariance analysis, cladistic biogeography). Thus, under the label ‘molecular biogeography’, we only considered those papers which: (1) were based on molecular-based phylogenies; (2) were devoted to taxon biogeography (not area biogeography); and (3) did not qualify in any of the other approaches based on cladograms. Alternative and more comprehensive definitions of molecular biogeography have been provided by other authors (e.g. Lomolino et al., 2010; Mantooth and Riddle, 2011; Riddle et al., 2008). We preferred the definition of Lavin et al. (2000) in order to maintain the taxonomy proposed by Crisci (2001) as accurate as possible, and to keep phylogeography as a separate approach in order to observe its increasing influence in historical biogeography. The ‘paleoenvironmental reconstruction’ approach grouped those papers that attempted to define paleoenvironments, based mainly on paleontological and geological evidence. Finally, the ‘combined approaches’ category grouped those papers that used more than one of the approaches defined herein.
III Results
1 General trends of publication and authorship on historical biogeography
A total of 2095 contributions (including guest editorials, research papers, comments and book reviews) were published in the JB in 1998–2010, 29% (610 papers) of which deal with historical biogeography. The total number of contributions and that of those on historical biogeography appeared to increase during the analyzed span (Figure 1). It is interesting to note that from 1998 to 2006 historical biogeographical contributions represented less than 25% of the total, while in the 2007–2010 span this proportion was over 40%; the highest being in 2009 and 2010, years in which historical biogeographical contributions represented more than 50% of the total publications. Furthermore, while the number of total contributions published in JB more than doubled from 1998 to 2010 (i.e. from 90 in 1998 to 200 in 2010), the historical biogeographical contributions increased almost 10 times during the same period (i.e. from 11 in 1998 to 105 in 2010). Empirical papers represented 80% of the 610 historical biogeographical contributions considered here, while papers dealing with theoretical questions accounted for 10%. The remaining 10% included book reviews, replies, and comments.
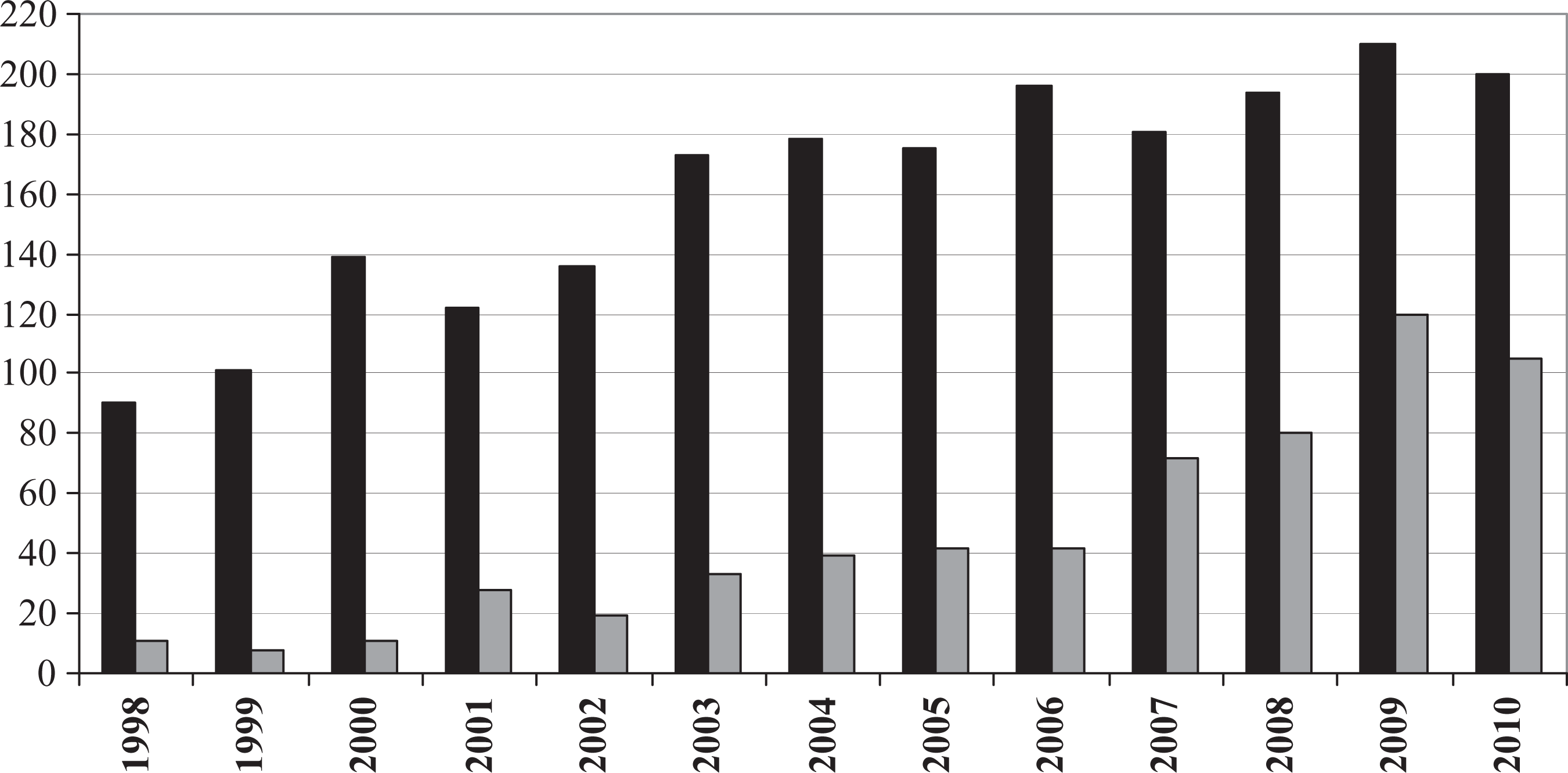
Number of articles published in each volume of the Journal of Biogeography during 1998–2010 (total = black bars; historical biogeography = gray bars).
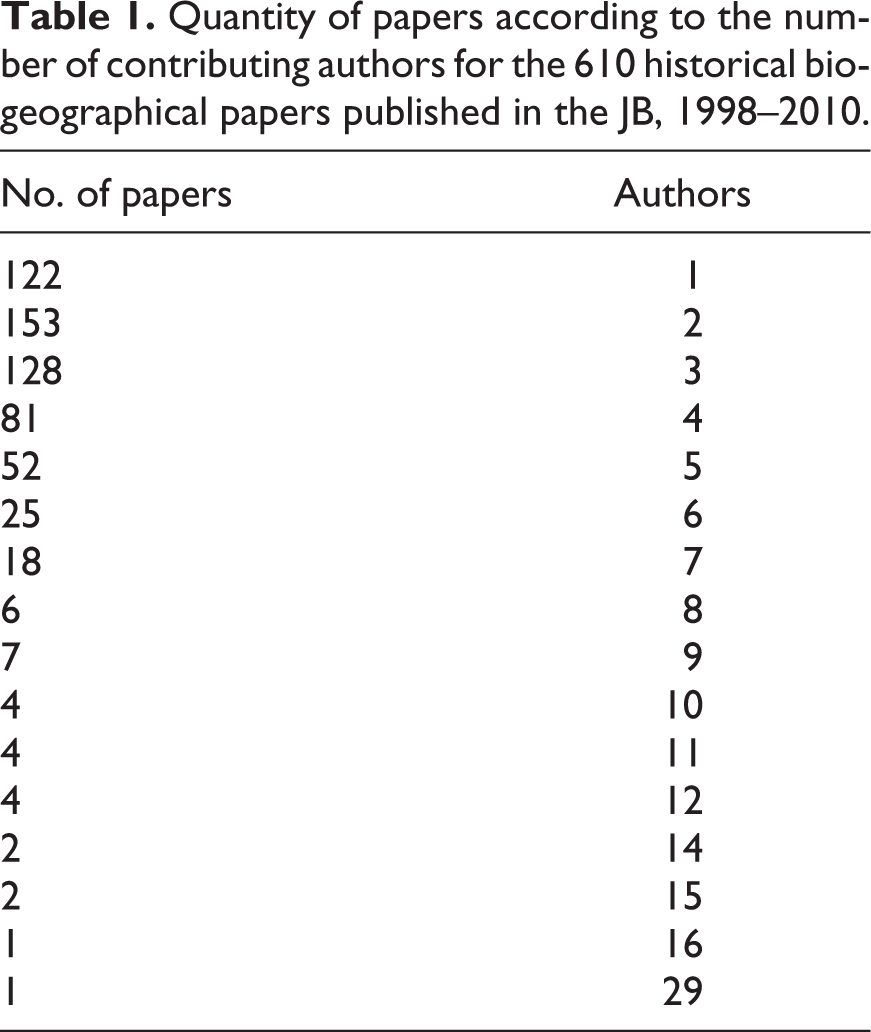
Regarding authorship, the 610 historical biogeographical papers considered here were written by 2018 authors, with a mean of 3.3 authors per paper (Table 1). Sixty-six percent of the papers were authored by one to three authors (20% with a single author, 25% with two authors, and 21% with three authors). Papers with four or more authors represented the remaining 34% of the published contributions (Table 1). The maximum number of co-authors for a paper was 29 (i.e. Pickett et al., 2004). A trend toward more authors per paper is evident (Figure 2), especially after 2004, when the percentage of papers written by a single author decreased, while the percentage of papers authored by four or more authors increased.
Quantity of papers according to the number of contributing authors for the 610 historical biogeographical papers published in the JB, 1998–2010.
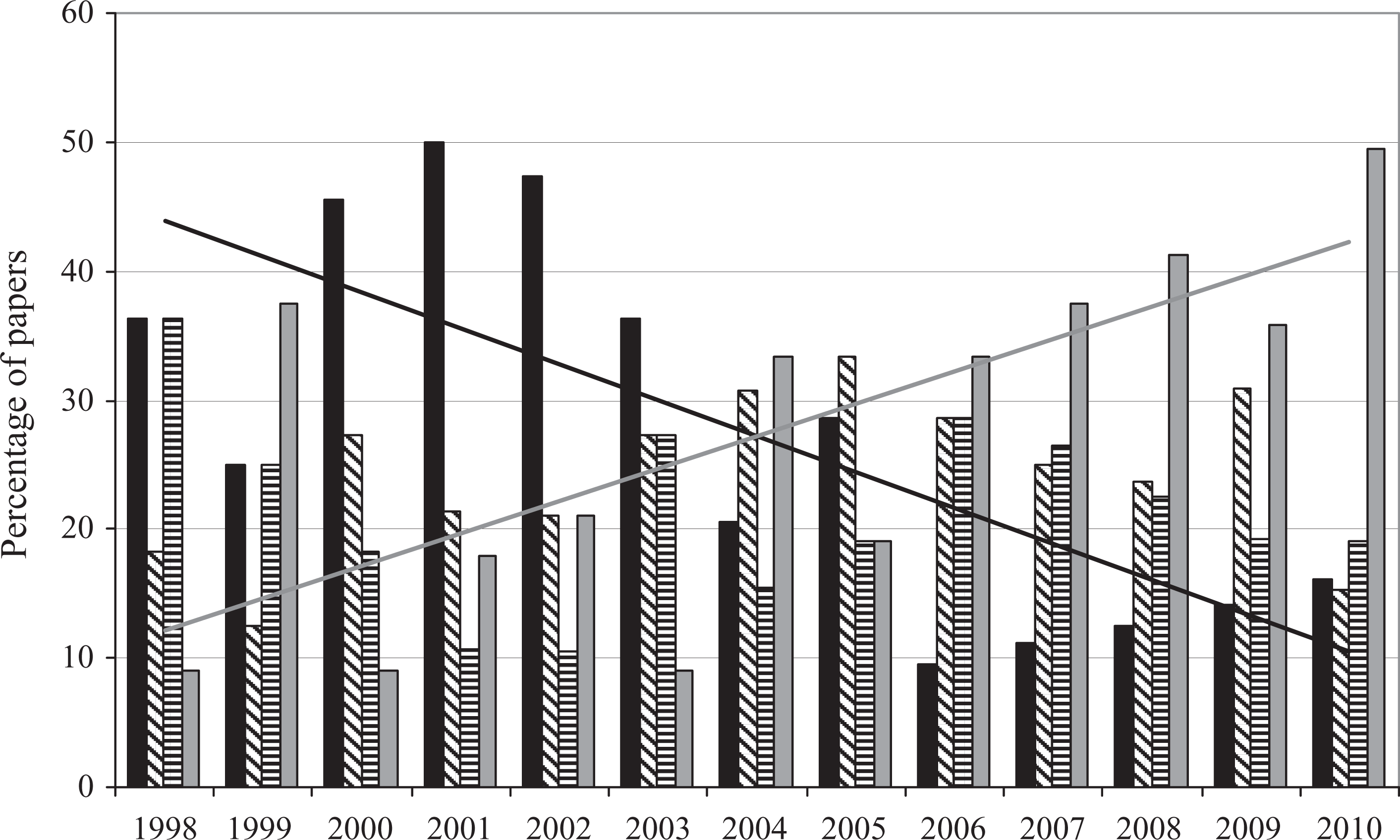
Percentage of papers authored by one (black bars), two (diagonal line bars), three (horizontal line bars), and four or more authors (gray bars) per year. Trend lines added for papers authored by a single author (black) and by four or more authors (gray).
When author’s place of residence was analyzed at a continental level (Figure 3), we found that most authors were European and North American (43% and 26%, respectively). Latin-American plus Caribbean (LAC) and Australian plus Polynesian accounted for 23% of authorships (12% and 11%, respectively). The remaining 8% were by African (3%) and Asian (5%) authors. The increasing number of authorships from each continent was a general trend during the span considered (Figure 4). Notwithstanding, this trend was specially marked for European and North American authors. Almost throughout the entire period, most authorship was by European and North American researchers. Between 1998 and 2005, the number of authorships from these two continents was practically equivalent. Since 2006, the number of authorships in both aforementioned continents has increased greatly, although this trend was more pronounced in Europe (Table 2).
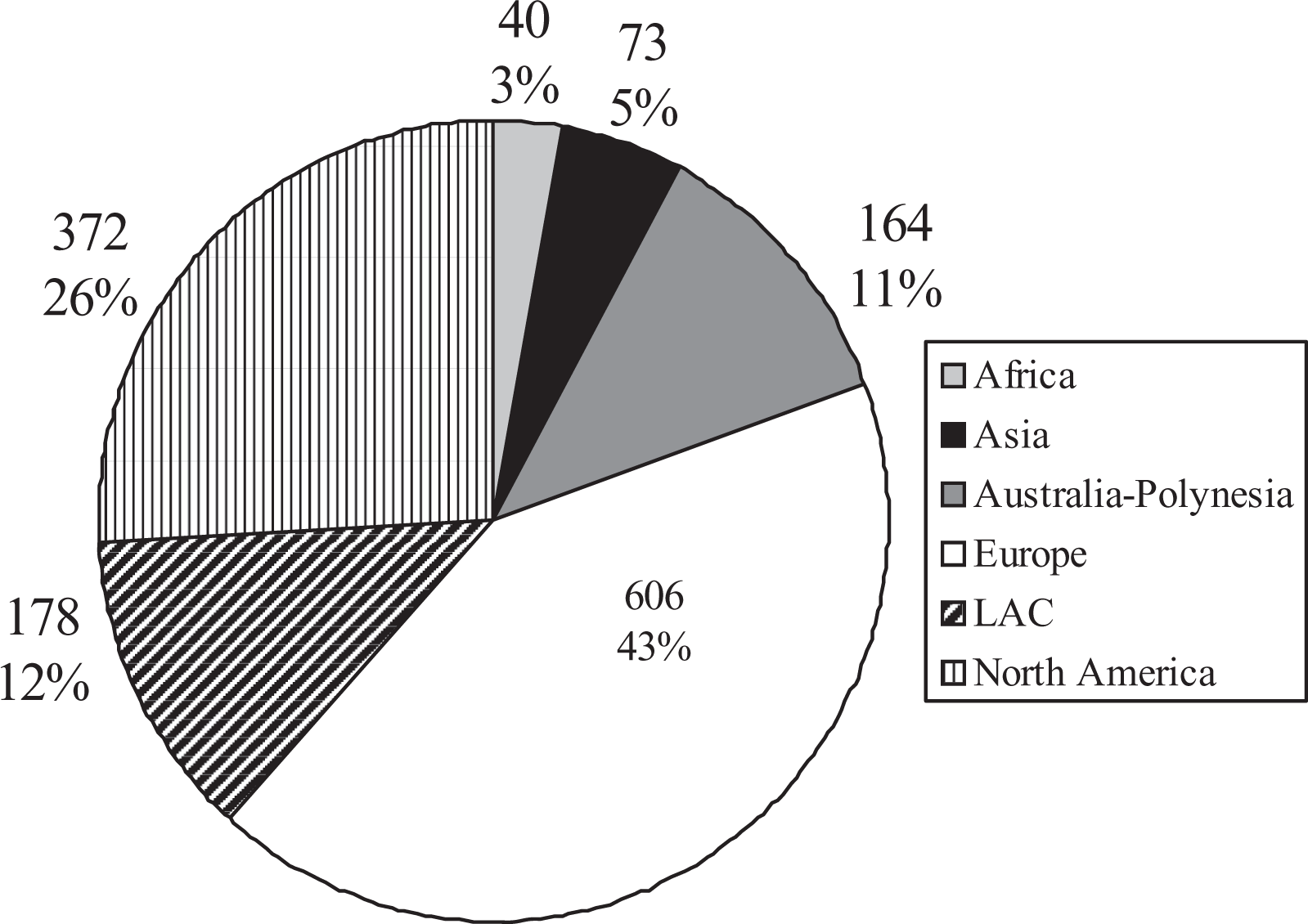
Contributing authors’ place of residence at the continental level. Only the first three authors of each paper were considered.
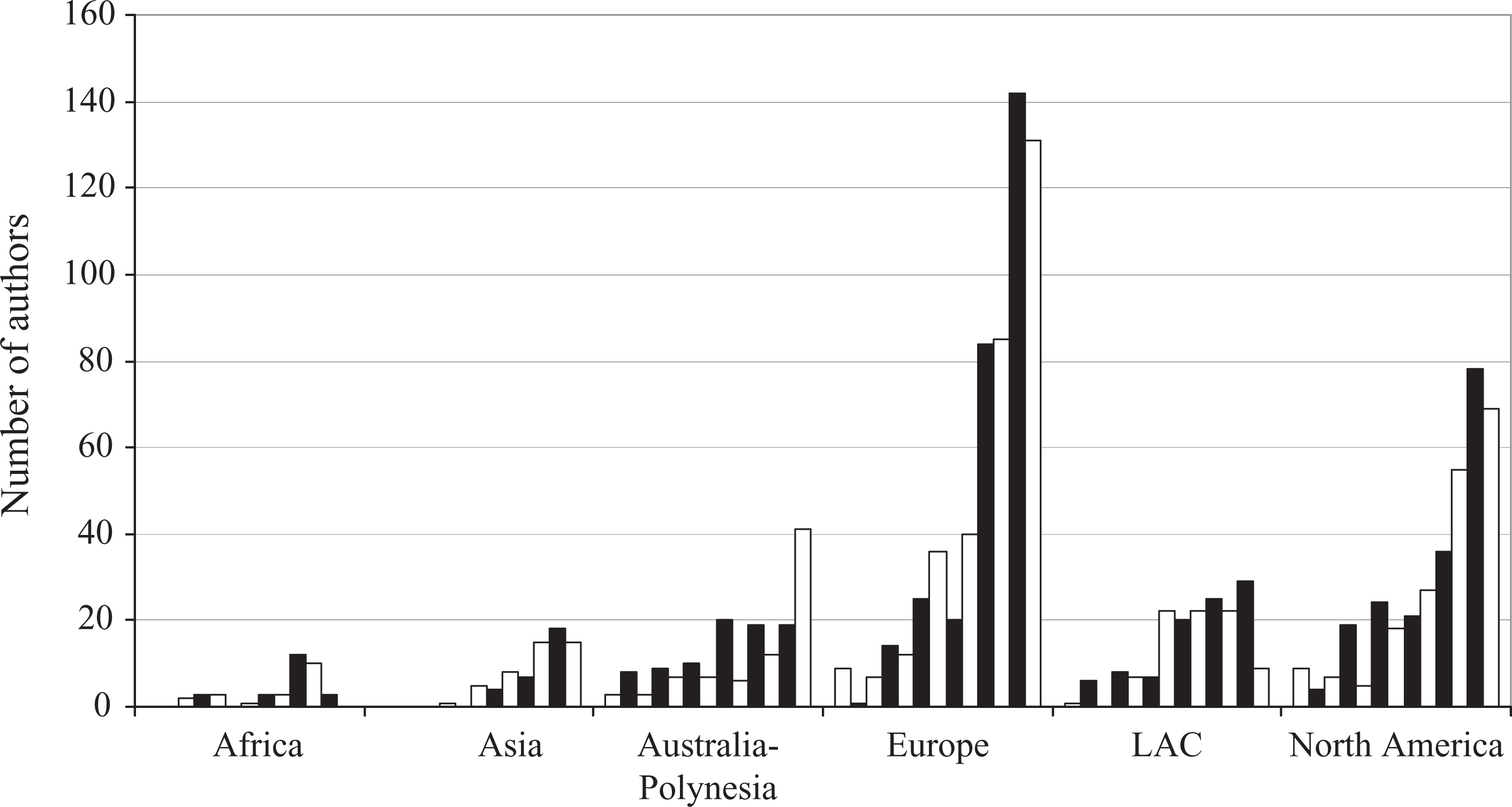
Residence of contributing authors at the continental level per year from 1998 to 2010. Only the first three authors of each paper were considered. White bars = even years; black bars = odd years.
Number of authorships per year by place of residence at the continental level (only the first three authors of each of the 610 papers were considered).
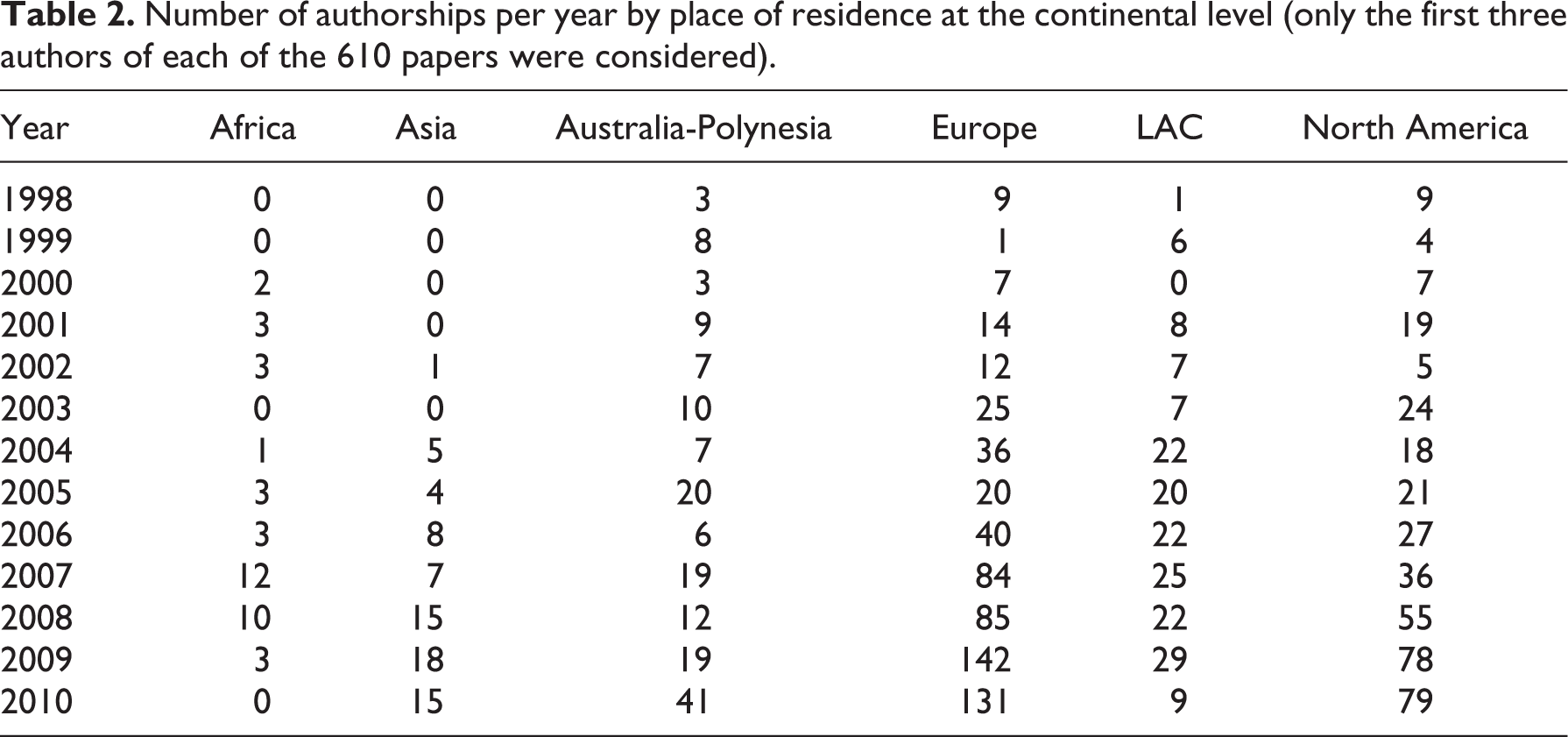
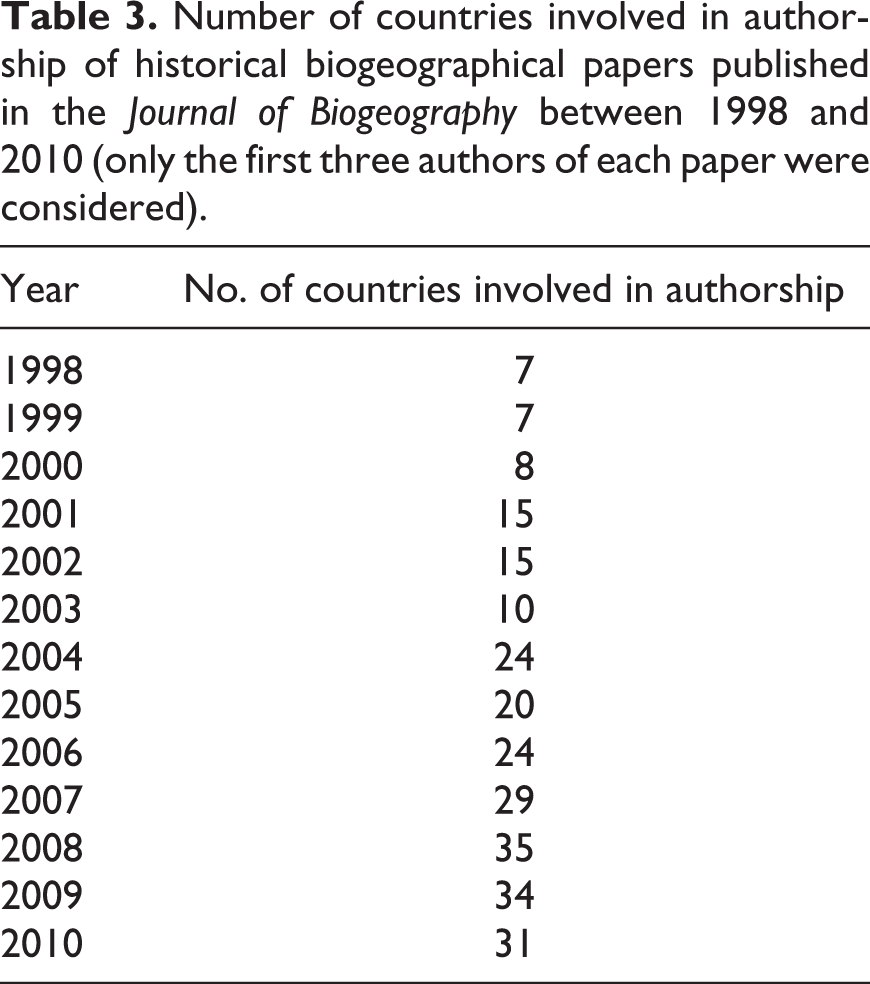
Analysis of authorship at the country level showed that authors from 62 countries authored papers on historical biogeography in the analyzed period. The United States of America (USA) contributed the most authorship, followed by the United Kingdom (UK), Australia and Spain (Figure 5). The number of countries from which authors contributed historical biogeographical papers increased as well during the 13 years considered (Table 3).

Country of residence of contributing authors. Only the first three authors of each paper were considered. (Black = first author; grey = second author; white = third author; NA = North America; LAC = Latin America and Caribbean.)
Number of countries involved in authorship of historical biogeographical papers published in the Journal of Biogeography between 1998 and 2010 (only the first three authors of each paper were considered).
2 Internal trends in historical biogeography
a Habitats, areas, taxa, and events
Seventy-five percent of the analyzed papers were devoted to terrestrial habitats. Aquatic habitats were the subject of the remaining 25%, with 12% freshwater habitats and 13% marine habitats (N = 540).
When major biogeographical regions were analyzed (N=545), we observed that more than one region was considered in 22% of the papers. Oceanic areas (Indian, Pacific, and Atlantic) were considered in 4% of the papers. A single continental region was considered in 74% of the contributions, with representation as follows: 35% on the Palearctic region; 11% on the Nearctic region and 4% on the Holarctic region as a whole; 15% on the Australasian region; 7% on the Ethiopic region; 1% on the Cape region; 14% on the Neotropical region; 4% on the Andean region; 5% on the Oriental region; 3% on the Wallaceae; and 1% on the Antarctic region.
From a taxonomic point of view, at the kingdom level (N=532) it was observed that 59% of the papers dealt with animal taxa, 35% with plant taxa, and 4% with taxa from more than one kingdom; only 2% of the papers dealt with taxa from other kingdoms (e.g. Protoctista, Fungi). When those papers which involved animal taxa were analyzed at the phylum taxonomic rank (N=312), we found that most studies were devoted to Chordata (54%), followed by Arthropoda (26%), and Mollusca (7%). Echinodermata, Cnidaria, Nematoda, and Annelida accounted for the remaining 13%. Within Chordata, 31% of the papers dealt with mammals; 26% with fishes; 17% with birds; 13% with reptiles; 10% with amphibians; and less than 1% with ascidians.
When those papers which involved plant taxa were analyzed at the division taxonomic rank (N=120), we found that 83% of the papers dealt with Magnoliophyta (‘Angiosperm’ or flowering plants), 12% with ‘Gymnosperms’, and 5% with other higher taxa (i.e. Pteridophyta – ferns – and Bryophyta – mosses). Within the ‘Angiosperms’, 85% of the papers dealt with dicotyledonous plants and 15% with monocotyledons. Fifty families of dicots were considered in the 85 papers dealing with this taxon; Asteraceae (8%), Fabaceae (6%), and Fagaceae (6%) were the families with more representation. Ten families were represented in the 15 papers devoted to monocots, with Poaceae (27%) as the most analyzed family. Finally, 71% of the 14 papers devoted to the ‘Gymnosperms’ dealt with the Pinaceae. When taxonomic ranks were considered (N=536), 69% of the papers treated taxa at the supraspecific level and 31% focused on the infraspecific level.
Historical biogeographical events (i.e. dispersal, vicariance, and extinction) were mentioned in 76% (463) of the 610 analyzed papers as: (1) possible explanations for taxon distributions, or (2) the main subject of the research. Forty-five percent of these 463 papers considered vicariance-dispersal explanations, 24% considered or discussed only dispersal events, 25% of them dealt only with vicariance, and only 7% discussed extinction (the key extinction events being the focus of most of them).
b Approaches and techniques
When analyzing historical biogeographical approaches and techniques, not only were the empirical papers applying a given approach considered, but also those papers discussing theoretical questions regarding a particular approach (e.g. Morrone, 2005) or those presenting a new technique (e.g. Wojcicki and Brooks, 2005).
Five approaches were used in almost 72% of the papers considered (N=557). Phylogeography ranked as the most used approach, at 35% of the total, followed by biota similarity and PAE approaches (13%), molecular biogeography (12%), and cladistic biogeography and event-based methods (6% each) (Figure 6; Table 4). The other approaches applied and the publication trends in each are summarized in Figure 7. It is interesting to note the explosive increase in phylogeographical papers from 2004 to 2010 (Figure 6). Although molecular biogeography exhibited an increasing trend after 2005 (with a fall during 2008), it is not as explosive as it was in phylogeography, in which the number of contributions tripled from 2003 to 2004. The increasing number of paleoenvironmental reconstruction papers is also remarkable, and this trend has been constant since 2003.
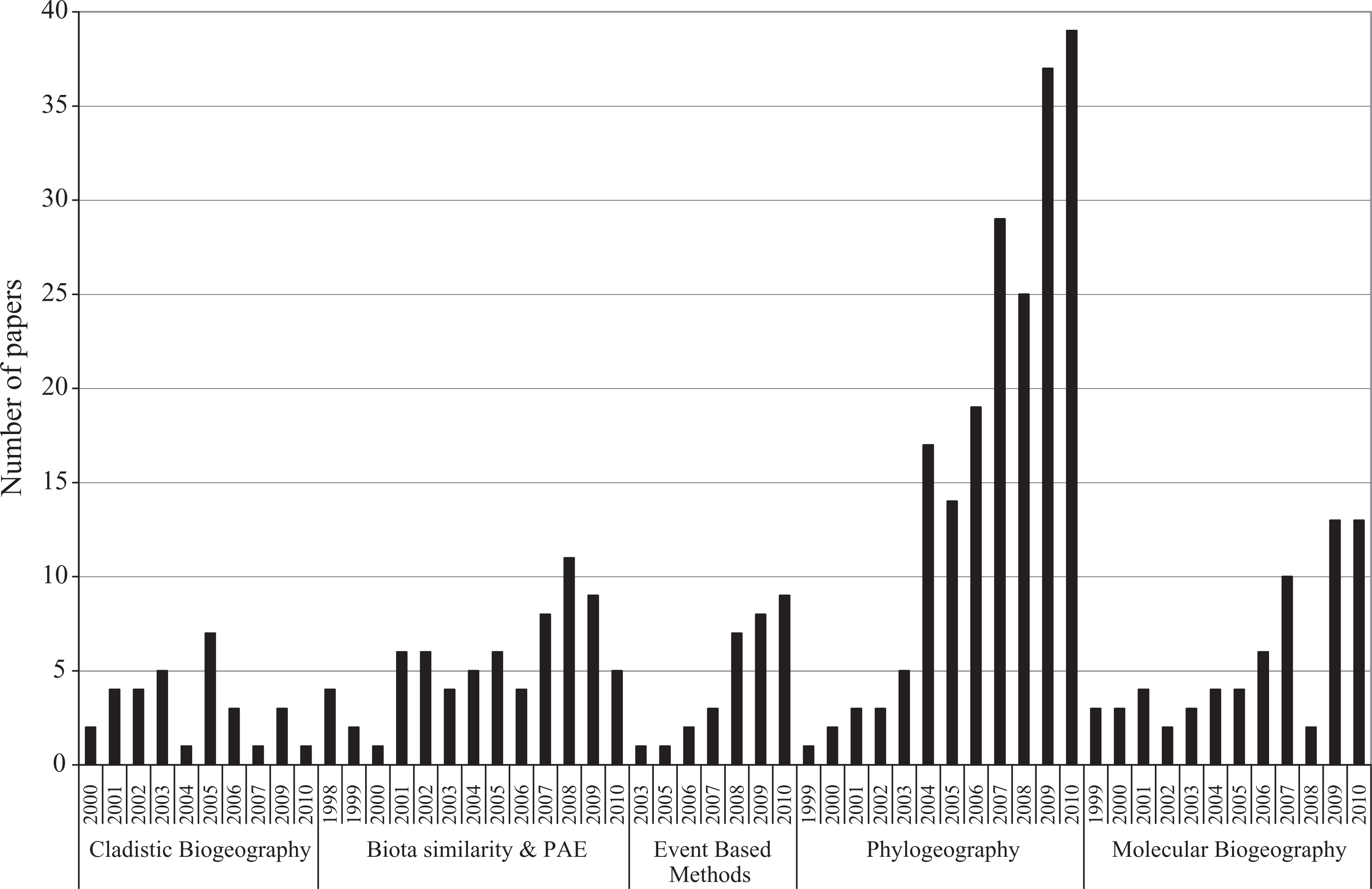
Distribution of papers applying the five most used approaches per year.
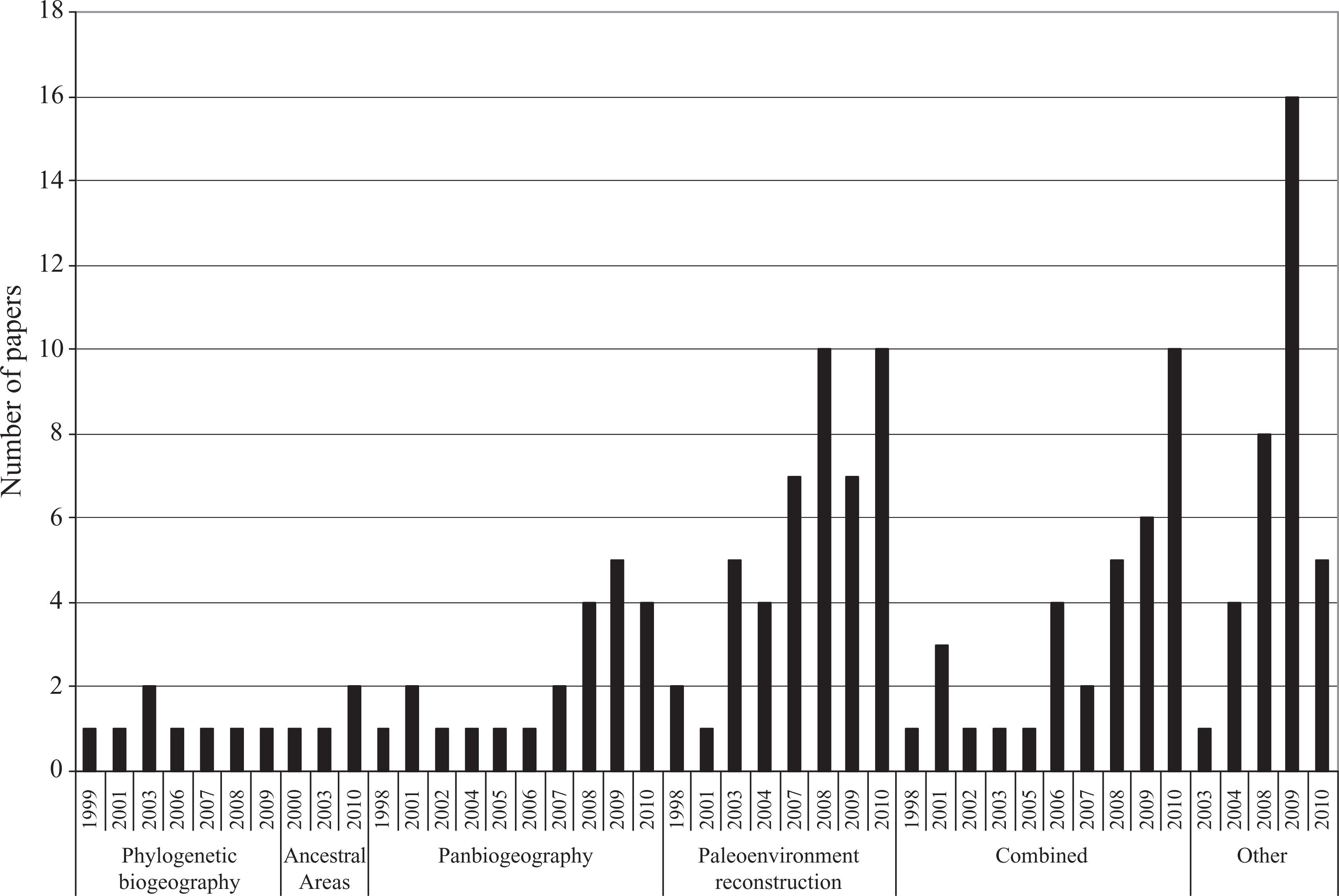
Distribution of papers applying other approaches per year.
Techniques used or discussed in the analyzed papers classified according to historical biogeographical approaches. None = no specific techniques applied or theoretical papers that did not use or discuss a specific technique. Techniques are named according to Crisci et al. (2003); exceptions are EA (endemicity analysis of Szumick et al., 2002) and PACT (phylogenetic analysis for comparing trees of Wojcicki and Brooks, 2005) which were developed after the publication of Crisci et al. (2003).

In the case of cladistic biogeography (N=31), 45% of the papers concern theoretical questions, 45% applied studies, and the remaining 10% comments and replies. This is the only area in which the number of theoretical and applied contributions is equal. In all other approaches, theoretical papers represented less than 10%.
Table 4 shows the techniques applied within each identified approach (N=557). The use of 15 different techniques was recorded, and several combinations within and between approaches were detected. The 35 papers under the category of ‘others’ included different subjects – for example, paleogeographical reconstructions, analysis of niche conservatism and niche modeling, definitions of glacial refuges, discussions related to historical versus ecological factors regarding distributions, and population genetics in a historical and geographical context.
Timing had been postulated as a priority item in historical biogeography (Crisci et al., 2003; Donoghue and Moore, 2003; Hunn and Upchurch, 2001). However, timing through molecular clocks has been considered or discussed in the 30% of the papers analyzed (N=577). Molecular biogeography and phylogeography constituted 71% of the 174 papers which applied molecular clocks. This 30% could be viewed as a bias due to the variety of approach categories analyzed. It is likely that this percentage will increase once molecular phylogenies become more analytically minable for traces of rate change, and calibration sources become more available.
IV Discussion
1 General trends of publication and authorship on historical biogeography
Throughout the analyzed period, the JB has shown a clear tendency to increase the number of contributions per volume, which seems to indicate that biogeography as a whole is an expanding research field. However, it is interesting to highlight the fact that historical biogeographical contributions have shown a more marked increase. This growth occurred just when the discipline is undergoing a period of scientific revolution (Crisci, 2001), with an increase in approaches and methodologies (see Crisci et al., 2003; Posadas et al., 2006). According to our data, in this study period most approaches tended to increase in number of contributions. However, one of the issues linked to the growth of historical biogeography is the extraordinary increase in the number of phylogeographical contributions. Contributions using paleoenvironmental reconstructions have also markedly increased in number. Comments on the notable increase in phylogeographical papers will be reserved for the discussion of ‘approaches and techniques’ below.
Regarding paleoenvironmental reconstruction papers, in our opinion their sharp increase has no simple explanation. The scope of this analysis did not allow us to determine if the aforementioned increase represented a general trend in geosciences, a shift within geosciences to paleoenvironmental questions, or a change in the strategy of publication by the geoscientist community. In recent years, there has been an attempt by paleontologists to expand the boundaries of their discipline in search of a more fluid and enriching dialogue with biologists. Historical biogeography is an especially appropriate discipline for opening a dialogue between paleontologists and neontologists, since its evolution has been strongly influenced by the paradigm of plate tectonics in the geosciences. Notwithstanding, it is interesting to note that 70% of the paleoenvironmental reconstruction papers published in the JB are focused on analyzing the dynamic of paleoclimatic changes during Quaternary times. Suggestively, the increase in this kind of paper coincides with the boom of phylogeography, whose explanations are focused on the same period of Earth history. Finally, another fact that converges with increasing papers on the Quaternary paleoenvironments and phylogeography is the issue of global climate change. Indeed, from the early 1990s, climate change (and particularly global warming) began to occupy an important place in the scientific agenda of the developed countries. The need to better understand the causes that led to climate changes resulted in an increase in the number of studies designed to elucidate the reasons for climate variations during the past, and especially during the Quaternary. These studies also benefited from the development of more sophisticated and powerful methodologies for better and more precise absolute dating and paleoclimatic estimates from different proxies (e.g. Adams et al., 1999; Ehlers and Gibbard, 2007; MacDonald et al., 2008; Mayewski et al., 2004; Meadows, 2012; Ramsey, 2008; Schreve and Candy, 2010; Wanner et al, 2008; Woodroffe and Murray-Wallace, 2012). In our opinion, this notable convergence of circumstances could explain, at least in part, the increase in the number of papers on paleoenvironmental reconstructions observed in JB.
The increasing percentage of papers with four or more authors, accompanied by the decrease in the percentage of papers with a single author, could be an indicator of increased cooperation/collaboration in historical biogeographical work. This pattern is not exclusive to historical biogeography, because a similar pattern can be found in most of the contemporary scientific journals. The increasing specialization of researchers, the emergence of more complex techniques and methods to address problems, and the presence of multidisciplinary teams in research institutions explain in part the growth of co-authored papers (e.g. Etzkowitz and Kemelgor, 1998; Hicks and Katz, 1996; Knorr-Cetina, 1998; Rogers, 2000). A question to discuss in the future is whether the cooperation is intra- or inter-institutional, national or international, and whether it is more frequent in one or other approaches (e.g. Bozeman and Corley, 2004). On the other hand, several studies have demonstrated that scientific collaboration has very significant impacts on productivity, citation rates, and, consequently, on the probability of promotion in the academic career (e.g. De Solla Price and Beaver, 1966; Figg et al., 2006; Lee and Bozeman, 2005; Noruzi, 2008, 2009; Pfirman et al., 2008; Reskin, 1977). Consequently, it is also possible that this increase in collaboration could be biased, at least in part, by external reasons.
European and North American authors are in the forefront of historical biogeographical research, accounting for 61% of the authorships of the analyzed papers. The greatest number of authorships (328) was from the USA, more than tripling the number of those from the UK (102), the second country in number of contributing authors. The first three countries which greatly contributed with authorships are English-speaking (i.e. USA, UK, and Australia). The fourth and sixth countries in authorship rank were Spanish-speaking (Spain and Mexico, respectively).
Posadas and Donato (2007: 30) noted that Latin American and Caribbean authors have made important contributions to the JB, positioning them almost at the same number as European and North American authors. However, this result does not seem to be confirmed by our analysis. When a greater span is considered, Latin American and Caribbean authors represented only 12% of the contributing authors, compared with 43% for Europe and 26% for North America. This difference is mainly the result of the sharp increase in European and North American authors since 2007 (see Figure 4). This increase in the number of European authors could be explained by three reasons: (1) the aforementioned global trend to increase the number of authors per paper (especially after 2004; see Figure 2); (2) the sharp increase in papers published by Europeans, especially in phylogeography, molecular biogeography, and paleoenvironmental reconstructions; and (3) the increasing number of European countries who publish in the JB. For North American (USA and Canada) authors, the strong increase in the number of authors could be related to the first two aforementioned reasons for European ones (except for paleoenvironmental reconstructions). On the other hand, the global trend to increase the number of authors per paper also involves Latin American and Caribbean, but in this region a significant increase in the number of published papers, as seen for European authors, is not evident. Additionally, only sporadic participation of authors from other LAC countries other than the leading three (i.e. Brazil, Mexico, and Argentina) are observed (Figure 5).
2 Internal trends of historical biogeography
a Habitats, areas, taxa, and events
We see a strong trend favoring works devoted to terrestrial habitats. The frequency of papers dealing with different habitats (i.e. 75% terrestrial, 12% freshwater, and 13% marine habitats) does not agree with the distribution of landmasses and waters over the Earth’s surface, because most of the Earth’s surface (70%) is covered with water, and the remaining 30% is taken up by the continental landmasses. On the other hand, the relationships among papers dealing with freshwater habitats versus papers dealing with marine habitats do not agree with the relationship between oceans (97%) and freshwaters (3%, and only 0.3% corresponding to surface waters) over the Earth’s surface. A possible explanation for this particular distribution of papers is that it is strongly biased by the degree of difficulty involving access to different habitats, as well as to the smaller average cost of accomplishing taxonomic works in continental environments (including freshwater) relative to marine environments. An alternative explanation for this pattern is related to the distribution of species diversity on continents (including freshwater habitats) and oceans. Of catalogued species, 87% inhabit continental habitats and the remaining 13% inhabit marine ones (calculated from Mora et al., 2011), that is almost the same proportion of historical biogeographical analysis devoted to each kind of habitat. Thus, the study of different habitats seems to be more influenced by taxon diversity than by the surface covered by each habitat.
Regarding continental regions analyzed, the distribution of papers dealing with the main continental biogeographical regions could be related to the geographic provenance of the biogeographers. In fact, most of the authors are European, followed by North American, inhabiting the northern regions analyzed in most papers. However, northern regions (i.e. Holarctic, Palearctic, Nearctic, and Oriental) represent the 55% of the papers devoted to one region, while these areas host 74% of total authorships (i.e. North American, European, and Asian authors). Conversely, the southern areas (Neotropical, Andean, Australasian, Cape, and Ethiopic regions) represent 41% of papers devoted to a single region and include 27% of authors. Thus, for example, of those papers focused on the Neotropical region, first authors work in LAC in 52% of the papers, while 42% of the papers have either North American or European first authors. Similarly, papers dealing with the Ethiopic region have an African first author in just 32% of the cases, while the remaining 68% have European or North American first authors. On the other hand, papers related to the Palearctic region have a European (83%) or Asian (12%) as first author in 95% of the cases, and those related to the Nearctic region have a North American first author in 93% of the cases. The aforementioned differences in the first author’s place of residence and the biogeographical regions analyzed could be related to the number of researchers and the expenditure in science and technology, which is greater in Europe and North America.
Concerning taxonomic groups analyzed at the rank of kingdoms, there appears to be a bias favoring the analysis of plant taxa related to the part of biodiversity that they represent, because plants are considered in 35% of the papers, while this kingdom represents just 16% of the catalogued species on Earth (Mora et al., 2011). Also, animal taxa are explored in 59% of the cases, while they represent 77% of the catalogued species (Mora et al., 2011). Within those papers dealing with plant taxa, there is a bias favoring gymnosperms, because they are considered in 2% of the papers dealing with plants while this group represents only the 0.29% of embriophytes (Crepet and Niklas, 2009). Angiosperms are considered in 83% of the papers related to plant taxa, a number similar to the 89% of embriophytes that they represent (Crepet and Niklas, 2009). Similarly, diversity within animals shows a strong bias regarding animal taxa analyzed in historical biogeographical papers. That is, the distribution of historical biogeographical papers does not reflect the real diversity of these phyla, because megadiverse animal groups such as arthropods (particularly insects), nematodes, and marine invertebrates are underrepresented. In fact, chordates represent 5% of the known living species, arthropods (e.g. insects, spiders, scorpions, and crustaceans) 89%, and molluscs 7% (this is the only phylum in which the percentage of biogeographical papers corresponds to the percentage of living species). Contrarily, ‘flagship’ groups like mammals and birds are also overrepresented in the papers dealing with historical biogeography. In fact, mammals only represent 9% of living vertebrate species, with birds at 16%, reptiles 15%, amphibians 10%, and fishes 50%. As with the molluscs, the percentage of biogeographical papers dealing with amphibians curiously matches the percentage of known living amphibian species (IUCN, 2011). Furthermore, the percentage of papers devoted to higher taxonomic groups correlates better with ‘the inefficiently distributed’ (May, 2010) labor force in taxonomy than with taxonomic diversity. According to May (2010), one-third of the taxonomy labor force is devoted to vertebrates, one-third to plants, and the remaining third to other animals. These proportions match the distribution of historical biogeographical papers among higher taxa (31% chordates, 35% plants, and 26% for animals other than vertebrates).
b Approaches and techniques
The theoretical versus empirical contributions are unbalanced for such an active discipline. It is interesting to note that of the 12 approaches analyzed here cladistic biogeography had the highest percentage of papers dealing with theoretical questions. This is not surprising, because this approach has continuously debated its theoretical framework and methodologies since its origin (see Crisci et al., 2003, and literature cited therein).
The origin of disjunct distributions has been a classical question in historical biogeography. From Darwin’s times until the second half of the 20th century, dispersal was the dominant answer to this question (Crisci and Katinas, 2009; de Queiroz, 2005). Both the acceptance of continental drift as a paradigm of geosciences and the spreading of cladistics as a paradigm in systematics in the 1960s and 1970s supported the idea that disjunct distributions could be explained by vicariance (Crisci and Katinas, 2009; de Queiroz, 2005). At the beginning of the 21st century, de Queiroz (2005) stated that there is a resurrection of trans-oceanic dispersal as an explanation of disjunct distribution based on molecular dating of lineage divergences, which favors dispersals over tectonic vicariances (but see Graur and Martin, 2004, and Heads, 2005, for a critical appraisal of molecular clock dating). An interesting finding of our analysis is the great number of papers using dispersalist and vicariant explanations, not as an alternative but jointly. It seems that this classical binary opposition in historical biogeography (Morrone and Crisci, 1995) is reaching an end. A similar conclusion was postulated by Riddle et al. (2008), who proposed that the availability of molecular phylogenies has provided biogeographers with evidence supporting that dispersal has played a role at least equivalent to the vicariance in the history of distribution of biodiversity. Regarding phylogeography, Avise (2000: 9) states that ‘research publications employing the word phylogeography in the title or as an index term have roughly doubled during each successive two-year interval since 1987’. A similar trend was observed for Riddle and Hafner (2004). According to our results, this trend is evident in the JB especially from 2004, and it continued and even increased from 2005 to 2010. Also, a cursory search of the Wiley-Blackwell homepage for papers using phylogeograph* in the title or keywords between 1990 and 1997 and between 1998 and 2010 confirms this trend. In the first interval we recovered only 14 articles, while in the second we recovered 487. Furthermore, Avise (2000: 9) states that ‘these articles represent only the small tip of the iceberg because many more studies have dealt with the topic implicitly though not by name’. We could confirm this assertion, as our search for the period 1998–2010 yielded 70 papers published in the JB that used phylogeograph* in the title or keywords, while our analysis in the same period revealed 195 contributions dealing with phylogeography.
The great surge in phylogeographical papers seems to lead the evolution of historical biogeography over the last years. The increase in the number of historical biogeographical contributions per year has accompanied the growth of phylogeographical papers (see Figures 1 and 6). Crisci et al. (2003) have postulated that molecular systematics is one of the external forces that are shaping biogeography. Crisci (2006) stated that biological systematics has entered into a molecular age, resulting in an unmatched revolution of the discipline. Our results indicate that historical biogeography is following biological systematics, and has entered its own ‘molecular age’ and it is already firmly entrenched in that age, since 47% of the analyzed papers utilized phylogeography and molecular biogeography and many papers classified under different approaches are based on molecular data. Also, the number of contributions using both aforementioned approaches increased greatly during the analyzed period. This is especially evident from the year 2004 in the JB (Figure 6). Furthermore, Riddle et al. (2008: 174) have postulated that ‘historical biogeography owes its renewed vitality to the molecular genetics revolution in systematic and population genetics’. Also, the molecular revolution has served to revitalize old arenas in biogeography and to generate new areas of research (Mantooth and Riddle, 2011), enriching the field of historical biogeography.
Arbogast and Kenagy (2001: 819) suggested that phylogeography ‘permits investigation of biogeographical questions on spatial and temporal scales that are smaller than those typically addressed with other approaches’ and that historical biogeography should continue to be a broadly integrative discipline, and, as such, it should include the perspective of phylogeography (Arbogast and Kenagy, 2001). The fact that phylogeography is the main force which is driving historical biogeographical research in recent years is reflected, for example, in the increase in papers dealing with infraspecific taxa and in those that use molecular clocks. Thus, the taxonomic scale is showing increasing representation of those papers focused on a single species or a few closely related species (from 1998 to 2003 contributions dealing with questions at infraspecific taxonomic hierarchy represent only 12% of the empirical papers published, while from 2004 to 2010 they represent 39% of the empirical papers published). Accordingly, the geographic scale would be reduced to a local or at most regional scale, and the timescale would be focused on a few million years. It is possible that if this trend continues historical biogeography would face a reduction of the taxonomic, geographical, and temporal analytical scales. A balance is required to avoid the rise of a new binary opposition in historical biogeography: phylogeography versus the rest of the approaches. Alternatively, it could be said that the rise of phylogeography has expanded the spatial scale of historical biogeography by incorporating Pleistocene in the scene of the discipline. Also, as Crisci and Katinas (2009) point out, some of the binary opposites in biogeography have been shown to be complementary rather than antagonistic approaches to the study of biogeography, and this is certainly the case of phylogeography regarding the other approaches. Many authors (e.g. Lomolino et al., 2010; Riddle, 2011; Riddle and Hafner, 2004, 2006; Whittaker et al., 2005) have discussed the value that the surge of phylogeography has provided in a growing relevance of historical biogeography in ecology, evolutionary biology, and conservation biology.
3 Historical biogeography: past, present and future
Crisci (2001) identified external and internal forces shaping historical biogeography. Global tectonics as the dominant paradigm in geosciences, phylogeny as the basic language of comparative biology, molecular systematics as a new window onto nature, and the biologist's perception of biogeography were determined to be the external forces which influenced historical biogeography. The proliferation of competing articulation and recourse to philosophy and the debate over fundamentals were determined to be the internal forces. Ten years later, we can assert that these forces have indelibly marked historical biogeography and that some of them prominently influence the discipline which will mark its future evolution.
The influence of global tectonics on historical biogeography was so huge that current biogeography is strongly linked to paleogeography (as evidenced, for example, by the increase in paleoenviromental reconstruction papers published in JB, or the papers using geological or geobiotic frameworks; see, for example, Erkens et al., 2009). Also, dispersal and vicariance have become alternative, even concomitant, and not opposite explanations for disjunct distributions, and both of them are tested regarding time and changing geography. The use of phylogenies has become common among historical biogeographers and many approaches are based on phylogenies. Also, methodological developments in phylogenetic systematics have been incorporated into historical biogeography (e.g. Bayes-DIVA by Nylander et al., 2008; PAE by Rosen, 1988). Furthermore, historical information encoded on phylogenies has been seen as a bridge for the gap between ecological and historical biogeography (Crisci et al., 2003; Wiens and Donoghue, 2004).
The influence of the molecular revolution in systematic and population genetics on biogeography is perhaps the strongest force that has shaped in the near past and will continue to shape historical biogeography in the near future. From phylogeography (Avise, 1992) to molecular biogeography (sensu Mantooth and Riddle, 2011), resolving the binary opposite between vicariance and dispersal explanations, introducing timing through molecular clocks, allowing the development of methods based on phylogenetic branch lengths, expanding the interest of historical biogeography to ecologist, evolutionary, and conservation biologists, giving insights into the future pattern of biodiversity distribution, molecular systematics, and populations genetics are modeling historical biogeography to the point that the discipline appears to be firmly entrenched in a molecular age.
From its origins, biogeography had been viewed as a subproduct of systematic biology. Crisci et al. (2003) stated that biogeography as a whole was perceived as an oddity by a vast majority of biologists. The statements of Nelson (1978), ‘Biogeography is a strange discipline. In general, there are not institutes of biogeography; there are not departments of it … It seems to have few authoritive spokesmen’, reflect this perception (Crisci et al., 2003). Today, biogeography is a growing discipline with its own journals, departments, and authoritative spokesmen. Biogeography has assumed its own place in the biological sciences and has started to take its place in conservation sciences (Richardson and Whittaker, 2010; Whittaker et al., 2005). Through our analysis, we have identified other forces that would be acting on historical biogeography – for example, the geographical distribution of researchers and resources for research and development throughout the world (Table 5) and the changes of research and development evaluation systems. These factors influence the discipline in several ways. For instance, there is a bias regarding regions analyzed, which correlates with authors’ places of residence and the consequent bias regarding taxon analyzed. Also, the increasing availability and accessibility of distributional data of taxa through the database systems is allowing the analysis and testing of existing biogeographical regionalization schemes, as well as the proposal of new schemes based on highly accurate data. This is reflected in the increase in papers corresponding to the biota similarity and PAE approaches (Figure 6).
Research and development statistics and number of authorships for North America (NA), Europe (EU), and Latin America and the Caribbean (LAC) regions. Values were calculated for each continent as the mean values for the countries with more authorships (NA = Canada and United States; EU = United Kingdom, Spain, Germany, and France; and LAC = Mexico, Brazil, and Argentina) from 1998 to 2009 based on data available from RICYT (2011; http://www.ricyt.org) and EuroStat (2011; http://epp.eurostat.ec.europa.eu). Researchers FTE = Researchers Full Time Equivalent; ES&T as % of GDP = Expenditure in Science and Technology as a percentage of the Gross Domestic Product; and ES&T in million dollars (PPP) = Expenditure in Science and Technology in current millions of dollars expressed in power payment parity.

The proliferation of competing articulation and recourse to philosophy and the debate over fundamentals are still internal forces that continue modeling the discipline, as is evidenced in the papers discussing theoretical aspects of the discipline and the emergence of new forms to analyze historical biogeographical questions (e.g. the increasing number of papers classified under ‘other approaches’ and those that combine methods from different approaches) (Figure 7).
V Conclusions
Historical biogeography is a research field that has increased its representation throughout the analyzed period (from about 15% of papers published in JB in 1998 to 40% in 2010; Figure 1). The emergence of new approaches, the combination of techniques, the resolution of some binary opposites and the arising of new ones, and the increase in questions indicates that historical biogeography is an active discipline.
Our results show: (1) an increase in historical biogeographical publications; (2) native English-speaker researchers, especially those from the USA, lead authorship in the field; (3) collaboration has become more frequent resulting in a greater number of authors per paper; (4) phylogeography has become the most represented approach in historical biogeographical publications; (5) as a consequence of that, research on short temporal scales and infraspecific taxa or closely related species has become more common in historical biogeography in the last years; (6) there is a major tendency to address historical biogeographical questions to terrestrial habitats rather than to aquatic ones; (7) the binary opposition between dispersal and vicariance (Crisci et al., 2003) seems to be resolved, because 45% of those papers discussing historical events deal jointly with dispersal and vicariance explanations; (8) paleoenvironmental reconstruction papers with a profile oriented to historical biogeographical studies are increasing their representation.
Some of the future challenges that historical biogeography faces could be summarized as follows: (1) to increase the study of those taxa underrepresented according to the part of the biodiversity they represent; (2) to balance the amount of work devoted to different biogeographical regions; (3) to increase biogeographical knowledge of marine and freshwater habitats; (4) to maintain the diversity of approaches, preventing the reduction of time, spatial, and taxonomic scales addressed by the discipline; and (5) to continue integrating historical biogeography along with other sources of information from other disciplines (e.g. ecology, paleontology, geology, isotope chemistry, remote sensing) into a richer context for explaining past, present, and future patterns of biodiversity on Earth.
Footnotes
Acknowledgements
Jorge Crisci, Liliana Katinas, and Sandra Miguel critically read a preliminary version of the manuscript. Jeannie Miller improved the English and made usefull comments about the final manuscript. Two anonymous reviewers made valuable comments and suggestions. Our research is financed by PIP 0729 (CONICET) and N662 (UNLP).