
Abstract
James H. Brown’s “Mammals on mountaintops: nonequilibrium insular biogeography,” published in 1971 in The American Naturalist, documented distributional patterns of small mammal species in the mountaintop islands of the Great Basin, USA. Distributional patterns suggested that this island-like system was not in equilibrium and represented some of the first evidence contradicting the seminal Theory of Island Biogeography. Brown’s findings suggested that ecological and historical mechanisms were integral to community assembly and maintenance in island-like systems, broadening the focus of research related to biogeographical patterns in islands. The work further highlighted the importance of species traits on distributional patterns. Here, I review the paper and its contributions.
I Introduction
In 1971, James H. Brown’s manuscript, “Mammals on mountaintops: nonequilibrium insular biogeography,” was published in The American Naturalist. It was one of many published during an initial flurry of research to test MacArthur and Wilson’s seminal Theory of Island Biogeography (TIB, MacArthur and Wilson, 1963, 1967). TIB, one of the first conceptual and quantitative theories of community ecology, proposed that species richness represented a dynamic equilibrium between colonization and extinction rates, which are determined by both island size and isolation. The authors speculated that TIB should apply to other insular systems, including caves, lakes, and forest patches (MacArthur and Wilson, 1967). As a result, research emerged focusing on the degree to which other insular systems behaved similarly to oceanic islands, a question that remains fundamental in island biogeography (e.g., Itescu, 2019; Patiño et al., 2017). Prior to the publication of “Mammals on mountaintops,” communities of island systems (both oceanic and habitat) were found to be in equilibrium (e.g., Culver, 1970; Diamond, 1969; Simberloff and Wilson, 1969, 1970; Vuilleumier, 1970). Brown’s findings of non-equilibrium, one of the first major challenges to the nascent TIB, set the stage for development of novel concepts in insular biogeography, facilitated the expansion of core island biogeography theory (sensu Warren et al., 2015), and revived montane biogeography research (Lomilino, 2001). As a result, it emphasized the importance of island-like systems in developing theories for true islands (Itescu, 2019), and pivoted research toward ecological, evolutionary, and historical causes of non-equilibrium, bringing the theory in better alignment with reality (Lomolino and Davis, 1997). Here, I summarize the salient points Brown articulated and discuss the manuscript’s legacy and effects on the trajectory of biogeographical research.
II James H. Brown
Much of the following information was summarized from Maurer and Marquet (1997). James (Jim) H. Brown (Figure 1) was born in Ithaca, New York, USA in 1942. Jim attended Cornell for his undergraduate degree, conducting an honors thesis on convergent evolution in multi-armed starfish. In 1967 he completed his doctoral work at the University of Michigan, focusing on the physiological mechanisms of adaptation under the advisement of Emmett T. Hooper and the close mentorship of William Dawson. He moved into a postdoc and then began his faculty career at the University of California Los Angeles, where he met Robert MacArthur and developed an interest in community ecology and biogeography. During his distinguished career, Jim has spent time as a professor at the University of Utah, University of Arizona, and University of New Mexico. He is currently a Distinguished Professor of Biology Emeritus at the University of New Mexico.

James H. Brown (Reprinted with permission from the Ecological Society of America, Maurer and Marquet (1997)).
Jim’s contributions to the ecological and biogeographical dialogue have evolved both ecology and biogeography. In a seminal 1989 paper, he and Brian Maurer laid the foundations for a new field of ecology, macroecology (Brown and Maurer, 1989), integrating empirical and theoretic frameworks to focus on questions at larger spatial and temporal scales. He and his wife Astrid defined the rescue effect (Brown and Kodric-Brown, 1977), facilitating new approaches to island biogeography and conservation; and he, along with collaborators, produced a fundamental theory on metabolic scaling that garnered him the Ecological Society of America’s MacArthur Award (Brown et al., 2004). Guided by curiosity, Jim’s willingness to follow the evidence where it leads, emphasize collaboration, and ability to see the threads connecting disparate disciplines has shifted and broadened the fields of biogeography, ecology, and conservation. His legacy is woven into the fields he influenced, and propagated by the scientists he mentored.
III Disequilibrium dynamics
In “Mammals on mountaintops”, Brown investigated whether the small mammal communities of boreal mountaintop islands in the Great Basin, USA (Figure 2), were in equilibrium as predicted by TIB. Using an elegant and straightforward design, Brown determined small mammal community composition in each of 17 mountain islands, estimated the parameters of the species area curve, assessed whether species richness declined with distance from the nearest mainland or other factors associated with colonization rates, and evaluated whether distributions were associated with species traits. His surprising conclusion—that the boreal small mammal faunas of the montane islands of the Great Basin did not represent equilibria between recurrent rates of colonization and extinction—was informed by multiple lines of evidence: the slope of the species area curve (z = 0.43) indicated limited colonization; species richness was not negatively associated with isolation; and other colonization pathways (e.g., stepping-stone processes) were unsupported. These conclusions were bolstered by paleontological evidence that, at the time, showed colonizations had not occurred since the late Pleistocene, when climatic barriers were absent and boreal forest connected now discontinuous mountaintop islands. Although updated contemporary species distributions and novel paleontological evidence of colonizations (Grayson, 2006) suggest immigration played a more important role than Brown’s data originally indicated (e.g., Lawlor, 1998), his imprint on TIB and disequilibrium dynamics remain significant.

Wheeler Peak in the Snake Range, Nevada, USA. Note the band of coniferous forest above the sagebrush matrix (Public domain photo).
By emphasizing that past climatic change could produce non-equilibrium states, Brown illustrated the important role of historical events in defining biogeographical patterns and shifted the dialogue to relative rates of colonization and environmental change. In addition, he recognized a time lag between habitat fragmentation and extinctions, noting that, for a system with zero colonization—as he documented for Great Basin small mammals—equilibrium would only be reached “in geological time spans.” In so doing, he acknowledged the potential for static non-equilibrium communities (e.g., Whittaker, 2000) and laid the foundation for the concepts of relaxation (e.g., Diamond, 1972) and extinction debt (Tillman et al., 1994), where habitat fragmentation, climate change, or invasions (Malanson, 2008), cause species richness to “relax” to equilibrium via extinctions over time. Perhaps more importantly, the findings of disequilibrium helped facilitate a shift in ecological thinking. The standing ecological paradigm at the time held that ecological systems were at or trending toward equilibrium; Brown’s empirical findings of disequilibrium contributed to a new perspective that systems are dominated by disequilibrium and imbalance (e.g., Worster, 1989; Zimmerer, 1994; Zimmerer 2000).
Historical context and time-lags, arguably the most influential disequilibrium concepts generated by Brown (1971), have been heavily woven into current biogeographical research, particularly in areas of global change. Disequilibrium dynamics are apparent in some island-like systems impacted by climate change (e.g., Marta et al., 2019), as well as in fragmented areas resulting from anthropogenic land use change (e.g., Bolger et al., 2000). In addition, the threads of these concepts are visible throughout conservation ecology, from the development of conservation areas, determination of their areal extent, and assessments of their efficacy (Diamond, 1975; Figueiredo et al., 2019; May, 1975; Stegner et al., 2017).
IV Variation by species and trait
No less important were Brown’s observations of trait- and taxon-specific patterns. The distribution of Great Basin small mammals was influenced by ecological traits including body size, trophic level, and habitat specialization (Figure 3). Because these characteristics are associated with population size, Brown concluded that differential extinction rates were a likely cause of the patterns he observed. The influence of species traits on biogeographical patterns is an active area of research in biogeographical theory.
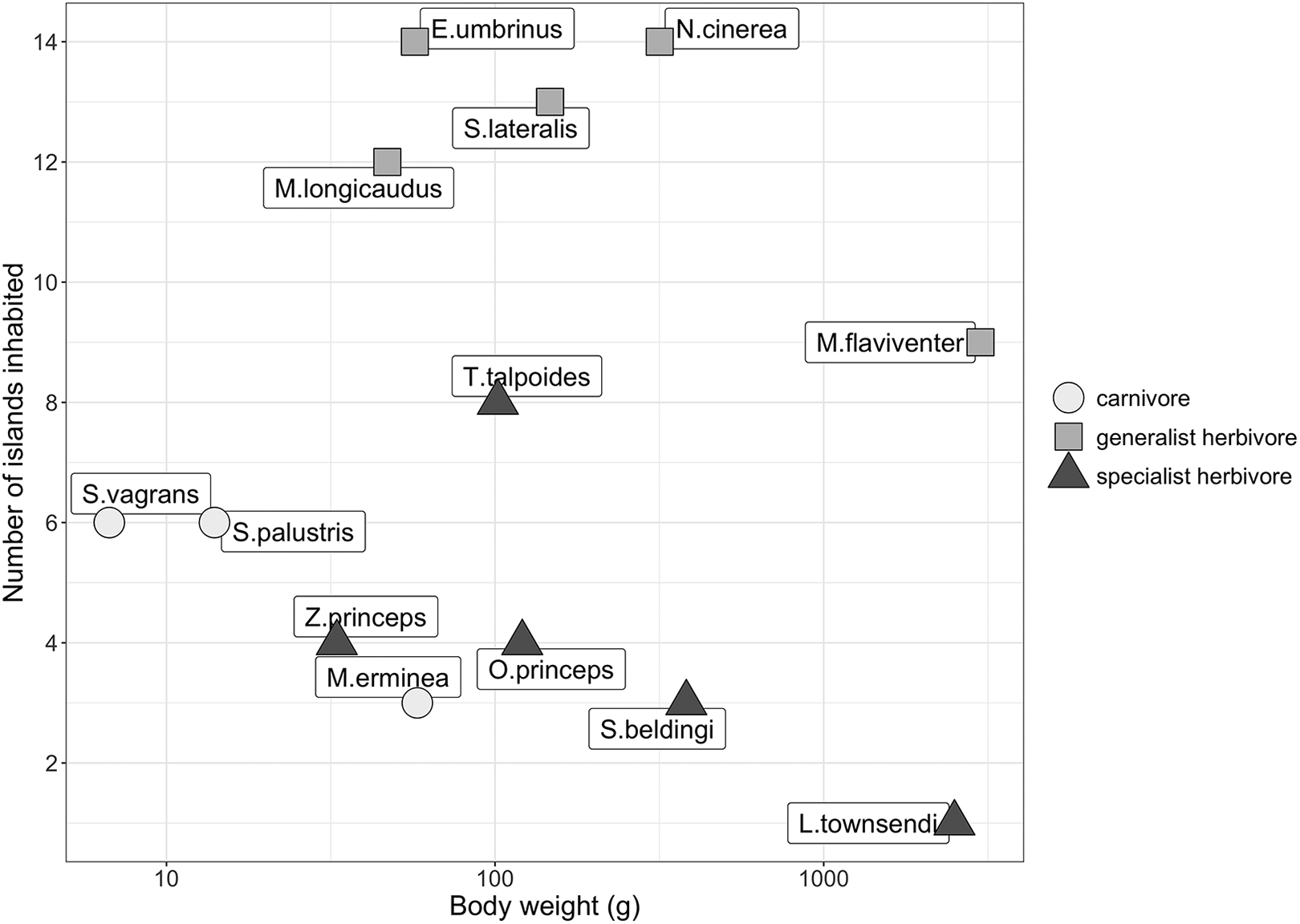
Recreation of Figure 3 in Brown (1971) from data presented therein. Trait-specific distributional patterns are apparent. These findings presaged much research into species specific responses and trait-based biogeography.
Brown acknowledged that dispersal ability of the focal taxon influenced colonization and extinction dynamics; although matrix permeability for small, non-volant (those who cannot fly) mammals was low, it may not be for other taxa. He later provided empirical support by comparing the avifauna and small mammal biogeography in the Great Basin, finding that the avifauna were in equilibrium, and supporting his original findings that the small mammals were not (Brown, 1978). His findings highlighted the importance of ecological traits (i.e., dispersal ability) in determining matrix permeability: matrixes are not equally permeable to all taxa. The influence of ecological traits on and taxa-specific differences of distributional patterns is at the forefront of biogeographical research because of their effects on colonization, extinction, and speciation (Whittaker et al., 2008; Patiño et al., 2017).
V Conclusions
Although TIB provides a useful neutral model for island biogeography, it has been the deviations from its expectations that have contributed most to our understanding (Warren et al., 2015; Whittaker et al., 2017). Brown’s work highlighted disequilibrium dynamics and began to shift TIB from near-paradigm status to a heuristic useful as a guide for biogeographers to better define and test the complexities of insular biogeography. Indeed, “Mammals on mountaintops” and Brown’s other studies on the insular fauna of the Great Basin (Brown, 1971, 1978) are most often viewed not as a repudiation of TIB, but as a lynchpin broadening its focus, adding nuance and realism to TIB as a neutral theory. The legacy of Brown’s work lies in its emphasis on historical and ecological contributions to species distributions, and it continues to shape the research trajectory of insular biogeography.
Footnotes
Acknowledgements
I am grateful to GP Malanson and DR Butler for their suggestions and input.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.