
Abstract
Objective
This study aimed to identify Nuclear-activated miRNA (NAmiRNA) and their target genes linked to the prognosis of Lung adenocarcinoma tumor (LUAD), and provide novel diagnostic and therapeutic targets for this devastating disease.
Methods
Putative target (Messenger RNA)mRNA of the prognostic (MicroRNA)miRNA was obtained by interesting the differentially expressed mRNAs with predicted targets from targetscan database. Functional validation experiments were conducted utilizing LUAD cell lines to corroborate the interactions between the identified miRNA and their target mRNAs. Collect cancer and adjacent tissues from patients to detect the expression of target genes. The effects of miRNA-target gene interactions on LUAD cell growth were investigated by (Cell Counting Kit-8)CCK8, colony formation assay and flow cytometry.
Results
Our findings revealed that both miR-31 and its target gene, ZWINT, were considerably overexpressed in LUAD individuals. The results of qPCR and WB indicated that miR-31 effectively upregulated the expression of (Zeste White 10 interactor)ZWINT. The transfection with miR-31mimic enhanced the activity of LUAD cells and promoted colony growth. Knockdown of ZWINT expression resulted in (G2 to mitosis)G2/M phase cell arrest, concurrent with a decrease in Cyclin B1 and (Cyclin-Dependent Kinase 1)CDK1 proteins.
Conclusion
(nuclear activating mirna-31)NAmiRNA-31 and ZWINT are upregulated in LUAD, suggest their potential as biomarkers for the unfavorable prognosis.
Introduction
Lung cancer continues to be one of the primary contributors to cancer-associated mortality globally. 1 Histologically, lung cancer is categorized into samll cell lung cancer (15% of all instances) and non-small cell lung cancer (NSCLC) (85% of all instances). Within NSCLC, three subtypes are acknowledged: adenocarcinoma, squamous carcinoma and large cell carcinoma. 2 Particularly, LUAD stands as the predominant subtype, representing about 40% of all lung cancer incidents. 3 Despite recent therapeutic advancements in LUAD, blocking inhibitory receptors and cytokines, immunotherapy, chemotherapy, and radiotherapy, the outlook remains disconcertingly bleak. The advent of “precision medicine” has underscored the potential of molecular pathology detection methods and targeted therapies in enhancing the overall survival (OS) rates of LUAD population. 4 Molecules like epidermal growth factor receptor, anaplastic lymphoma kinase, FNDC4, LMNB2, recognized as pathological detection hallmarks and therapeutic targets for adenocarcinoma, nonetheless, have proven beneficial solely for a minor fraction of LUAD patients.5,6 Hence, the pursuit of novel, more efficacious indicators for the molecular pathological diagnosis and prognostic evaluation of LUAD patients is of paramount importance.
MicroRNA (miRNA), a class of endogenous non-coding RNAs, exhibits substantial regulatory functions, with its aberrant expression often being indicative of prognostic significance across a range of physiological and pathological processes. 7 Notably, cell cycle-associated miRNAs have been implicated in LUAD. 8 A class of miRNAs, denoted as nucleus-enriched abundant microRNA (NAmiRNA) and present in the nucleus, are crucial in gene activation through enhancer activation.9,10 NAmiRNAs have been observed to influence tumor metastasis and proliferation, as evidenced by miR-1246's role in fostering osteosarcoma metastasis and miR-339's involvement in promoting breast cancer cell proliferation and migration. 9 The expression of miRNA-144-5p was significantly down-regulated in LUAD, but overexpression of it repressed proliferation and spheroidization, and promoted apoptosis of LUAD cells. 11 In LUAD tissues, the expression of miR-21-5p was found to be significantly higher than in normal tissues. MiR-31 is another significant oncogenic miRNA. MiR-31-5p has been found to be significantly upregulated in LUAD tissues and cell lines. Its overexpression has been shown to promote cell proliferation and migration while inhibiting apoptosis. 12 Despite these insights, the understanding of the contribution of NAmiRNA to LUAD remains scant, necessitating further investigation.
ZWINT regulates kinetotubule attachment and spindle assembly checkpoints. Its overexpression existed in various human malignancies, including lung cancer and hepatocellular carcinoma, making it a potential biomarker for cancer.13,14 The knockdown of ZWINT inhibits proliferation, migration, invasion, and colony-forming ability of pancreatic cancer cells, along with increased apoptosis. 15 ZWINT gene was involved not only in controlling the cell cycle development in LUAD and LUSC but also in regulating processes such as cell division, chromosomal segregation, nucleoplasm, cell periphery, etc. The ZWINT gene has a stronger influence on the development and stability of cell membranes in LUAD and a greater influence on the extracellular matrix and some immunomodulatory responses.Specifically, ZWINT expression was negatively associated with the activities of the T cell receptor signaling pathway, B cell receptor signaling pathway, toll-like receptor signaling pathway, and cytokine-cytokine receptor interactions.In addition, ZWINT expression was positively related to several cell death pathways in patients with KIRC and THCA.ZWINT expression was negatively related to several cell death pathways in patients with CESC, ESCA, and GBM. 16 Moreover, ZWINT has been found to modulate the expression of several cyclins, such as PCNA, CyclinB1, Cdc25C and CDK1. 17 Dysregulation of cyclin expression is linked to the abnormal cancer cell growth. 18 CDK1, a crucial regulator of the cell cycle, is a contributor to G2/M phase transition during mitosis. 19 Elevated CDK1 expressionhas been associated with the development and poor prognosis of various cancers, including colorectal cancer, liver cancer, lung cancer, and pancreatic cancer. 20 Nevertheless, the mechanism underlying the interplay between ZWINT and CDK1 in LUAD remains obscure.
A thorough bioinformatics ananlysis revealed the association between NAmiRNA-31, its target gene ZWINT, and LUAD progression. Then the hypothesis proposing the involvement of the miR-31/ZWINT/CDK1 pathway in promoting LUAD progression was put forth. Subsequent validation through clinical investigations and in vitro cell experiments further substantiated this hypothesis, enhancing our understanding of the underlying mechanisms driving LUAD pathogenesis and progression. Moreover, these findings hold promise for identifying potential therapeutic targets for clinical interventions aimed at improving the prognosis of LUAD patients.This study aimed to identify NAmiRNA and their target genes linked to the prognosis of LUAD, and provide novel diagnostic and therapeutic targets for this devastating disease.
Materials and methods
Differential expression gene and survival analysis
The TCGA-LUAD downloaded database (https://portal.gdc.cancer.gov/) was employed to retrieve the miRNA and mRNA count matrices from patients with lung adenocarcinoma and adjacent tissue carcinoma. After removing miRNA and mRNA data with counts < 0, differential analysis was implemented utilizing the the “edgeR” package in the R program. The results were visualized through the volcano plots and heat maps. Concurrently, OS data of TCGA-LUAD were obtained, and individual univariate Cox analysis was conducted on the miRNA and mRNA datasets to evaluate their association with lung cancer patient survival. The analysis results were incorporated into the differential analysis findings, leading to the identification of differentially expressed miRNA and mRNA that exhibited potential associations with survival outcomes. Subsequently, as per the median gene expression, the individuals were classified into high- and low-expression categories for comprehensive survival analysis.
mRNA screening for miRNA targeting
Targeted mRNAs of miRNAs were screened utilizing the targetscan repository (https://www.targetscan.org/). The correlation between miRNAs and genes exhibiting high differential expression and dismal outcome in LUAD was analyzed to identify positively correlated genes.
GSEA analysis
The LUAD patients from the TCGA-LUAD repository were stratified into high- and low-expression categories per to the median gene expression. Bayesian analysis was implemented using the “limma” package in R to derive the T-value for the entire genome. 21 Furthermore, pathway analysis was performed by the GSEA algorithm. 22
Specialmen collection
Collect tissues from patients diagnosed with LUAD from the First Affiliated Hospital of Harbin Medical University. The collected samples will be stored at a temperature of −80 °C. These studies have been approved by the ethics committee of the First Affiliated Hospital of Harbin Medical University Scientific Research, Article Ethics Review 202418. All participants were informed and consented.
Cell culture and transfection
The LUAD cell lines A549 and H1975 were procured from the Shanghai Institute of Biological Sciences, Chinese Academy of Sciences (Shanghai, China). The purchased cells were P4 generations, which were stably transmitted in the laboratory for two generations for subsequent experiments.These cells were raised in RPIM1640 medium fortified with 10% FBS and 1% penicillin-streptomycin at 37°C in a 5% CO2. A549 and H1975 cells were plated in 6-well plates and incubated until they reached about 80% confluency, the cell passage cycle was 2–3 days, in a ratio of 1:3. For experimental groups, transfection of miRNA-31-mimic, miRNA-31-inhibitor, miR-NC, sh-NC and sh-ZWINT was conducted, transfection uses mirnas or inhibitors at working concentrations of 50 nM. After transfection, cells were harvested 48 h later for subsequent detection.
qPCR
Total RNA was isolated from A549 and H1975 tilizing the Trizol reagent (Invitrogen, California, USA). Subsequently, cDNA synthesis was carried out employing the PrimeScipt RT kit (TaKaRa, Dalian, China). Real-time polymerase chain reaction (PCR) was implemented employing the SYBR Premium EX Taq kit (TaKaRa, Dalian, China). The expression results were analyzed employing the 2−△△Ct technique.
Western blotting
Total proteins were isolated utilizing RIPA buffer, and the protein content was detected applying a BCA kit (Thermo Fisher, Waltham, USA). The proteins were subsequently resolved by 12% SDS-PAGE and transferred onto polyvinylidene fluoride (PVDF) membranes. The membranes were sealed at room temperature for 1 h, and then raised overnight with the primary antibody at 4°C. The next day, the membrane were incubated with the secondary antibody at room temperature for 1 h and subsequently exposed. The grayscale values of the protein bands were calculated employing Image J.
Cell growth assay
To measure cell proliferation, the CCK8 assay was employed. Cells in the logarithmic growth phase were prepared as cell suspensions and added into 96-well plates (2000 cells/well), following the experimental groups for cultivation and treatment. After the designated processing, 10μl of CCK8 solution was introduced to every well and the plates were raised at room temperature for 1 h. The absorbance at 450 nm was then detected utilizing an enzyme-labeled apparatus.
Cell cycle analysis
To analyze the cell cycle, the processed cells were collected and further processed according to the instructions provided in the Cell Cycle Analysis Kit (BioVision, Inc). After processing the cells with the Cell Cycle Analysis Kit, the cell cycle was then evaluated applying FACScan flow cytometry from BD Biosciences. The obtained data from the flow cytometry analysis can be analyzed using the CellQuest software.
Statistical analysis
The data obtained from the experiments was analyzed using IBM SPSS 21.0 software(IBM, Armonk, NY, USA). To assess the significance of the observed differences, independent sample T-test and ANOVA method were employed. p < .05 indicates statistical significance. (*p < .05, **p < .01, ***p < .001, ****p < .0001).
Results
Analysis of miRNA overexpression and its prognostic correlation in LUAD
Differential analysis was implemented on the miRNA data obtained from 521 LUAD individuals and 46 adjacent tissues. The analysis identified 273 miRNAs with considerably higher expression and 117 miRNAs with considerably lower expression. Volcano maps and heat maps depicting the differentially expressed genes were generated (|log2FC| > 1, FDR < 0.05), with red indicating genes with considerably high expression, green indicating genes with considerably reduced expression, and black indicating genes with insignificant expression (Figure 1(a) and (b)). The total survival data of LUAD patients were collected, and univariate Cox analysis was conducted on the miRNA profiles. The findings from the analysis were combined with the results of differential analysis, which led to the identification of 35 miRNAs with significantly high expression ((p < .05) and HR > 1, Figure 1(c)), as well as 6 miRNAs with significantly low expression ((p < .05) and HR < 1, Figure 1(d)). Based on the median expression levels of these 41 miRNAs, the individuals were classified into high- and low-expression categories for OS analysis. These findings revealed high expression of miR-31 and miR-7974 was linked to poor prognosis(p = .02) (Figure 1(e) and (f)), while decreased levels of miR-99a was also linked to an unfavorable outcome(p = .003) (Figure 1(g)). Previous literature has suggested that the miR-17∼92 sites are cancer-associated enhancer regions. 9 Consequently, this study plans to conduct follow-up investigations specifically focusing on miR-31.
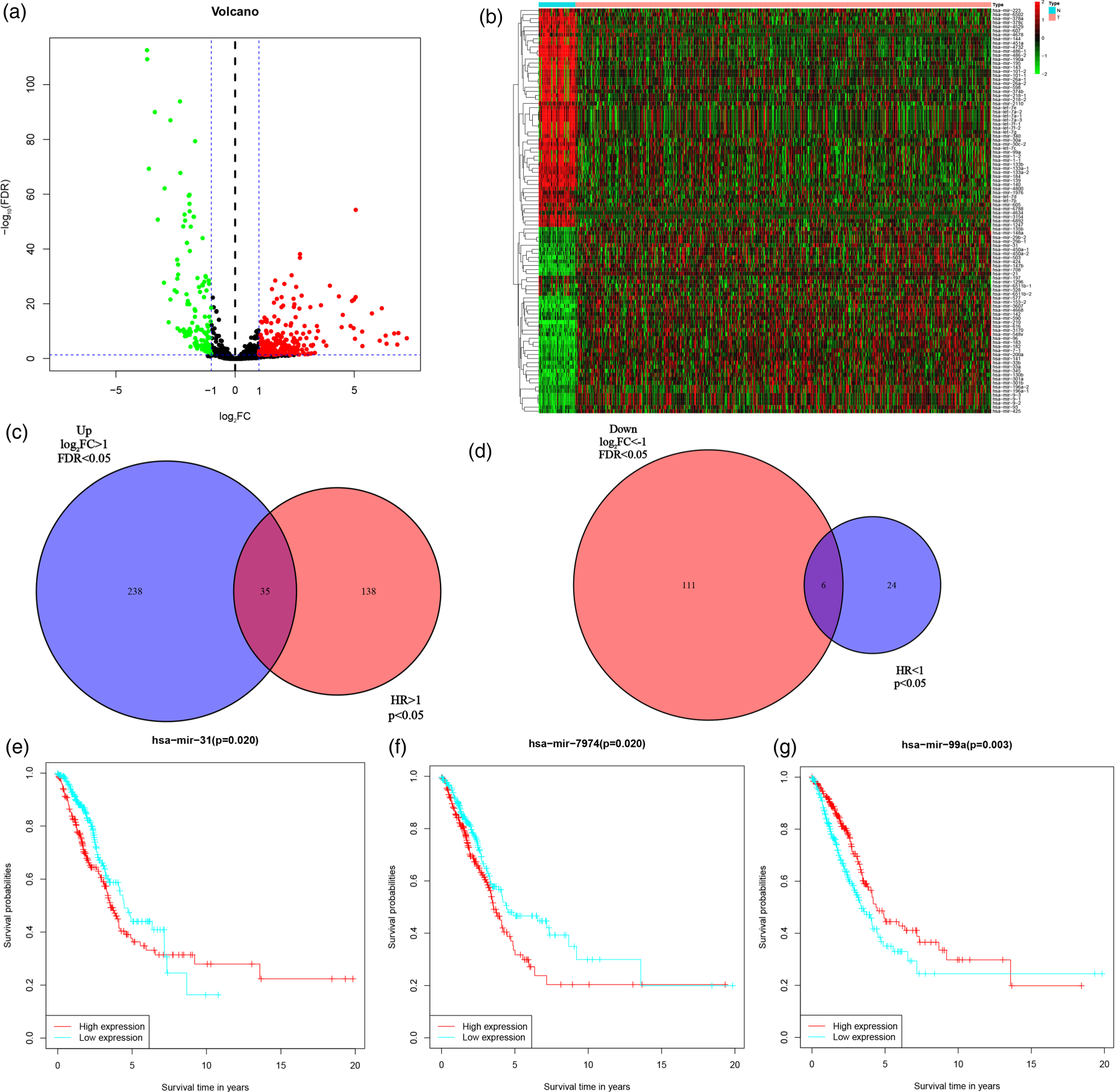
miRNA expression and its prognosis analysis. (a) Volcano map depicting the differentially expressed miRNA genes in LUAD patients and adjacent tissues; (b) Heat map illustrating the differentially expressed miRNA genes in LUAD patients and adjacent tissues; (c) Univariate Cox analysis of miRNA genes intersected with a Venn map highlighting differentially high-expressed genes; (d) Univariate Cox analysis of miRNA genes intersected with a Venn diagram highlighting differentially low-expressed genes; E: Relationship between miR-31 expression level and patient prognosis; (f) miR-7974 expression level and its impact on patient prognosis; G: miR-99a expression level and its impact on patient prognosis.
Differential analysis was conducted on mRNA data obtained from 541 LUAD patients and 59 adjacent tissues. The analysis identified 6439 considerably high-expressed and 939 considerably low-expressed genes. Volcano and heat maps displaying the differential expression patterns are presented in Figure 2(a) and (b) (|log2FC| > 2, FDR < 0.05, significantly high-expressed genes are red, significantly low-expressed genes are green, and insignificant genes are black). The total survival data of LUAD patients were downloaded, and univariate Cox analysis was conducted on mRNA profiles. The analysis results were then compared with the differential analysis results, yielding 637 significantly high expression genes (p < .05, HR > 1, Figure 2(c)) and 159 significantly low expression mRNA genes (p < .05, HR < 1, Figure 2(d)). Subsequently, per to the median expression of these 796 genes, the patients were classified into high- and low-expression sets for OS analysis. The analysis revealed that 246 high expression genes and 77 low expression genes were associated with a poor prognosis (Supplementary Tables 1 and 2).

mRNA expression analysis in LUAD. (a) Volcano map illustrating differentially expressed mRNA genes; (b) Heatmap displaying the mRNA differential expression patterns; (c) Single-factor Cox analysis of mRNA genes with a Venn diagram demonstrating the intersection of differentially high-expressed genes; (d) Single-factor Cox analysis of mRNA genes with a Venn diagram displaying the intersection of differential low-expressed genes.
2387 miR-31 targeted mRNA were identified from the targetscan database (Supplementary Table 3). A correlation analysis was conducted between miR-31 and genes that were highly expressed and associated with poor prognosis in the 246 LUAD samples mentioned earlier, resulting in 59 genes displaying statistical significance (p < .05) (Supplementary Table 4). By intersecting the 2387 target mRNA with the 59 genes, 6 target genes were detected, namely GOLM1, AGMAT, MCM4, GPR37, AURKA, and ZWINT (Figure 3(a)). Among these genes, ZWINT exhibited the highest correlation with miR-31 and a significant positive correlation (p = 1.1052e-04) (Figure 3(b)). qPCR detection showed that the expression levels of miR-31 and ZWINT were significantly higher in tumor issue than in normal tissue of patie (Figure 3(c)). Subsequently, A549 and H1975 LUAD cells were transfected with miR-31-mimic, miR-31-inhibitor and NC, and the effect of miR-31 on ZWINT expression was validated through qPCR and WB. The qPCR results demonstrated that in both cell types, the levels of miR-31 and ZWINT was considerably upregulated in the miR-31-mimic set compared to the NC set (Figure 3(d)). Conversely, the production of both genes was substantially downregulated in the miR-31-inhibitor set (Figure 3(e)). The WB results further confirmed that increased miR-31 protein production correlated with increased ZWINT expression, and vice versa. (Figure 3(f) to (g)). This evidence indicate that miR-31 positively regulates ZWINT levels in LUAD cells.
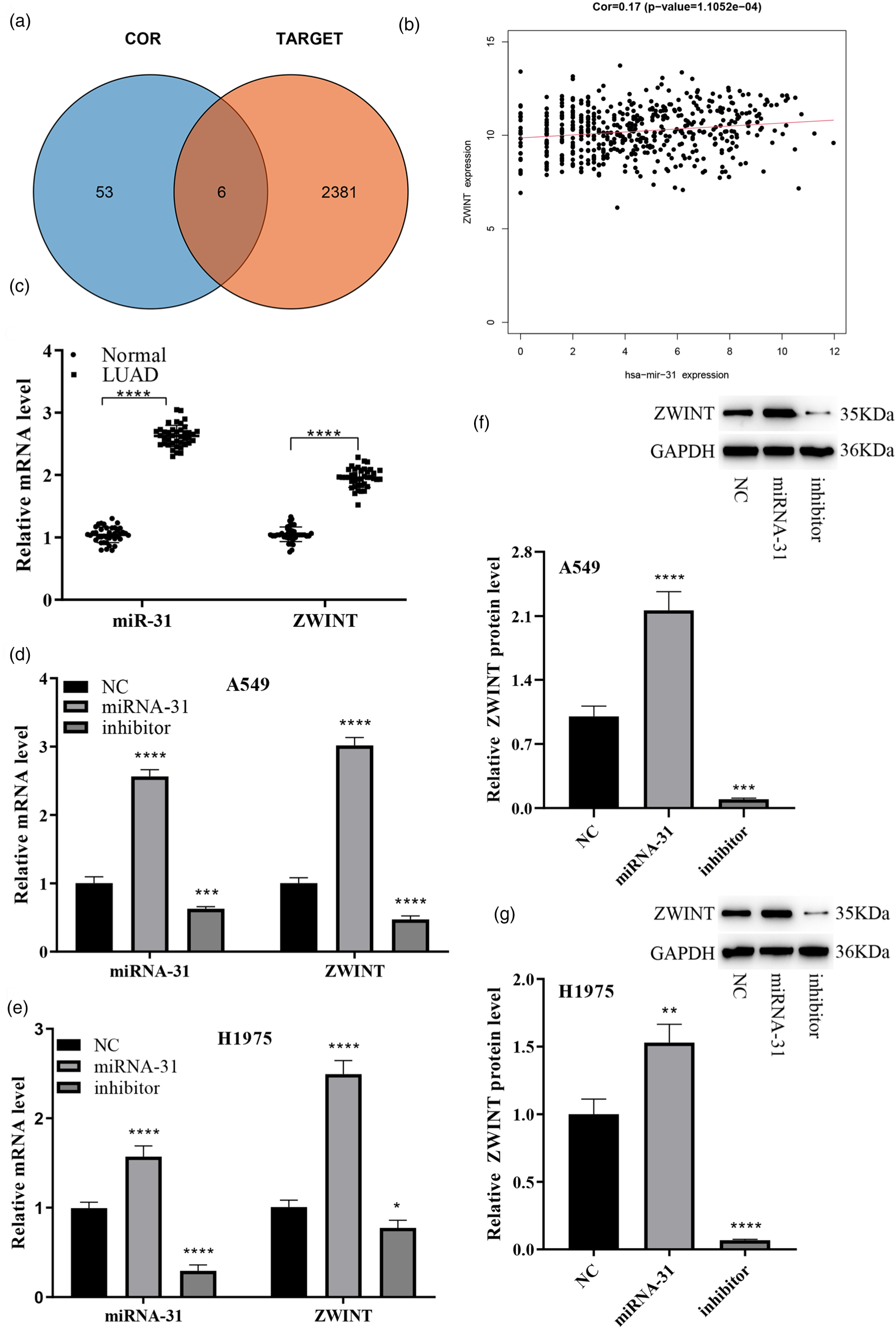
Analysis and validation of miR-31 positive regulation of ZWINT expression. (a) Venn diagram illustrating the intersection of miR-31 target mRNA and differentially expressed mRNAs in LUAD; (b) Correlation graph depicting the connection between miR-31 and ZWINT; (c) qPCR detection of miR-31 and ZWINT expression in tissues of lung adenocarcinoma patients; (d-e) qPCR demonstrating the impact of miR-31 on ZWINT production; (f-g) WB examination showing the influence of miR-31 on ZWINT expression.
The present research confirms the positive regulation of ZWINT transcription by miR-31, highlighting their significant overexpression in LUAD. Our findings lead us to conclude that miR-31 specifically activates ZWINT, exerting a profound impact on the LUAD cell growth. Initially, A549 and H1975 cells were transfected with miR-31-mimic and NC, followed by an assessment of cell proliferation through CCK8 and colony formation experiments. Our results demonstrated a significant increase in cell viability and colony count upon miR-31-mimic transfection, underscoring the function of elevated miR-31 production in enhancing cell growth (Figure 4(a) and (b)). Then we transfected cells with sh-NC and sh-ZWINT vectors and detected their proliferation capabilities. Both CCK8 and colony experiments revealed a substantial reduction in cell proliferation ability and colony count in the sh-ZWINT set relative to the sh-NC set (Figure 4(c) and (d)). This substantiates the inhibitory effect of ZWINT downregulation on cancer cell growth. Subsequently, we transfected cells with the sh-ZWINT vector and treated them with NC and miR-31-minic to investigate the interplay between the two factors in cell growth. The results evidenced a substantial rise in cell viability and colony count in the sh-ZWINT+miR-31-mimic group as compared to the sh-ZWINT+NC group (Figure 4(e) and (f)). This indicates that the augmented expression of miR-31 activates ZWINT expression, partially reversing the decline in cell proliferation caused by ZWINT knockdown.
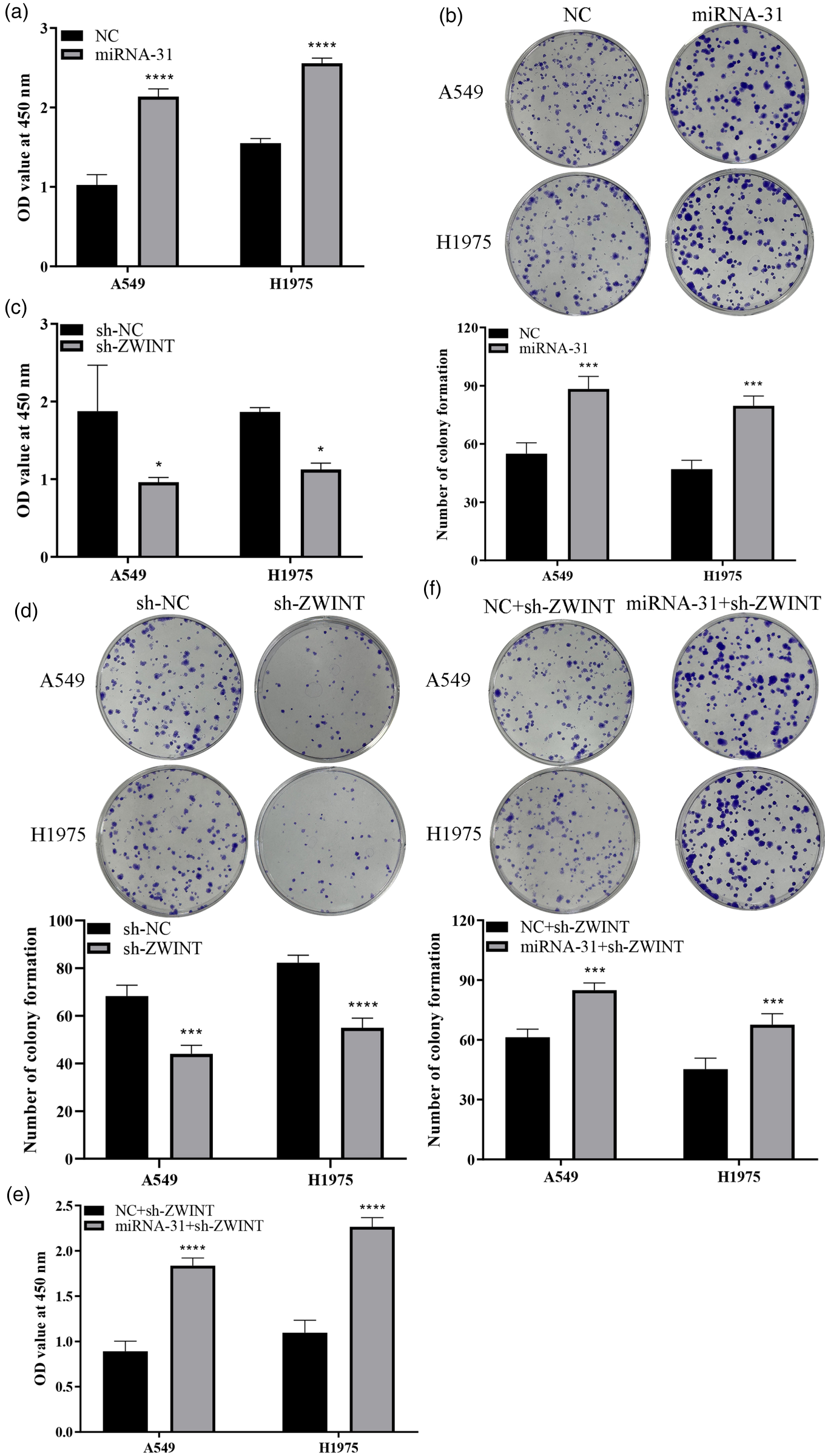
Influence of altered miR-31 and ZWINT expression on LUAD cell growth A549 and H1975. a: Transfection with miR-31 mimetic considerably enhances cancer cell vitality; b: Transfection with miR-31 mimetic elevated the number of colonies; c: Knockdown of ZWINT inhibits cancer cell vitality; d: Knockdown of ZWINT leads to a substantial elevation in the number of colonies; e: Overexpression of miR-31 reverses the reduction in cell viability caused by ZWINT knockdown; f: Overexpression of miRNA-31 counteracts the decline in cell colony count caused by ZWINT knockdown.
MiR-31 promotes LUAD cell proliferation by activating ZWINT expression, but the underlying molecular mechanism remains to be elucidated. Firstly, we divided 541 individuals into high- and low-expression categories according to ZWINT levels and conducted GSEA. We observed a strong association between high ZWINT expression and the cell cycle pathway (Figure 5(a)). We then transfected A549 and H1975 cells with sh-NC and sh-ZWINT vectors and detected cell populations at different phases utilizing flow cytometry. Comparing to the sh-NC group, the sh-ZWINT group exhibited a substantial rise in the number of G2/M phase cells, while no notable variations were detected in the number of G1 and S phase cells (Figure 5(b)). These findings indicate that downregulation of ZWINT expression induces G2/M phase cell arrest. Cyclin B1 and CDK1, essential proteins linked to G2/M phase regulation, were detected by WB. Consistent with the flow cytometry results, the levels of Cyclin B1 and CDK1 considerably reduced with the decrease in ZWINT expression (Figure 5(c)). This further indicates that reduced ZWINT levels lead to cell cycle arrest.

MiR-31 modulates the cell cycle pathway by regulating ZWINT. (a) GSEA pathway analysis utilizing ZWINT median expression; (b): ZWINT knockdown considerably impacted cell numbers in different periods; (c) ZWINT knockdown resulted in decreased expression of Cyclin B1 and CDK1; (d) Overexpression of miR-31 rescued G2/M phase cell arrest induced by reduced ZWINT expression; (e): Overexpression of miR-31 rescued the reduced expression of Cyclin B1 and CDK1 proteins caused by reduced ZWINT expression.
Given that cell cycle progression underlies cell proliferation, we speculated that the mechanism by which miR-31 regulates cell proliferation through ZWINT may involve the cell cycle pathway. To test this hypothesis, we transfected cells with sh-ZWINT followed by transfection with either miR-31-mimic and NC. Flow cytometry analysis revealed a substantial reduction in G2/M phase cells in the miR-31-mimic set relative to the NC set (Figure 5(d)). This indicates that overexpression of miR-31 mitigates cell cycle arrest induced by ZWINT downregulation. Consistently, WB results displayed that miR-31 overexpression considerably elevated the levels of ZWINT, CyclinB1, and CDK1 proteins (Figure 5(e)). Overall, the results displayed that miR-31 promotes the levels of cell cycle-associated proteins, including CyclinB1 and CDK1, by activating ZWINT expression. Consequently, this facilitates cell cycle progression, ultimately influencing cancer cell proliferation.
LUAD stands out among other cancers due to its aggressive nature and predominantly advanced stage at diagnosis, resulting in a considerably lower 5-year survival rate of less than 20% for individuals. 23 Consequently, there is an urgent need for a more comprehensive understanding of LUAD's development mechanism and the identification of effective therapeutic targets. Researchers have used an artificial neural network (ANN) model to distinguish three different subtypes of lung cancer with an accuracy greater than 93%. In addition to providing biomarkers and statistical models to diagnose lung cancer and the differentiation of lung cancer subtypes, changes in the glycophospholipid network of different subtypes of lung cancer were also found.24,25 At present, artificial intelligence (AI) has reached a high degree of accuracy in diagnosing various diseases. 26 Artificial intelligence-assisted diagnosis system could predict the invasive subtypes of early-stage lung adenocarcinoma appearing as pulmonary nodules, and has a certain predictive value.27,28 Based on miRNA data examination, our study indicated that high expression of miR-31 and miR-7974 was considerably linked to poor prognosis in LUAD. Additionally, low expression of miR-99a was linked to unfovourable prognosis, as determined through differential expression, univariate cox analysis, and survival analysis. Furthermore, we identified 246 differentially high-expressed genes and 77 differentially low-expressed genes, all of which were related to poor prognosis, based on mRNA data analysis employing the aforementioned methods. Wu et al. conducted a biogenic analysis of the lncRNA-miRNA-mRNA ceRNA network and demonstrated a substantial correlation between miR-31 and OS in LUAD individuals. 29 Other studies have also reported the detection of high levels of miR-31 in patients with metastatic LUAD. Moreover, it was demonstrated that miR-31 can enhance tumor advancement both in vitro and in vivo by activating the (Mitogen-Activated Protein Kinase)EMK/(Extracellular signal-regulated kinase)ERK signal transduction. 30 These observations suggest that miR-31 may be a novel tumor marker for LUAD and is critical in cancer onset and advancement. We proceeded to analyze the target gene of miR-31 and identified several positively correlated genes, including GOLM1, AGMAT, MCM4,GPR37, AURKA and ZWINT, following the intersection with the 246 mRNA target. Previous studies have confirmed ZWINT as a potential biomarker for poor prognosis in LUAD patients. 31 Additionally, considerably elevated expression of ZWINT has been observed in adrenal cortical carcinoma. 32 In our study, ZWINT exhibited the strongest correlation with miR-31 among the identified target genes. We confirmed through qPCR and WB experiments that increasing the expression of miR-31 resulted in a substantial rise in ZWINT expression, and vice versa. The results indicate that miR-31 positively regulates the transcription of ZWINT. We detected the expression of ZWINT mRNA and protein after overexpression or knockout of miRNA-31, and the results showed that miR-31 was likely to directly inhibit the expression of ZWINT by complementary pairing with its 3’UTR. It is worth noting that whether miR-31 indirectly regulates ZWINT through known pathways (such as ERK or Wnt/β-catenin) requires further experiments to verify binding sites or intermediate regulators. MiRNA are known to influence mechanisms, including cell growth, differentiation and apoptosis via modulating mRNA translation, inhibition or degradation. In general, miRNA degrade or inhibit the expression of target genes by binding to their 3’UTR, exerting a tumor suppressor effect. For instance, miR-576-3P targets SGK1 to suppress the growth and invasion of LUAD cells. 33 However, these are cases where miRNA can promote gene expression and contribute to cancer progression. For example, miRNA-222 promotes the growth of LUAD cells by targeting ETS1. 34 Given the positive regulation of ZWINT expression by miR-31 and the substantial upregulation of both genes in LUAD, we hypothesized that miR-31 may promote the development of LUAD by activating ZWINT expression. To validate this hypothesis, we conducted cell experiments. These observations displayed that ZWINT suppression considerably inhibited the activity and colony growth of LUAD cells, but these effects were reversed upon transfection with miR-31-mimic.
To further explored the mechanism by which miR-31 and ZWINT influence LUAD, we conducted additional investigations. LUAD individuals were categorized into high- and low-expression categories per to the median ZWINT expression. Pathway analysis by GAEA algorithm revealed a strong correlation between ZWINT and the cell cycle. Previous investigations in hepatocellular carcinoma demonstrated that ZWINT knockdown reduced cell proliferation, clonal formation, and cell cycle in HCC cells. 17 Similarly, in breast cancer, ZWINT levels revealed a positive correlation with CDK1 expression. 35 CDK1 regulates cell cycle progression by forming a complex with CyclinB1, termed CyclinB1-CDK1. 36 In line with these findings, the research demonstrated that decreasing ZWINT levels caused a substantial reduction in the protein production of CyclinB1 and CDK1, accompanied by an elevation in the proportion of G2/M phase cells. These results indicated that downregulation of ZWINT expression induced G2/M phase arrest. Upon transfection with miR-31-mimic, the proportion of G2/M phase cells was decreased, and the levels of CyclinB1 and CDK1 were considerably increased. These results indicate miR-31 has the ability to reverse the cell cycle arrest induced by ZWINT knockdown and promote the proliferation of LUAD cells.
Conclusion
Our findings unveiled that both miR-31 and its target gene ZWINT were upregulated in LUAD patients and exhibited a strong connection with poor prognosis. Notably, miR-31 exerts a positive regulatory effect on ZWINT expression. Overexpression of miR-31 alleviated the G2/M phase arrest and downregulation of cell cycle regulators CyclinB1 and CDK1 induced by ZWINT silencing in LUAD cells. These findings demonstrate that miR-31 contributes to the cell cycle progression by activating ZWINT expression, consequently driving the development of LUAD.Targeting the miR-31/Zwint axis has potential value in the treatment of lung adenocarcinoma and may drive drug development in the following ways. Novel miRNA therapies: Development of inhibitors or gene-editing tools based on the cancer-promoting effects of miR-31. Zwint targeted drugs: Explore its key role in cell cycle regulation and develop specific inhibitors.
Supplemental Material
sj-txt-1-thc-10.1177_09287329251343503 - Supplemental material for NAmiRNA-31 regulates cell cycle by activating ZWINT and promotes lung adenocarcinoma tumor development
Supplemental material, sj-txt-1-thc-10.1177_09287329251343503 for NAmiRNA-31 regulates cell cycle by activating ZWINT and promotes lung adenocarcinoma tumor development by Yan Liu, Feng Yu and Jiuyang Jiang in Technology and Health Care
Supplemental Material
sj-txt-2-thc-10.1177_09287329251343503 - Supplemental material for NAmiRNA-31 regulates cell cycle by activating ZWINT and promotes lung adenocarcinoma tumor development
Supplemental material, sj-txt-2-thc-10.1177_09287329251343503 for NAmiRNA-31 regulates cell cycle by activating ZWINT and promotes lung adenocarcinoma tumor development by Yan Liu, Feng Yu and Jiuyang Jiang in Technology and Health Care
Supplemental Material
sj-xlsx-3-thc-10.1177_09287329251343503 - Supplemental material for NAmiRNA-31 regulates cell cycle by activating ZWINT and promotes lung adenocarcinoma tumor development
Supplemental material, sj-xlsx-3-thc-10.1177_09287329251343503 for NAmiRNA-31 regulates cell cycle by activating ZWINT and promotes lung adenocarcinoma tumor development by Yan Liu, Feng Yu and Jiuyang Jiang in Technology and Health Care
Supplemental Material
sj-docx-4-thc-10.1177_09287329251343503 - Supplemental material for NAmiRNA-31 regulates cell cycle by activating ZWINT and promotes lung adenocarcinoma tumor development
Supplemental material, sj-docx-4-thc-10.1177_09287329251343503 for NAmiRNA-31 regulates cell cycle by activating ZWINT and promotes lung adenocarcinoma tumor development by Yan Liu, Feng Yu and Jiuyang Jiang in Technology and Health Care
Footnotes
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.