
Abstract
Background
Osteosarcoma (OS) has long presented a formidable challenge to human health and well-being. While traditional treatments, such as clinical chemotherapy and surgical intervention, have shown efficacy, they are frequently accompanied by adverse effects and often lead to a poor prognosis.
Objective
Thymoquinone (TQ) is recognized for its antitumor properties; however, the specific molecular mechanisms underlying its effects against OS remain inadequately understood. Emerging evidence suggests a strong correlation between p53 gene deletion and the onset and progression of various human cancers. This study aimed to elucidate the pharmacological targets and anti-OS mechanisms of TQ using systems bioinformatics approaches, including network pharmacology and molecular docking simulations.
Methods
A comprehensive screening process identified 23 potential targets associated with the anti-OS effects of TQ. Subsequent bioinformatics analysis identified 8 core targets involved in TQ's anti-OS activity. Enrichment analysis indicated that these core targets modulate a range of biological processes and may influence multiple molecular pathways.
Results
Preliminary in vitro data indicated that TQ effectively reduces OS cell proliferation, induces apoptosis, and downregulates the expression of P53 and HMOX1 proteins.
Conclusion
Our findings elucidate the molecular mechanisms underlying TQ's effectiveness against OS, highlighting potential apoptosis-related therapeutic targets, such as P53 and CYCLIN D1, for the treatment of OS with TQ.
Introduction
Osteosarcoma (OS) represents the most aggressive form of malignant bone disease, predominantly affecting children and adolescents. Its global annual incidence ranges from 1 to 3 cases per million. 1 The standard treatment regimen typically involves a combination of neoadjuvant chemotherapy and surgical resection, which has led to improved survival rates; however, challenges remain due to high recurrence rates and mortality associated with metastasis. 2 Thymoquinone (TQ), a naturally occurring compound with a historical usage of nearly 2000 years as a volatile oil, has demonstrated antitumor activity with minimal adverse effects in various in vitro and in vivo studies, including those focused on colon, lung, leukemia, and pancreatic cancers.3–5 Notably, TQ has been shown to inhibit cancer cell growth and survival while preserving healthy cells. 6 Although current research highlights TQ's inhibitory effects on OS, a more thorough investigation into its underlying mechanisms of action against this malignancy is necessary. 7 The field of preclinical bioinformatics has advanced significantly with the introduction of cyberpharmacology applications, which can comprehensively identify the targets and mechanisms of natural compounds in cancer treatment. 8 The integration of network pharmacology with molecular docking simulations has been pivotal in the investigation and development of potential anticancer therapies. 9 Therefore, this study aimed to elucidate the biological targets and molecular mechanisms through which TQ exerts its anti-OS effects, utilizing an extensive bioinformatics framework that incorporates network pharmacology and molecular docking analyses. Such computational pharmacology platforms represent emerging health technologies that support precision oncology by integrating large scale biological data, predictive modeling and therapeutic target prioritization, thereby enhancing translational efficiency in cancer research. Furthermore, the bioinformatics results were corroborated by experimental evaluations in cell cultures. In summary, molecular docking technology and protein validation have elucidated the substantial therapeutic potential of TQ in the treatment of osteosarcoma. When considered alongside research findings on other natural compounds, TQ's multi-target and multi-pathway mechanisms of action not only offer theoretical support for its clinical application but also establish a foundation for future experimental and clinical investigations. These findings underscore the indispensable role of molecular docking technology in uncovering the anticancer mechanisms of natural compounds, thereby providing novel directions and opportunities for osteosarcoma treatment.
Materials and methods
Screening of TQ and OS targets and identification of common targets
“Thymoquinone” as the primary keyword, we screened PharmMapper, STITCH, and SwissTargetPrediction databases to obtain potential targets associated with TQ. By combining TQ and de-emphasizing TQ-related potential target genes from the aforementioned databases, we standardized protein target information using the Uniprot protein database.
Using “Osteosarcoma” as keywords, we searched the OMIM and GeneCards disease databases for targets linked to OS.
Subsequently, by inputting the identified TQ and OS target genes into Venn 2.1, we identified 23 common gene targets through the intersection of potential TQ and OS gene targets.
Network construction and core target identification-computational health technology workflow
By inputting the shared targets related to TQ and OS into the STRING database (https://cn.string-db.org/), and specifying the “Homo sapiens” species along with a confidence level exceeding 0.4, we constructed a protein-protein interaction (PPI) network for these targets. Subsequently, the generated PPI network was imported into Cytoscape software, and using the NetworkAnalyzer tool, we calculated key topological properties such as Degree, Betweenness, and Clustering coefficient. Nodes with a Degree greater than the median were identified as core targets and were organized into a PPI graph. The computational workflow applied in this study represents a health technology driven pipeline integrating multi source biomedical databases, network analysis and molecular simulation tools. This framework enables systematic target identification and prioritization for osteosarcoma therapy.
Core target gene enrichment analysis
Conducting Gene ontology (GO) and Kyoto encyclopedia of genes and genomes (KEGG) enrichment analyses on the prospective core targets for TQ treatment using Metascape (https://metascape.org/gp), we established the minimum enrichment factor at 1.5, minimum overlapping targets at 3, and set the threshold at P < 0.01. The enrichment analysis was achieved through microbiometric online mapping. Visualization of the enrichment outcomes was achieved via bubble plots utilizing the Microbiology online mapping platform (https://www.bioinformatics.com.cn/).
Molecular docking
Molecular docking was performed to investigate the interaction of TQ with potential core targets identified for OS treatment. The 2D compound structure was converted to a 3D structure using Open babel 2.3.2 software. The target 3D structure was obtained from the PDB database (http://www.rcsb.org/pdb) and pre-processed using PyMOL 2.5.1 software. The 3D structures of the active ingredient and the target were converted to PDBQT format using AutoDockTools 1.5.6 software. Subsequently, semi-flexible docking was carried out using AutoDock Vina 1.1.2 molecular docking software. The best conformations were selected and visualized using PyMOL 2.5.1 and Ligplot v.2.2.8 to analyze the docking results. All analyses were conducted using established, widely adopted computational platforms and potential adaptability of this digital workflow to other malignancies”.
Cell culture
The anti-proliferative effects of TQ against OS were assessed using the MG-63 cell line. MG-63 OS cells (MEM-EBS: Minimum Essential Medium [MEM Eagles with Earle's Balanced Salts] + 10% fetal bovine serum [FBS] + 1% NEAA + 1% Penicillin-Streptomycin Solution) were provided by the Institute of Basic Research, Peking Union Medical College. The cells were cultured as monolayers in their respective media and maintained at 37°C under saturated humidity with 5% carbon dioxide. TQ (Sigma) was dissolved in anhydrous alcohol to achieve a concentration of 20 mmol/L.
Cell cycle analysis
The percentage of cells in the G0-G1, S, and G2-M phases of the cell cycle was determined using the Cycle Test™ Plus DNA kit (4A BIOTECH, Suzhou, China). MG-63 cells were seeded in 6-well plates at a density of 1 × 105 cells per well. After treatment with the IC50 value of TQ for 48 h, the supernatant was discarded, and the cells were washed with phosphate-buffered saline (PBS) and then centrifuged. The cells were further washed twice with ice-cold PBS and subsequently treated with 3 ml of ice-cold 70% ethanol. Following incubation at 4°C for 1 h, the cells were washed twice with PBS, and 10 mg/ml RNase A was added. Propidium iodide (PI) was added to the tubes at a final concentration of 0.05 mg/ml, and the cells were incubated in the dark for 30 min at 4°C. Subsequently, the cells were centrifuged and analyzed using flow cytometry (Beckman, USA) as previously described to determine the percentage of cells at each stage of the cell cycle. These assays were performed in triplicate and according to the manufacturer's instructions.
Western blot assay
The protein expression of P53, HMOX1, CYCLIN D1, and β-actin (Cytosci, Beijing, China) was assessed using Western blotting. The procedure followed the established protocol described in the literature. Briefly, cell lines were washed twice with PBS, homogenized by sonication in lysis buffer, and centrifuged to remove impurities. Total protein lysate samples (20 µg) were resolved on 12% polyacrylamide SDS gels and transferred onto polyvinylidene difluoride (PVDF) membranes using electrophoresis. The membranes were blocked with 5% skim milk in TBST buffer (TBS containing 0.1% Tween-20), incubated with the respective primary antibodies, and subsequently probed with alkaline phosphatase-conjugated secondary antibodies. Protein bands were visualized using 5-bromo-4-chloro-3-indolyl phosphate/nitroblue tetrazolium. Afterward, the membranes were washed, and the protein bands were detected by exposing the membrane to X-ray film. β-actin was used as a loading control.
Statistical analysis
The data were presented as mean ± standard deviation (SD) from a minimum of three independent experiments. Statistical significance was determined using the Student's t-test, and a P-value of less than 0.05 was considered statistically significant.
Results
Identification of TQ and OS Targets and construction of core target networks
A total of 40 targets for TQ and 5324 genes associated with OS were identified. Using the Venn2.1 tool, 23 potential gene targets for TQ treatment of OS were identified from the overlapping genes. These targets were further analyzed using the STRING database to construct a network. The resulting network contained 23 nodes and 61 edges, with an average node degree of 5.3. To filter for core targets, the filtering condition was set to a degree greater than 10, resulting in the identification of 8 key targets. These targets were then reconstructed into a PPI graph, as depicted in Figure 1. Figure 1 shows the PPI network diagram.
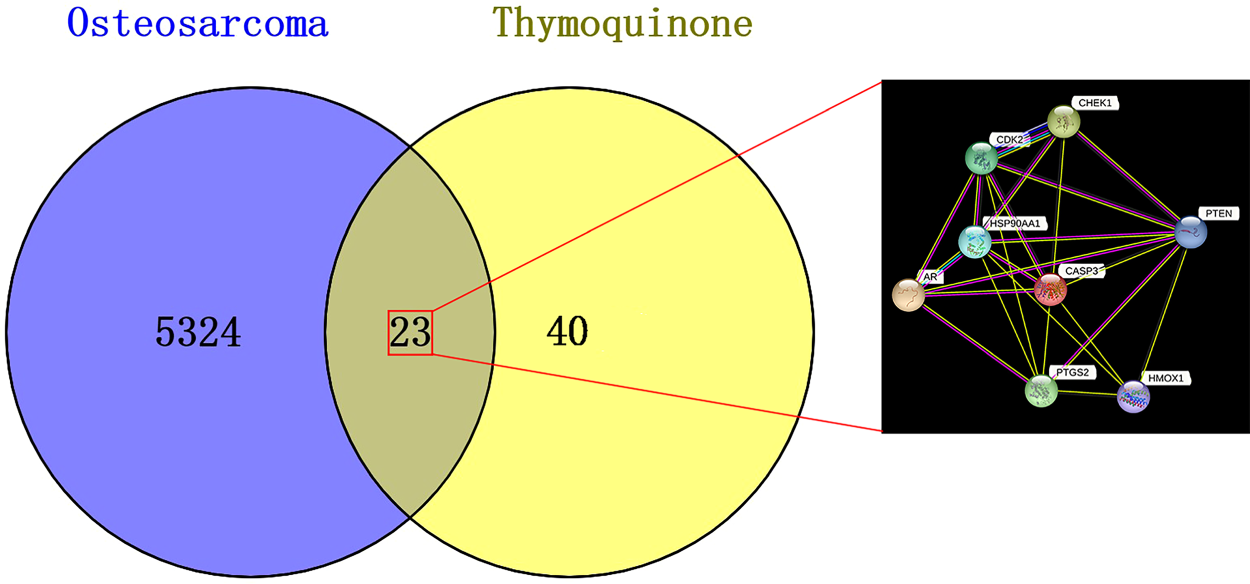
Venn diagram depicting the shared targets between TQ and OS, highlighting the common targets within the gene linkage network.
Pathway enrichment analysis and network visualization are pivotal methodologies in the realm of computational biology, primarily supporting data-driven target prioritization. These approaches collectively empower researchers to derive meaningful insights from complex biological data, especially in the context of drug discovery and personalized medicine. To explore the functional implications of the potential core targets of TQ in OS treatment, GO enrichment analysis was conducted. The top three results in the biological process (BP) category included extrinsic apoptotic signaling pathway, peptidyl-serine phosphorylation, and cellular response to abiotic stimulus (Figure 2). In the cellular component (CC) category, the top five results were myelin sheath, membrane raft, and membrane microdomain (Figure 3). For molecular function (MF), the top five enriched terms were histone kinase activity, protein tyrosine kinase binding, and proteinase binding (Figure 4).
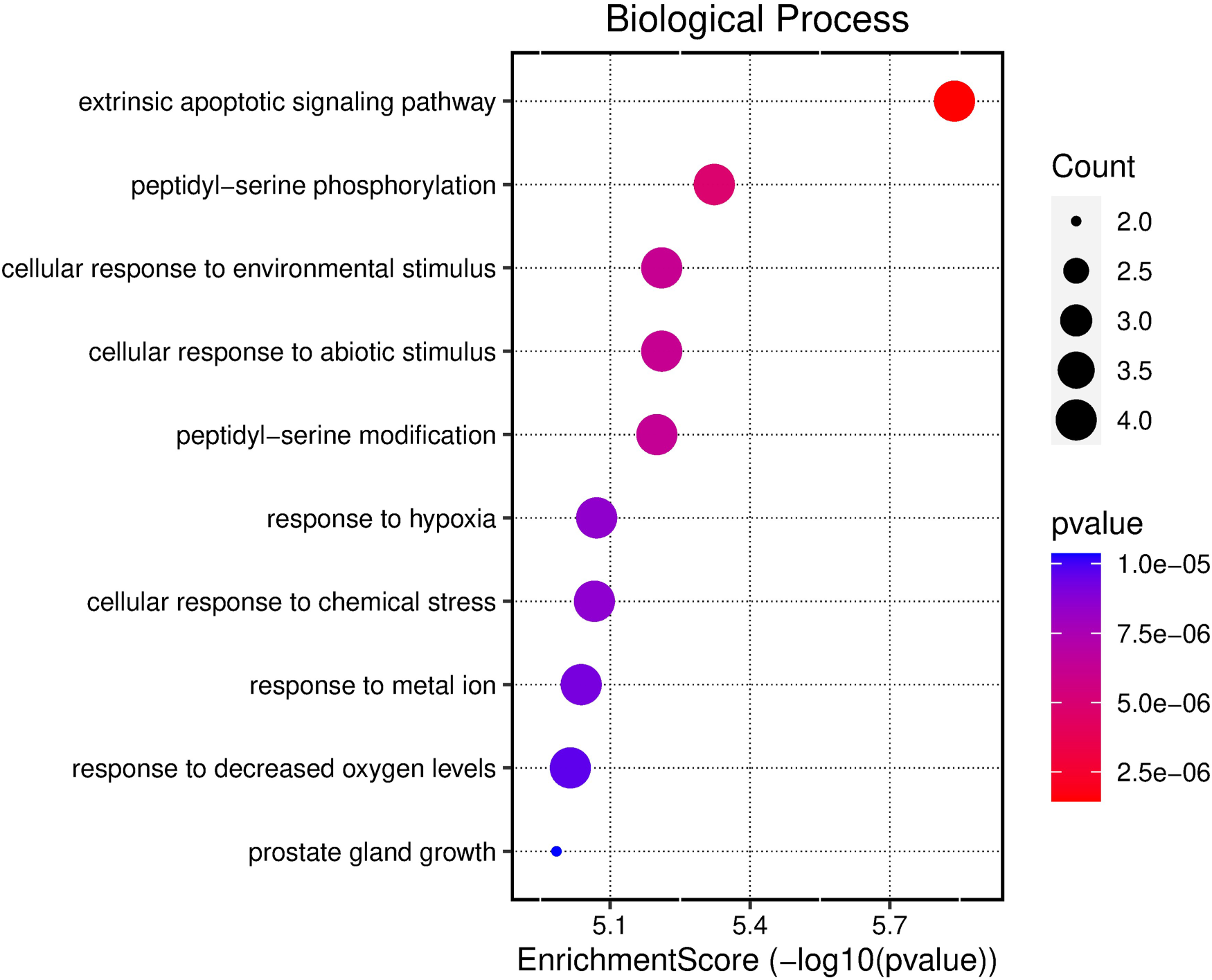
Bubble chart illustrating the results of GO functional enrichment analysis using all core targets to determine the BPs of TQ's anti-OS activity.
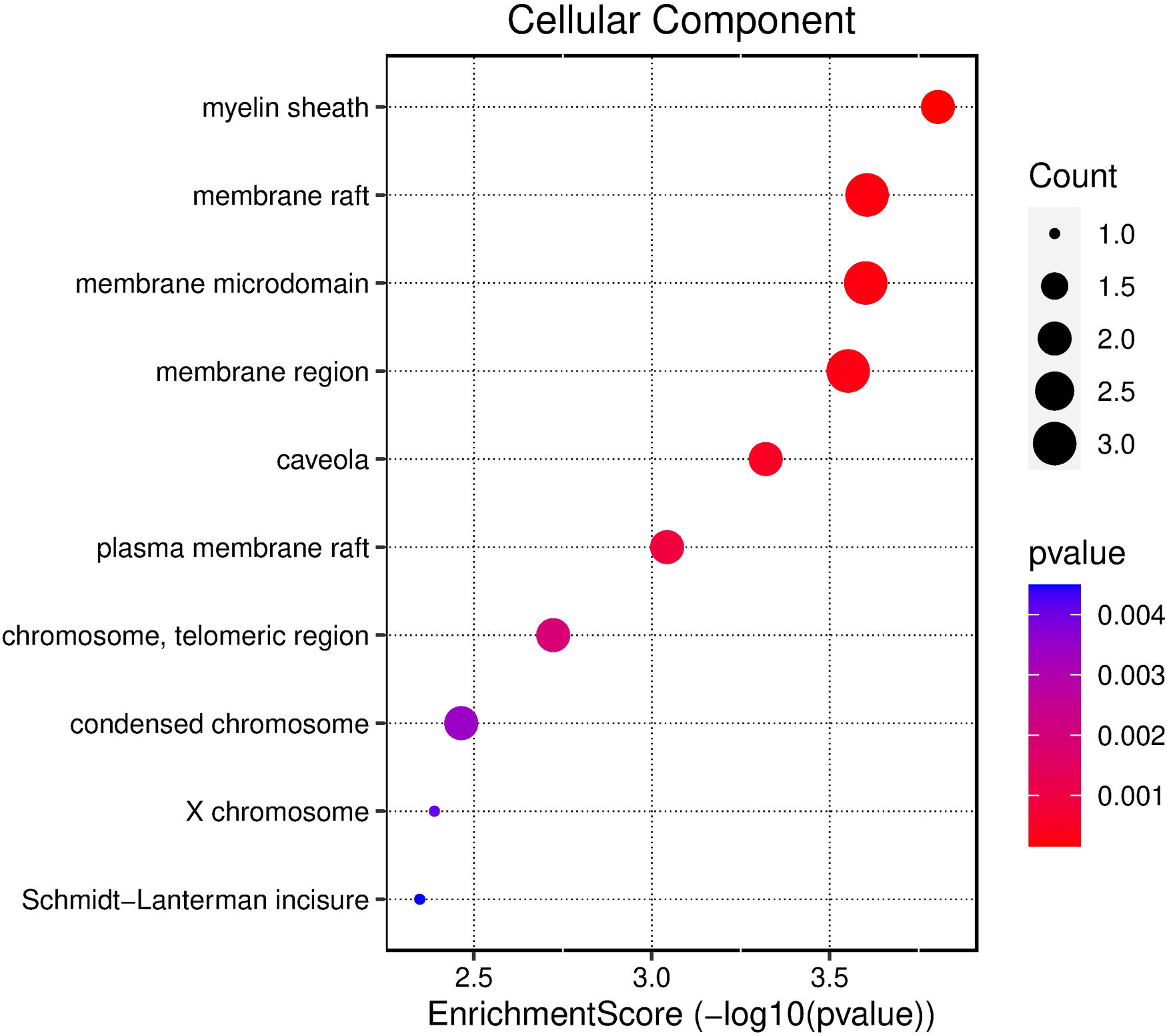
Bubble chart depicting the results of GO functional enrichment analysis using all core targets to determine the cellular localization of TQ's anti-OS activity.
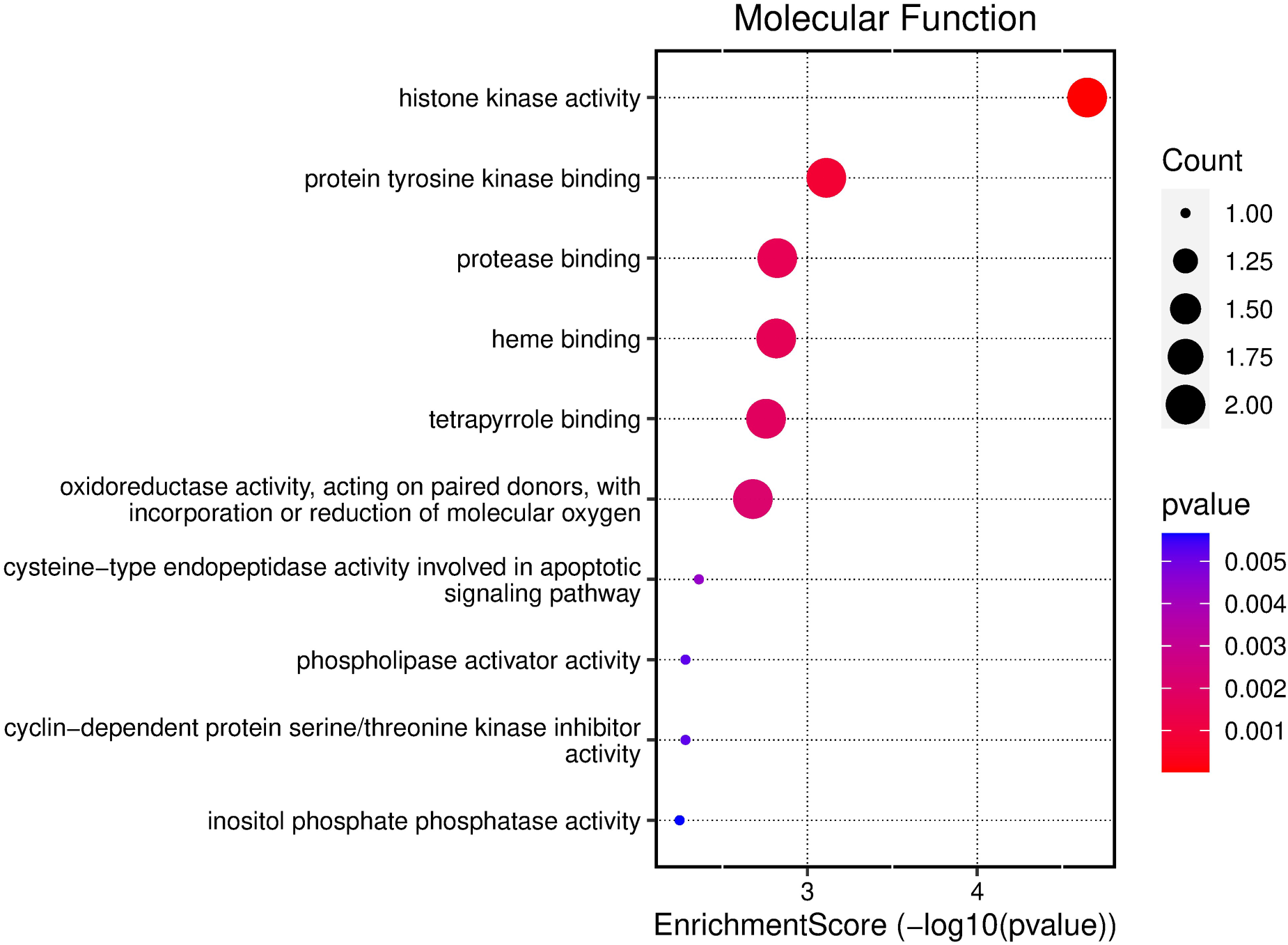
Bubble chart illustrating the results of GO functional enrichment analysis using all core targets to determine the MFs of TQ's anti-OS activity.
In addition, KEGG pathway analysis revealed the top three enriched pathways as the p53 signaling pathway, small cell lung cancer, and prostate cancer (Figure 5). Cytoscape software was used to integrate the 10 KEGG signaling pathways enriched in the core targets of TQ in OS treatment, resulting in the construction of a component-target-pathway network diagram (Figure 6). From a health technology standpoint, enrichment and network visualization enabled rapid identification of clinically relevant signaling pathways, demonstrating the feasibility of computational tools in supporting oncological mechanism discovery.
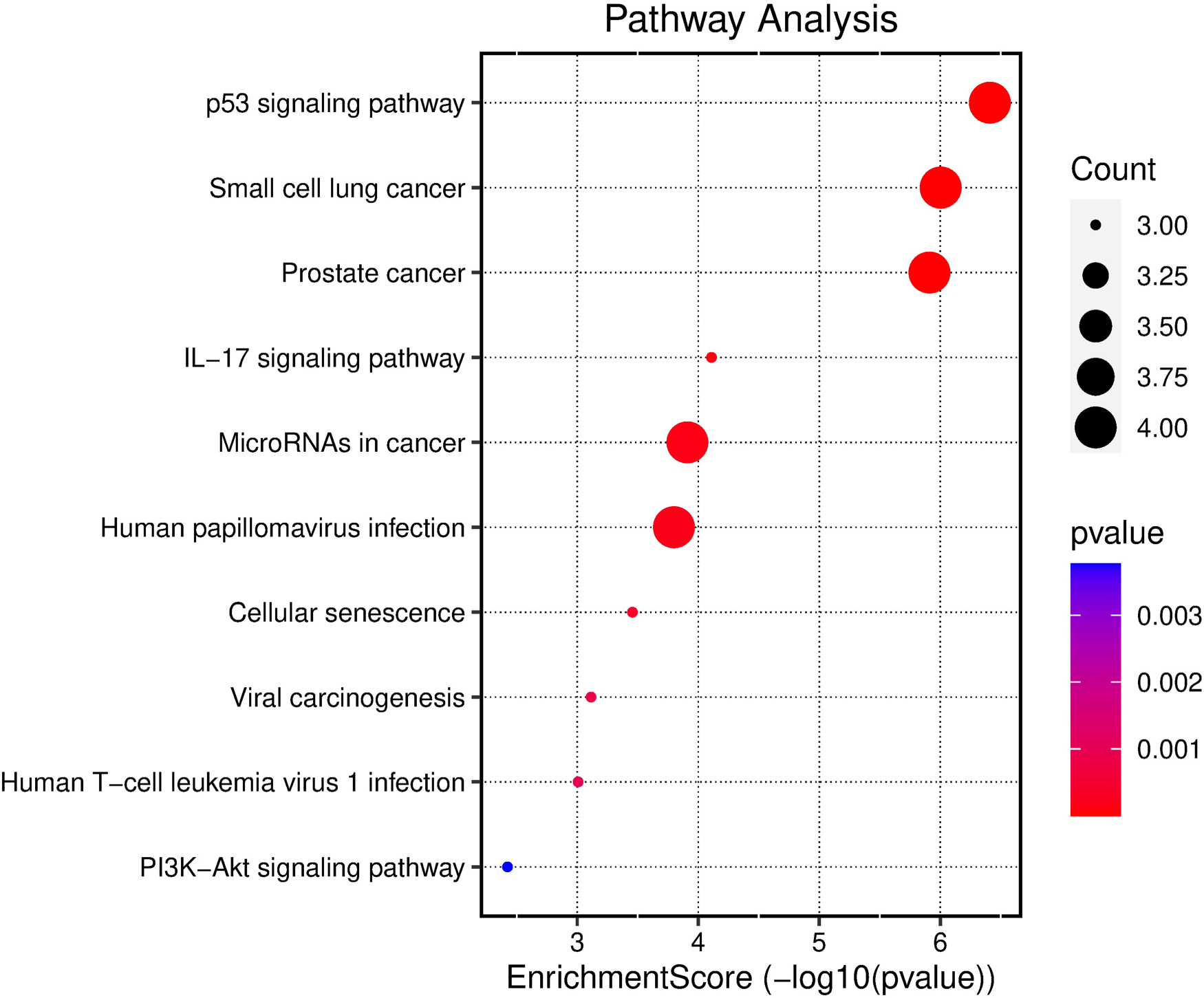
Bubble chart displaying the results of KEGG pathway analysis for all core targets of TQ in the treatment of OS.
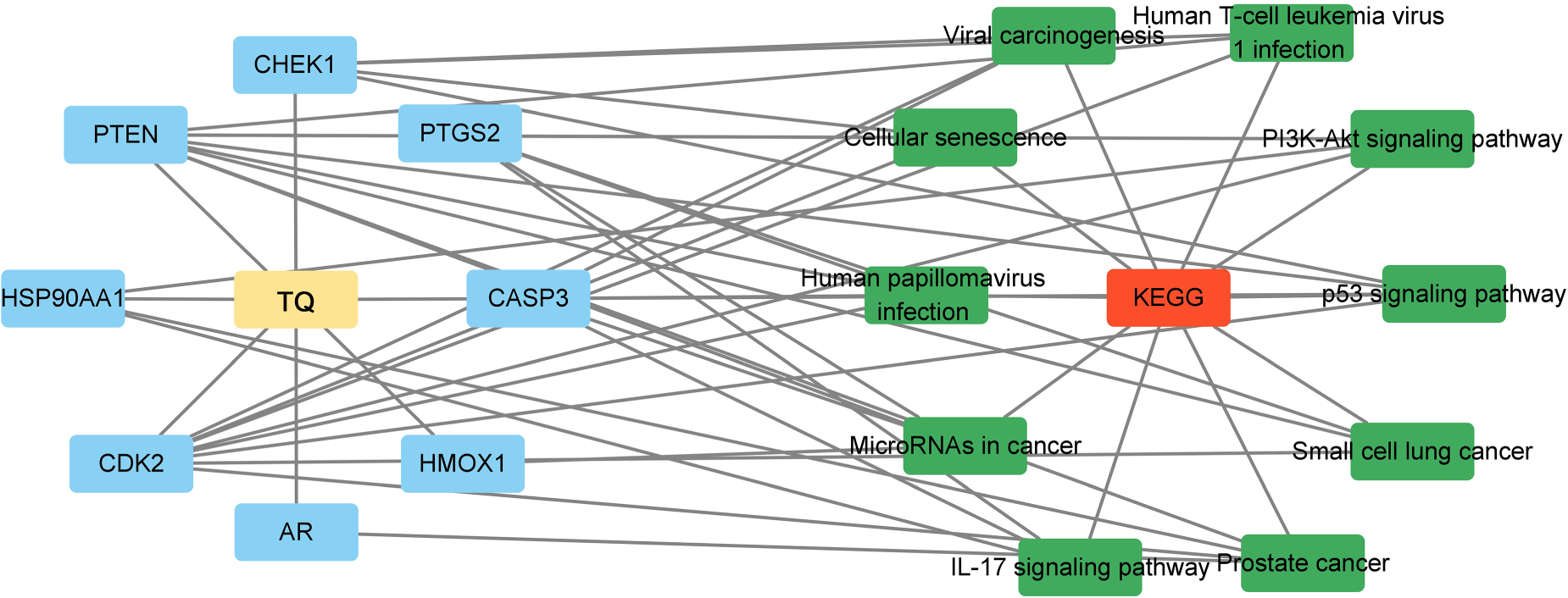
Cytoscape software was used to integrate the 10 enriched KEGG signaling pathways in the core targets of TQ for the treatment of OS. A component-target-pathway network diagram of TQ was constructed for the treatment of OS.
The stability of molecular docking can be evaluated based on the magnitude of the binding energy. 10 A smaller binding energy indicated a lower energy requirement for binding and signified a more stable interaction. Based on the docking score data, the absolute value of the binding energy between TQ and HMOX1 was 7.40 kcal/mol, indicating a strong binding affinity between the active ingredient and the core target. Figure 7 is the molecular docking diagram showing the interaction between TQ and the HMOX1 protein. Hydrophobic interactions were observed between TQ and residues Ser53, Asn210, Arg136, Phe33, Phe166, Phe167, Leu54, Phe37, Met34, Val50, and Phe214 of the HMOX1 protein.
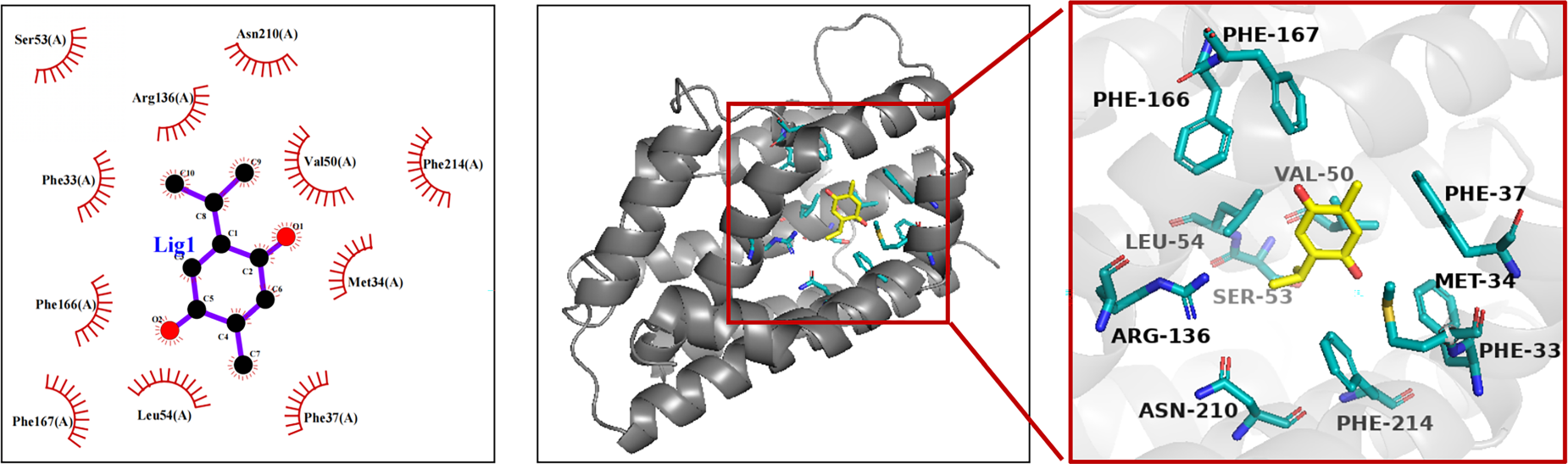
Molecular docking analysis illustrating the interaction of TQ with the core targets.
To determine the effect of TQ on cell growth, the MG-63 cell line was exposed to various concentrations of TQ (ranging from 0 to 20 µM) for 24 and 48 h. Cell viability was assessed by MTT assay. As shown in Figure 8, TQ treatment resulted in a dose- and time-dependent inhibition of cell growth in the MG-63 cell line. The IC50 values, representing the half-maximal inhibitory concentrations of TQ, were determined to be approximately 15.66 µM at 24 h and 10.20 µM at 48 h. Notably, treatment with TQ at 10 and 15 µM for 48 h had a potent antiproliferative effect on the MG-63 cell line, independent of time of action at 20 µM.
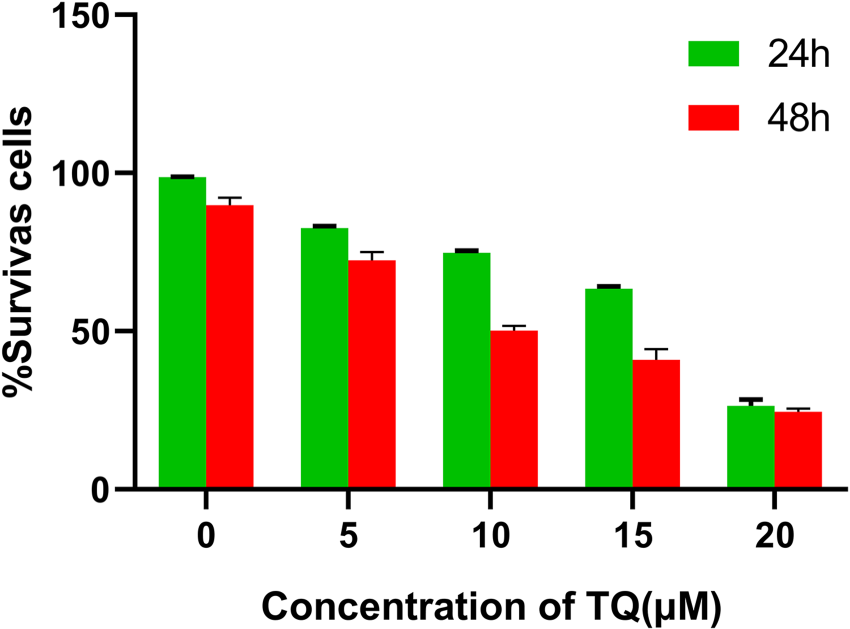
Inhibition of MG-63 cell activity by TQ in vitro.
Building upon the preliminary evaluation of TQ's impact on MG-63 cell growth, the researchers analyzed cellular DNA content using flow cytometry to examine the effect of TQ on cell cycle distribution. As shown in Figure 9, treatment with TQ for 48 h resulted in a significant dose-dependent arrest of MG-63 cells in the G0/G1 phase of the cell cycle. The percentages of cells in the G0/G1 phase for TQ treatments of 0, 5, 10, 15, and 20 µM were determined to be 38.81 ± 2.09%, 38.37 ± 1.26%, 33.73 ± 5.15%, 28.30 ± 3.05%, and 22.42 ± 3.76%, respectively (Figure 9). The increase in the percentage of cells in the G2/M phase was accompanied by a corresponding decrease in the percentage of cells in the G0/G1 phase. The observed increase in the proportion of G2/M phase cells was consistently accompanied by a decrease in the proportion of cells in the G0/G1 phase.
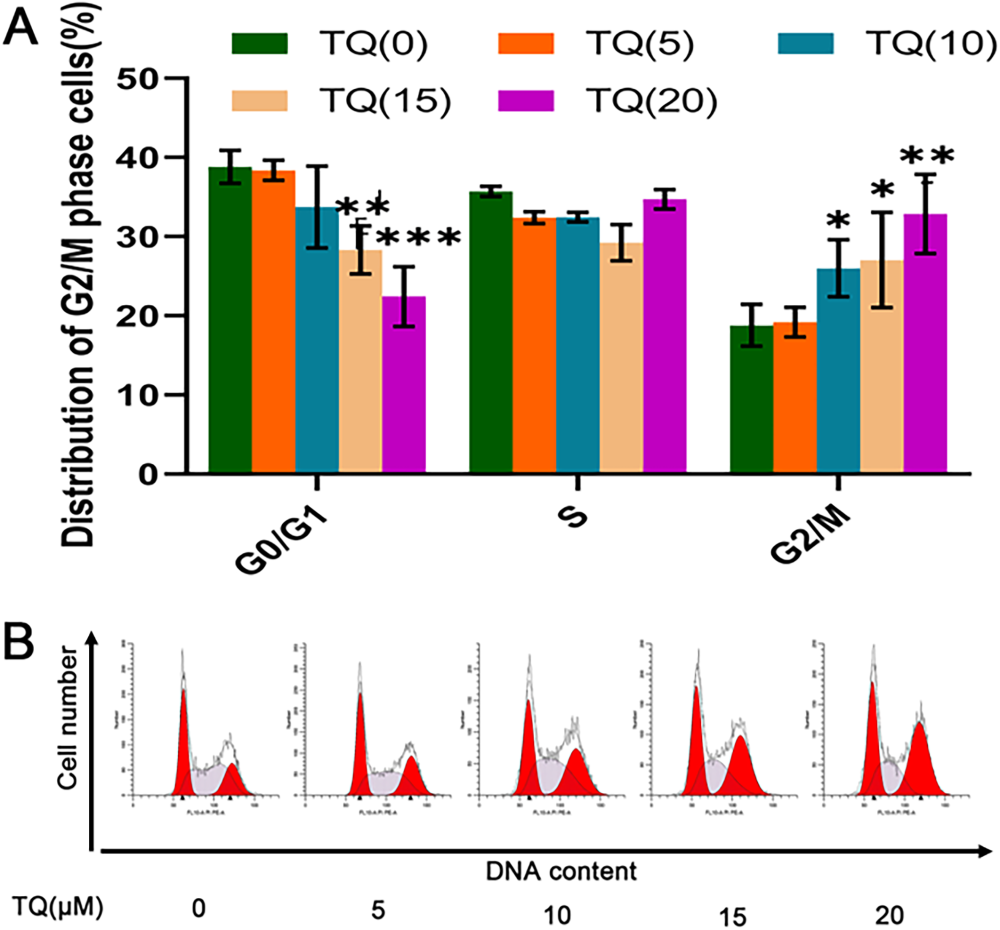
In vitro cell cycle progression. (A) Representative flow cytometry histograms of cell cycle distribution of MG-63 cells treated with lysate control and TQ (5, 10, 15, and 20 µM) for 48 h. (B) The cell cycle distribution data represents the results from at least three independent experiments. Compared to the control, *p < 0.05; **p < 0.01; ***p < 0.001.
Cell line experiments were performed to validate the results obtained from molecular docking bioinformatics analysis. Comparing the untreated cells in the control group, the protein expression of p53 showed the highest inhibition at 20 µM, with statistically significant (P < 0.001). Additionally, there was a significant difference between the 10 µM and 20 µM groups (P < 0.05). Similarly, HMOX1 and Cyclin D1 proteins also showed the highest inhibition at 20 µM with statistical significance (P < 0.001). However, there were no significant differences observed between the 5 µM and 10 µM groups for HMOX1 protein (P > 0.05), nor between the 10 µM and 15 µM groups for Cyclin D1 protein (P > 0.05). Moreover, there was no significant difference between HMOX1 and Cyclin D1 proteins at 20 µM (P < 0.001), the 5 µM and 10 µM HMOX1 proteins (P > 0.05), or the 10 µM and 15 µM Cyclin D1 proteins (P > 0.05). These results, as shown in Figure 10, indicate that TQ effectively inhibited the upregulation of proteins in MG-63 cells at concentrations of 15 µM or higher.
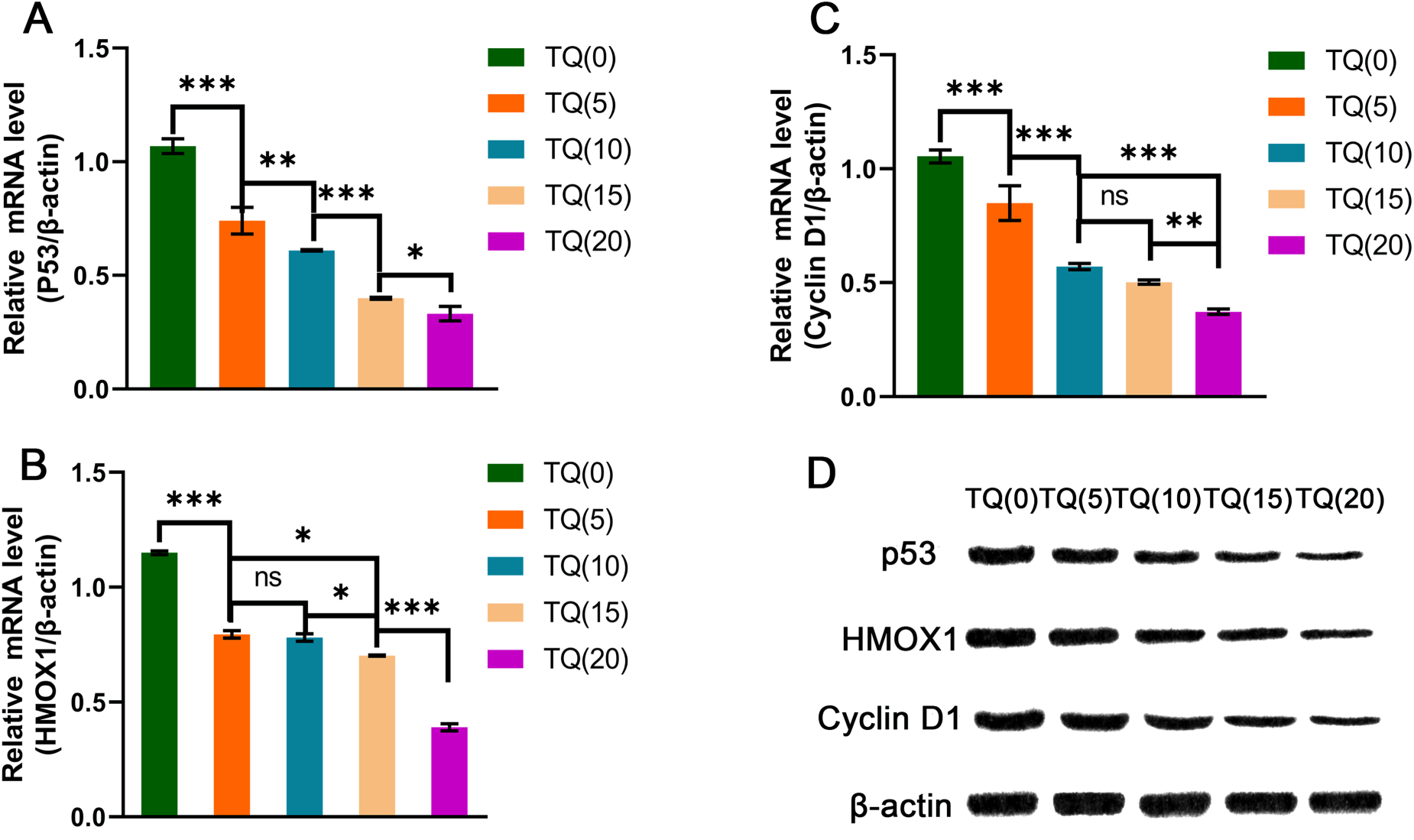
mRNA levels in MG-63 cells and Western blot analysis. nsp > 0.05; *p < 0.01; **p < 0.01, ***p < 0.001.
OS represents a prevalent primary malignant neoplasm of the bone, primarily impacting adolescents and young adults. Although advancements in therapeutic interventions have been made in recent years, the management of osteosarcoma continues to pose significant challenges, especially in cases of metastasis and recurrence. Traditional chemotherapy protocols have demonstrated limited effectiveness in the treatment of osteosarcoma, thereby underscoring the need for the development and investigation of innovative therapeutic approaches to enhance patient survival outcomes and quality of life. OS is a complex disease with multifactorial origins, characterized by genomic instability, chromosomal abnormalities, and associations with specific syndromes. 11 A key factor contributing to the proliferation of OS cells is their resistance to apoptosis, which allows them to exhibit remarkable resilience and continue to grow despite disruptions in cell-cell and cell-matrix attachments. 12 The transcription factor p53 plays a critical role in regulating gene-associated DNA repair and initiating apoptotic cell death following cellular injury. 13 OS occurs in approximately 30% of patients with Li-Fraumeni syndrome, and p53 deficiency is observed in 18–26.5% of sporadic OS cases. 14 Therefore, targeted anti-value-added therapy may represent a promising therapeutic strategy for OS. To address the clinical challenges posed by value-added-induced drug resistance and poor long-term prognosis, TQ, a compound with significant antitumor effects, has been shown to impact various signaling pathways and cellular processes, effectively targeting numerous cancer types. 7 In time-dependent experimental studies, TQ has been demonstrated to upregulate p53 expression and induce apoptosis in MCF7 cells, there by effectively inhibiting cancer cell proliferation. 15 Additionally, recent investigations into the molecular mechanisms of TQ in A549 lung cancer cells have shown that TQ incubation results in a reduction of proliferating cell nuclear antigen (PCNA) expression, a proliferation marker, as well as a decrease in Cyclin D1 levels. 16
In our study, we initially examined the effects of TQ on the MG-63 cell line by administering concentrations ranging from 0 to 20 µM over 24 and 48 h. Our findings indicated that the efficacy of TQ was contingent upon the duration of treatment, suggesting a time-dependent effect.17,18 The p53 gene is integral to the regulation of apoptosis. Activation of the p53 transcription factor is induced by external stressors and can be initiated by oncogene activation. 13 Once activated, p53 orchestrates a range of cellular processes, including apoptosis, cell cycle progression, autophagy, and DNA repair.12,19,20 In our study, we found that treatment with TQ at a concentration of 15 µM resulted in a reduction of p53 protein levels, suggesting an absence of excessive oncogene activation that would otherwise elevate p53 expression. Additionally, our cell cycle analysis demonstrated a pronounced G0/G1 arrest, marked by the cessation of protein synthesis and cell division. The expression of Cyclin D1, a key regulator of the G1 phase, was consistencyt with these cell cycle findings. The p53 signaling pathway is implicated in early oncogenesis, and its disruption can lead to significant alterations in cell cycle dynamics.2,4
In this study, a network pharmacological analysis was conducted to examine the relationship between TQ and target genes associated with OS. The findings indicated that four out of eight core genes, HMOX1, CDK2, AR, and PTGS2, demonstrated binding energies with absolute values exceeding 6 kJ/mol, signifying a strong interaction with TQ. 21 Molecular docking experiments further revealed that TQ exhibited the highest affinity for HMOX1. Subsequent analysis utilizing Gene Ontology (GO) functional data identified enriched terms across BP, CC, and MF categories. Within the BP annotations, terms related to the extrinsic apoptotic signaling pathway, peptidyl-serine phosphorylation, and cellular response to abiotic stimulus were associated with the formation and progression of OS, as well as the anti-OS effects of TQ. Furthermore, enrichment analysis identified other significant pathways through the KEGG. The study elucidated TQ's targeted anti-OS mechanisms by highlighting signaling pathways enriched with target genes, including the p53 signaling pathway, small cell lung cancer, and prostate cancer. The p53 signaling pathway is recognized for its pivotal role in OS cell proliferation through the regulation of key MFs.10,22 Furthermore, a positive association between the p53 signaling pathway and OS cell proliferation underscores its critical importance in OS pathogenesis. The influence of Thymoquinone (TQ) on this pathway indicates its potential as an anti-OS agent by modulating essential MFs.
Researchers have continuously explored novel therapeutic strategies to improve the prognosis of osteosarcoma patients. Among these, thymoquinone (TQ), as a promising therapeutic agent for osteosarcoma, has gradually garnered attention in the academic community. TQ emerges as a promising candidate for anti-OS therapy, interacting with core target genes within the p53 signaling pathway. Herbal isolates, such as TQ, have been traditionally utilized in the management of various human cancers.6,7 Nevertheless, the precise mechanisms and primary targets of TQ in OS treatment remain inadequately elucidated. In this study, molecular docking analysis was employed to assess the applicability of network pharmacology in identifying potential targets. Experimental validation using cell line samples corroborated the molecular docking findings. Beyond biological findings, this study highlights the utility of network pharmacology and molecular docking as health technology tools in oncology research. Such digital approaches can assist clinicians and researchers in prioritizing therapeutic targets, designing combination strategies and informing personalized treatment hypotheses. These results underscore the utility of network pharmacology as an effective tool for target identification in OS treatment. Thymoquinone (TQ), as an emerging therapeutic agent for osteosarcoma, demonstrates substantial antitumor efficacy when used as a monotherapy and shows potential synergistic effects when employed in combination therapies. Future investigations should aim to elucidate the precise mechanisms underlying TQ's action and assess its clinical applicability in osteosarcoma treatment, with the ultimate goal of offering more effective therapeutic strategies for patients afflicted with this malignancy.
Conclusion
The comprehensive findings derived from network pharmacology, molecular docking, and experimental validation suggest that TQ exhibits significant therapeutic potential in the treatment of osteosarcoma by specifically targeting genes and pathways associated with apoptosis. The compound's synergistic interactions with other anticancer agents, along with its ability to modulate multiple signaling pathways, underscore its promise as an effective anti-osteosarcoma agent. These findings offer essential theoretical and experimental underpinnings for the clinical application of TQ in osteosarcoma therapy, thereby justifying further research and development in this area. Importantly, this work demonstrates the value of computational health technology tools in cancer research, providing a reproducible and scalable framework for data driven therapeutic exploration in osteosarcoma.
Footnotes
Acknowledgments
Fund program: Heilongjiang Postdoctoral Fund of China (LBH-Z23256)
Author contributions
Zhange Yu conceived and supervised the study; A series of operations were performed by Chengliang Zhou that included experiment performed, new software developed, datas analysed and the manuscript wrote; Ruiyao Wang, Jiaqi Bi, Fang Liu and Kun-Yu Liu made the manuscript revisions. All reseachers reviewed the experimental datas and approved the manuscript eventually published.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Heilongjiang Postdoctoral Fund of China, (grant number LBH-Z23256).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.