
Abstract
Prior research indicates that being reared in stressful environments is associated with earlier onset of menarche in girls. In this research, we examined (a) whether these effects are driven by exposure to certain dimensions of stress (harshness or unpredictability) during the first 5 years of life and (b) whether the negative effects of stress on the timing of menarche are buffered by secure infant-mother attachment. Results revealed that (a) exposure to greater harshness (but not unpredictability) during the first 5 years of life predicted earlier menarche and (b) secure infant-mother attachment buffered girls from this effect of harsh environments. By connecting attachment research to its evolutionary foundations, these results illuminate how environmental stressors and relationships early in life jointly affect pubertal timing.
Decades of theory and research converge on the conclusion that exposure to stressful environments in childhood accelerates pubertal development in girls as measured by onset of menarche (e.g., Belsky et al., 2007; Belsky, Steinberg, Houts, & Halpern-Felsher, 2010; Deardorff et al., 2011; Ellis & Essex, 2007; Ellis & Garber, 2000; Moffitt, Caspi, Belsky, & Silva, 1992; Surbey, 1990; for reviews, see Ellis, 2004, and Belsky, 2012). Although age at menarche is heritable (e.g., Belsky, 2000; Rowe, 2000), studies incorporating genetic controls have confirmed that the association between early-life stress and age at menarche is robust (e.g., Belsky, Houts, & Fearon, 2010; Tither & Ellis, 2008).
Many studies, however, have operationalized early-life stress broadly at the cost of specifying the types of stress that may be more or less important in predicting menarcheal timing. Drawing on Belsky, Steinberg, and Draper’s (1991; Belsky, 1997) evolutionary theory of socialization and Ellis, Figueredo, Brumbach, and Schlomer’s (2009) life history model of environmental stress, we address two important questions about the connection between stress and reproductive maturity: Do specific types of environmental stress encountered early in life uniquely predict age of menarche? Are the effects of environmental stress on menarcheal age buffered by the security of the infant-mother attachment relationship?
Life-History Theory, Stress, and Age of Menarche
Life-history theory explains how organisms allocate their inherently limited resources to optimize reproductive fitness (Kaplan & Gangestad, 2005; Stearns, 1992). According to this evolutionary biological theory, developing organisms face a fundamental trade-off between allocating resources to somatic effort (i.e., growth and maintenance) versus reproductive effort (i.e., sexual maturation and mating). Central to life-history thinking is the premise that the specific strategy an individual adopts depends in part on his or her early rearing environment (Belsky et al., 1991; Chisholm, 1999). In environments that are harsh or unpredictable (or both), for example, faster strategies that prioritize greater reproductive effort increase the likelihood of reproducing before death. However, when harshness or unpredictability (or both) are low (or can be managed), slower strategies that prioritize investment in somatic effort are more adaptive (Belsky et al., 1991; Ellis et al., 2009). Neither fast nor slow strategies are inherently better. Instead, the evolutionary benefit of a given strategy depends on the environment in which an individual develops (Belsky, 1997; Belsky et al., 1991; Caudell & Quinlan, 2012; Kuzawa, McDade, Adair, & Lee, 2010; Simpson & Belsky, 2008, in press).
Belsky et al. (1991) were the first to specify how life-history strategies should unfold across the life span. They advanced a unique prediction about the effects of childhood stress on development: Exposure to greater childhood stress (e.g., inadequate resources, an absent father, insensitive or harsh parenting, marital conflict) should accelerate pubertal maturation. Considerable evidence consistent with this proposition has emerged for females, particularly with regard to age of menarche (Belsky, 2012). For example, greater family disruption (especially fathers’ social deviance) predicts earlier age of menarche (Tither & Ellis, 2008). Maltreated girls reach pubertal maturity 8 months earlier than girls who are not maltreated (Costello, Sung, Worthman, & Angold, 2007), and earlier harsh maternal care forecasts earlier menarche (Belsky et al., 2007; Belsky, Steinberg, et al., 2010). Taking advantage of an existing situation, Pesonen et al. (2008) observed that young Helsinki girls who were evacuated from their homeland during World War II and sent to other countries reached menarche at a younger age (and also bore more children by late adulthood) compared with young girls who remained at home and avoided the trauma of separation from their families.
In modern Western societies, in which nutritional deprivation is not as severe as in underdeveloped countries, lower socioeconomic status accelerates pubertal maturation (Ellis, 2004). When environments are harsh (because of non-food-related factors such as social competition) but calories are sufficient, menarche should and does occur earlier in life (see Ellis et al., 2009). The current research extends prior work by moving beyond the assessment of general stress levels or very specific stressors to examine two fundamental dimensions of environmental variation that many living organisms encounter in the service of fitness goals.
Environmental Stress Dimensions: Harshness and Unpredictability
Ellis et al. (2009) proposed that the development of life-history strategies is regulated by two environmental dimensions: harshness and unpredictability. Harshness refers to the mean level of extrinsic mortality and morbidity in a population, which in modern populations is closely tied to socioeconomic status (Belsky, Schlomer, & Ellis, 2012; Chen & Miller, 2012; Simpson, Griskevicius, Kuo, Sung, & Collins, 2012). Unpredictability refers to fluctuations in environmental conditions; in modern populations, this relates to frequent residence changes and family instability (i.e., having family members, especially paternal figures, move in and out of the home).
According to Ellis et al. (2009), exposure to harsh or unpredictable environments early in life should have unique effects on shaping life-history strategies, including accelerating pubertal timing. Recent empirical evidence about the unique effects of harshness and unpredictability on pubertal and reproductive timing, however, is mixed. Several longitudinal studies indicate that exposure to harsher or more unpredictable environments (or both) early in life does uniquely forecast fast-strategy outcomes in adolescence, such as greater delinquency and more sexual partners (e.g., Belsky et al., 2012; Brumbach, Figueredo, & Ellis, 2009; Simpson et al., 2012); some other studies find that unpredictability, as measured specifically by the absence of the father, is not uniquely associated with age of menarche (Ryan, 2015) or age at first sex (Carlson, Mendle, & Harden, 2014). Thus, the current study sought to test the unique effects of environmental harshness and unpredictability.
Moderating Effects of Early Attachment Security
Is it inevitable that exposure to stress early in life accelerates reproductive development? According to both attachment theory (Bowlby, 1973, 1988) and life-history approaches (e.g., Belsky et al., 1991; Del Giudice, 2009), the attachment bond reflects a more proximal developmental process than many distal sources of stress (e.g., poverty, absent father). The attachment bond, therefore, may be able to override the distal sources of stress, thereby buffering secure girls exposed to high levels of stress early in life and preventing them from undergoing earlier menarche.
Children develop secure attachment relationships with their primary caregiver (usually the mother) primarily in reaction to receiving responsive, situationally appropriate parental care. Children are also more likely to be secure if their caregivers have good mental health, if their parents have a happy relationship, and if sufficient social support is available (Belsky & Fearon, 2008). Because of these factors, securely attached children have greater confidence (compared with insecurely attached children) that their caregivers will provide safety, comfort, and emotional reassurance, especially in challenging or stressful situations (Ainsworth, Blehar, Waters, & Wall, 1978). Because they have received better and more consistent care, securely attached children usually adopt slower reproductive strategies across the life span (Belsky et al., 1991; Simpson & Belsky, 2008, in press). Secure children appear to learn that their caregivers can be reliably counted on for comfort and support when stressed. Accordingly, girls with insecure attachment histories experienced menarche at a younger age than secure girls (Belsky, Houts, & Fearon, 2010).
The current research extends this prior work, which has documented a main effect of attachment, to determine whether attachment moderates the effect of stress on pubertal timing. According to the model from Belsky et al. (1991), the nature and quality of the parent-child attachment relationship early in life should moderate the effect of stress on reproductive strategies and pubertal timing. Consistent with this claim, evidence from recent studies has shown that high-quality parenting protects individuals against the adverse effects of early-life stress on pro-inflammatory responses (Chen, Miller, Kobor, & Cole, 2010) and externalizing behavior problems (Skopp, McDonald, Jouriles, & Rosenfield, 2007). Thus, we propose that exposure to stress may differentially affect secure and insecure children, potentially buffering secure children from accelerated reproductive maturity.
The Current Longitudinal Study
Drawing on data from the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD) Study of Early Child Care and Youth Development (SECCYD), we address two questions: (a) Does exposure to harsher or more unpredictable environments (or both) during the first 5 years of life uniquely predict earlier onset of menarche in girls (controlling for their mother’s age of menarche)? (b) If so, are secure children protected from this stress-accelerating effect? Although it remains unclear which environmental dimension should forecast age of menarche, we predicted that girls with secure attachment histories would be protected from the adverse effects of stress.
Method
Participants
The NICHD SECCYD recruited 1,364 families through hospital visits shortly after the birth of each child in 1991 at 10 U.S. locations. For a detailed description of recruitment procedures and sample, see NICHD Early Child Care Research Network (2001). Complete data and further information are available at http://www.icpsr.umich.edu/icpsrweb/ICPSR/series/00233.
The analysis sample included 492 females for whom we had data on age of menarche. Three hundred ninety-eight were White (80.9%), 60 were Black (12.2%), 10 were Asian (2.0%), and 24 reported another ethnicity (4.9%). Compared with all other sample members, those in our analysis sample were more likely to be securely attached at 15 months of age, t(1189) = −2.50, p = .013, came from higher-income families (i.e., income-to-needs ratio), t(1353) = −2.72, p = .007, and experienced fewer paternal transitions during childhood, t(1340) = 2.34, p = .020, but did not differ on residential or parental job changes.
Measures
Environmental harshness
Following the approach of Belsky et al. (2012) and Simpson et al. (2012), we indexed environmental harshness by each family’s economic resources, a prime indicator of socioeconomic status. The economic resources of families were assessed using the income-to-needs ratio, an index of family’s income as a proportion of the federal poverty line for a family of that size. Family income was divided by the poverty threshold, adjusted for total family size. Thus, a ratio of 1 indicated that family income equaled the federal poverty threshold for a family of that size. In 1991 (the year that participants were born), the poverty threshold for a family of four was an annual income of $13,812. A higher income-to-needs ratio indicated greater financial resources per person in the household. During each participant’s first 5 years of life, the income-to-needs ratio was assessed repeatedly (when participants were 1, 6, 15, 24, 36, 54, and 60 months of age), based on his or her mother’s detailed reports of family finances.
To create an index of early environmental harshness during the first 5 years of life, we averaged income-to-needs ratios over time (α = .95). They ranged from 0.17 to 23.79 (M = 3.63, SD = 2.73). Because higher income-to-needs ratios indicate less-harsh environments, the index was reverse-scored.
Environmental unpredictability
Consistent with past research designs (e.g., Belsky et al., 2012; Simpson et al., 2012), three measures were used to assess environmental unpredictability across the first 5 years of life (see Belsky et al., 2012, and the Supplemental Material available online): (a) paternal transitions—the number of changes in the male parental figure within the home, based on face-to-face or telephone interviews about household composition when children were 1, 3, 6, 9, 12, 15, 18, 21, 24, 30, 33, 36, 42, 46, 50, 54, and 60 months old; (b) household moves—changes in residence, based on whether and when families relocated during the first 5 years of each child’s life; and (c) parental employment—the number of changes in the mother’s and the father’s or boyfriend’s employment during each child’s first 5 years, based on reports at the ages mentioned earlier. To create a scale of early environmental unpredictability, the three scores were standardized and averaged (α = .54). The unpredictability scale ranged from −1.00 to 3.39 (M = 0.08, SD = 0.68).
Infant attachment
When children were 15 months old, infant-mother attachment was assessed using the Strange Situation procedure (Ainsworth et al., 1978). Infant-mother attachment relationships were classified as secure or insecure based on how each infant responded to his or her mother after a series of brief, stressful separations. When the parent and child were reunited, secure infants usually approached their mothers, were soothed by them, calmed down quickly, and then resumed normal activity (e.g., play, exploration). Insecure infants either did not approach their mothers or displayed anger toward them, never fully calming down and not resuming normal activity during the assessment. Among our participants (children), 127 (26.8%) were classified as insecure at 15 months, and 347 participants (73.2%) were classified as secure.
Age of menarche
Age of menarche was reported at seven assessments, during which the participants were between 9 and 15 years of age. Some participants reported it at more than one assessment, and others reported it just once. When age of menarche was reported multiple times, the reports were averaged. Age of menarche ranged from 9.03 to 15.50 years (M = 12.40 years, SD = 1.13).
Maternal age of menarche
Each participant’s mother also reported her age of menarche, which was used to partially control for shared genetic effects that could account for her child’s menarcheal timing. Maternal age of menarche ranged from 9.00 to 18.00 years (M = 12.71 years, SD = 1.46).
Birth weight
The mother of each participant was interviewed within 24 hr after delivery and reported her baby’s birth weight. Participants’ birth weights ranged from 2.00 to 5.34 kg (M = 3.43 kg, SD = .49).
Results
Descriptive statistics
In Table 1, we present the relations between all of the variables. Ethnicity, which was coded 0 (non-White) or 1 (White), was significantly associated with most of the variables. Compared with non-White children, White children were heavier at birth, experienced less harsh and less unpredictable environments, and had their first menstrual cycle later. Greater harshness (i.e., lower income-to-needs ratio) was associated with greater unpredictability and earlier age of menarche. Girls classified as secure at 15 months experienced menarche later (as reported by Belsky, Houts, & Fearon, 2010). Maternal age of menarche was positively associated with girls’ age of menarche, but not with girls’ environmental harshness, unpredictability, or attachment security. Birth weight was not significantly related to any of the variables.
Correlations Between Variables in the Study
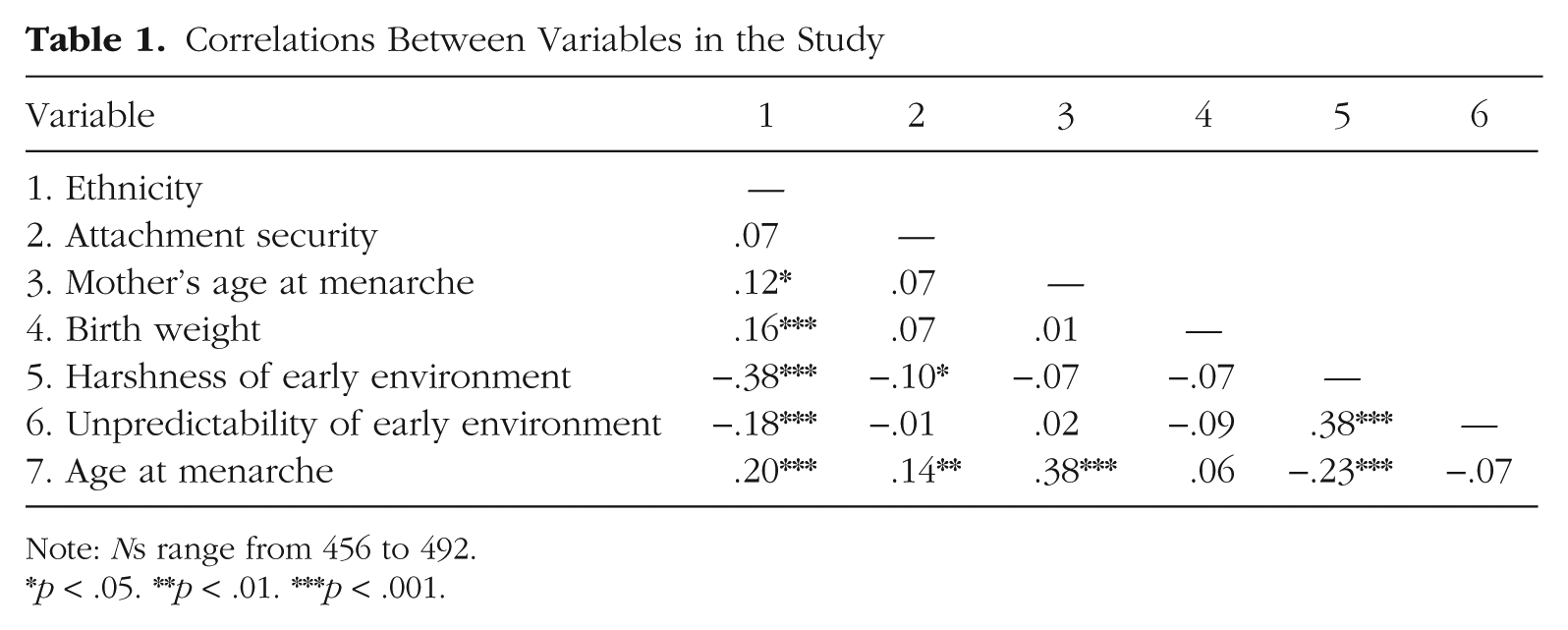
Note: Ns range from 456 to 492.
p < .05. **p < .01. ***p < .001.
Tests of hypotheses
To test (a) whether harshness or unpredictability uniquely predicted age of menarche and (b) whether infant-mother attachment security moderated these effects, we conducted a hierarchical regression analysis. The main effects of environmental harshness, unpredictability, and attachment security, along with three covariates—maternal age of menarche, ethnicity (White or non-White), and birth weight—were entered in the first block, and all two-way interactions involving the first three predictors were entered in the second block. Listwise deletion was used to deal with missing values, because Little’s missing completely at random (MCAR) test (in IBM SPSS Version 22) was positive, χ2(3, N = 421) = 5.18, p = .16. A square-root transformation was applied to the environmental-harshness and unpredictability measures, given their positively skewed distributions.
As shown in Table 2, greater environmental harshness (i.e., lower income-to-needs ratio; β = −0.26, p = .003), but not unpredictability (β = −0.10, p > .25), uniquely predicted earlier age of menarche. In addition, and as reported previously (Belsky, Houts, & Fearon, 2010), being securely attached at 15 months also predicted later menarche (β = 0.10, p = .023). Note that girls who were exposed to harsher early-life environments but who were securely attached to their mothers were buffered from accelerated reproductive development (see Fig. 1). Specifically, the Harshness × Attachment interaction was significant (β = 0.16, p = .048). Greater harshness predicted earlier menarche, but only for girls who were insecurely attached to their mothers at 15 months of age (b = −0.51, SE = 0.15, p = .003), not for girls who were securely attached (b = −0.14, SE = 0.11, p > .25).
Results of the Hierarchical Regression Analysis Predicting Age of Menarche
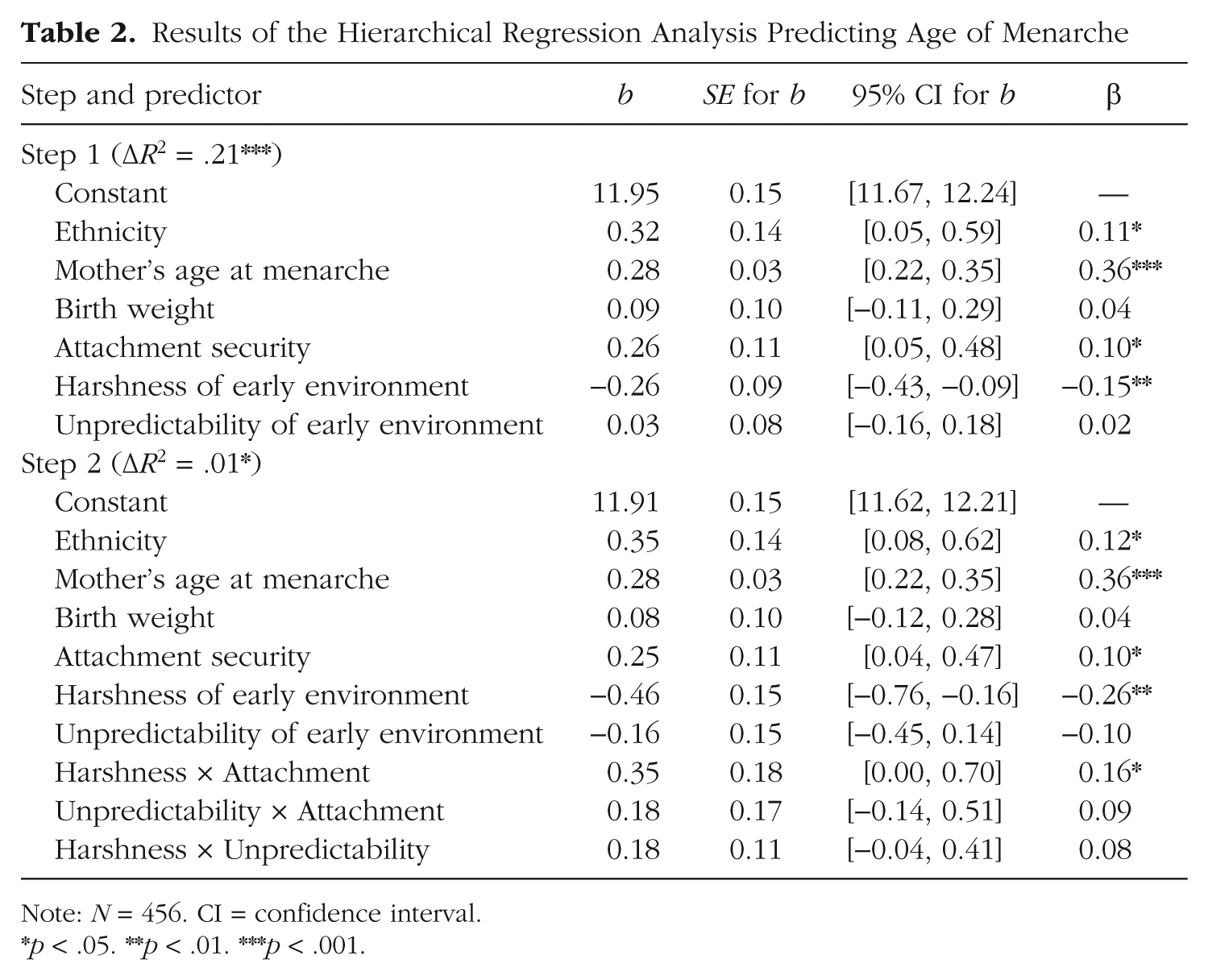
Note: N = 456. CI = confidence interval.
p < .05. **p < .01. ***p < .001.
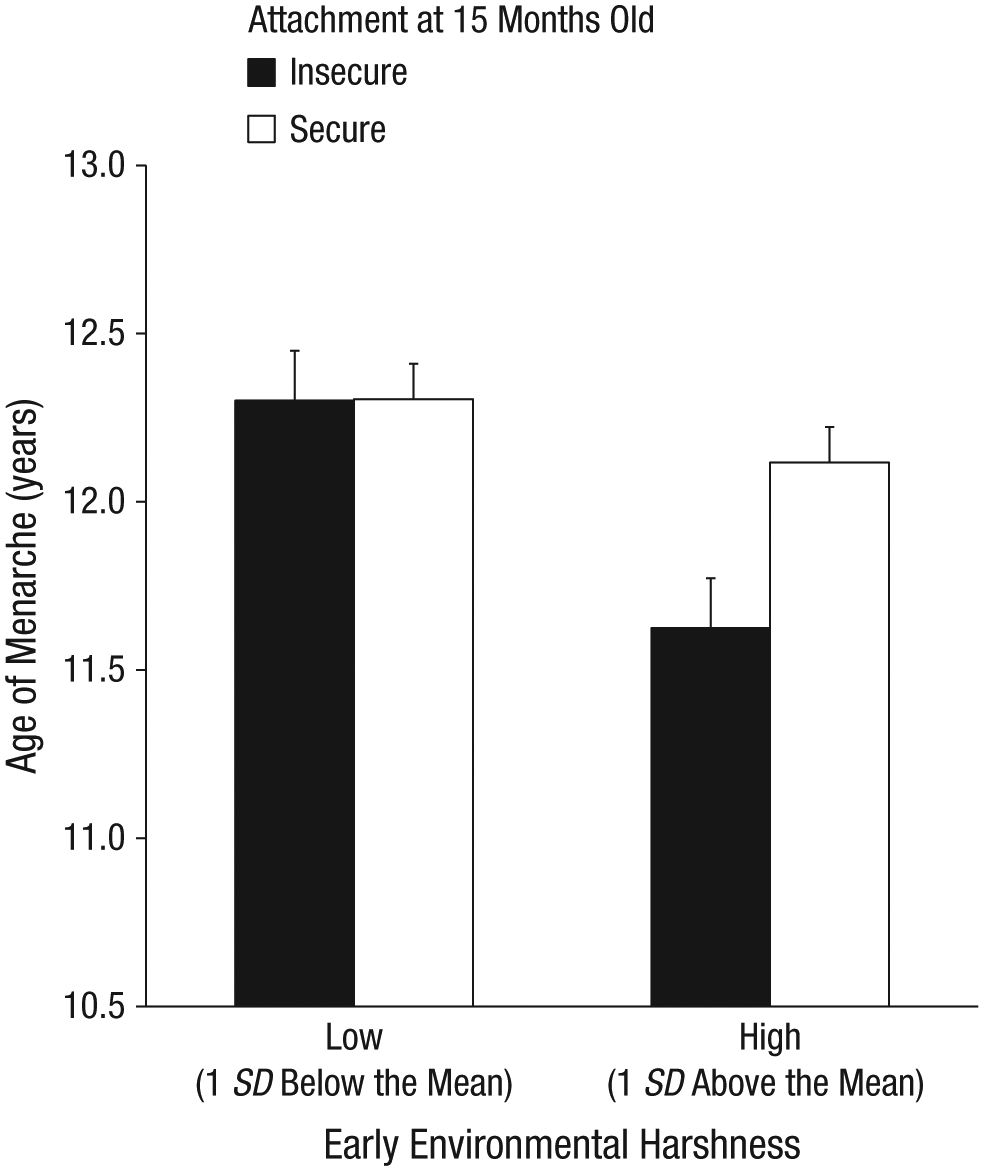
Mean age of menarche among participants who experienced low levels (1 SD below the mean) and high levels (1 SD above the mean) of early environmental harshness, graphed separately for those with secure and insecure attachments at age 15 months. Error bars indicate +1 SE.
Note that all of the effects presented here remained significant when we partially controlled for mothers’ and daughters’ shared genes (indexed by mothers’ age of menarche), which independently and positively predicted the timing of menarche. We also conducted a series of additional analyses to examine alternative models and other possible predictions (see the Supplemental Material). These analyses revealed that the effects reported earlier continued to hold when we statistically modeled and controlled for both male presence in the home and maternal sensitivity early in development (Analyses 1 and 2). Further analyses confirmed that the moderating effect of attachment security on the relation between early harshness and age of menarche was also found for latent measures of harshness and unpredictability (Analysis 3). Finally, attachment security did not mediate the link between early harshness and age of menarche, although it did moderate this connection (Analysis 4).
Discussion
Drawing on Belsky and his colleagues’ evolutionary theory of socialization (Belsky et al., 1991; Belsky, 1997) and Ellis and his colleagues’ (2009) life-history model of environmental stress, we addressed two questions in this study: Do specific dimensions of environmental stress—harshness and unpredictability—uniquely predict the earlier onset of menarche in girls? Does secure infant-mother attachment protect girls from the developmentally accelerating effect of stress?
Regarding the first question, this is the first study to show that early-life harshness—but not unpredictability—uniquely predicts the timing of menarche in girls in the theoretically anticipated manner. Greater stress stemming from a harsh environment resulted in accelerated reproductive development. It should be noted, however, that the SECCYD sample has few truly impoverished families, and Ellis et al. (2009) acknowledged that different life-history outcomes may emerge depending on the absolute level of stress to which children are exposed early in life. Future research is needed to test whether results might be different in samples containing more families that have experienced extreme unpredictability.
Regarding the second question, having a secure infant-mother attachment buffered girls from accelerated reproductive development, even when they grew up in a family with limited economic resources. This finding extends previous research chronicling the stress-protective effect of a supportive parent-child relationship (see Belsky et al., 1991; Belsky, Houts, & Fearon, 2010; Chen & Miller, 2012). In so doing, it also suggests that supportive proximal relationship processes might be able to override more distal, stressful environments.
Despite its longitudinal design, reasonably diverse sample, and good measures collected at different points of development, the current research has some limitations. First, we predicted an interactive effect of early environmental harshness and infant-mother attachment security; this effect, though statistically significant, was not large. It needs to be replicated in future longitudinal samples.
Second, our measures of harshness and unpredictability assess these constructs indirectly. We used these measures to be consistent with prior research (e.g., Belsky, Houts, & Fearon, 2010; Belsky et al., 2012; Simpson et al., 2012) and facilitate comparisons of findings with earlier studies. In future research, however, different possibilities for operationalizing both constructs should also be examined using methods that are consistent with the way in which harshness and unpredictability have been conceptualized theoretically.
Third, some prior studies have found that harshness and unpredictability both forecast certain life-history outcomes in adolescents (e.g., sexual behavior in girls; Belsky et al., 2012), whereas others have found that unpredictability alone predicts certain life-history outcomes in young adults (e.g., sexual behavior, deviance, and aggression in both sexes; Simpson et al., 2012). These different outcomes might be partly attributable to the demographic characteristics of the samples being studied (e.g., samples with higher or lower socioeconomic status), sampling error, or the indirect measurement of harshness and unpredictability. Future research should address and rectify these limitations.
Finally, in relation to the aforementioned demographic characteristics of the sample, (a) the effects of unpredictability may have been obscured by the absence of impoverished families experiencing extreme household chaos, (b) only some factors that might affect menarche were examined, and (c) the findings may have been different if other indices of reproductive maturation were studied. Future research is needed to address these limitations.
In conclusion, this research lies at the intersection of and integrates two major theories, attachment theory (Bowlby, 1973, 1988) and evolutionary models grounded in life-history theory (Belsky et al., 1991; Ellis et al., 2009). It connects attachment theory to its original theoretical roots in evolutionary biology, a foundation largely neglected over the past four decades (Belsky, 1997; Simpson & Belsky, in press), by showing how early attachment is related to reproductive development, not just psychological and behavioral development. More specifically, it illuminates conditions under which exposure to harsh environments in infancy does—and does not—accelerate female reproductive development.
Footnotes
Action Editor
Steven W. Gangestad served as action editor for this article.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This research was supported by National Science Foundation Grant 1057482 (to J. A. Simpson and V. Griskevicius).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.