
Abstract
The current study investigated how autistic traits modulate peer interactions using functional near-infrared spectroscopy (fNIRS) hyperscanning. Across three experiments, we tested the effect of copresence, joint activity, and a tangible goal during cooperative interactions on interbrain coherence (IBC) in school-aged children between 9 and 11 years old. Twenty-three dyads of children watched a video alone or together in Experiment 1, engaged in joint or self-paced book reading in Experiment 2, and pretended to play a Jenga game or played for real in Experiment 3. We found that all three formats of social interactions increased IBC in the frontotemporoparietal networks, which have been reported to support social interaction. Further, our results revealed the shared and unique interbrain connections that were predictive of the lower and higher parent-reported autism-spectrum quotient scores, which indicated child autistic traits. Results from a convergence of three experiments provide the first evidence to date that IBC is modulated by child autistic traits.
Approximately 1/100 children are diagnosed with autism-spectrum disorder (ASD) around the world (Zeidan et al., 2022). ASD is related to the overgrowth of brain volume in early childhood (Hazlett et al., 2017). Autistic individuals have shown challenges in sensory perception (Lau et al., 2021; Nayar et al., 2023; Robertson & Baron-Cohen, 2017), theory of mind (Kimhi, 2014), reading (Nally et al., 2018), and figurative-language comprehension (Kalandadze et al., 2018), which vary in individuals along a continuum of severity (Lord et al., 2018). Most autistic children go to mainstream schools, which provide normative social cultures but also introduce challenges for autistic children in their interactions with their typically developing (TD) peers in complex school environments with no special aids (Williams et al., 2019). Autistic children have been reported to have greater difficulties making peer friends in schools and higher chances of experiencing bullying and victimization compared with their TD peers (Rowley et al., 2012). The current study aimed to examine the neural mechanisms of peer interaction—the principal social interaction for schoolers—and the impacts of varying autistic traits on social synchronization.
Previous studies have used neuroimaging hyperscanning techniques—the monitoring of the brain activity of two or more people simultaneously—to examine interbrain coherence (IBC) during social interactions for autistic individuals. Those dyadic settings included autistic children and their parents (Hasegawa et al., 2016; Kruppa et al., 2021; Wang et al., 2020), ASD and TD adolescents (Key et al., 2022), and ASD and TD adults (Quinones-Camacho et al., 2021; Tanabe et al., 2012). These studies showed that children and adults with ASD had lower IBC, that is, weaker brain connections in the frontal cortex (FC) and temporoparietal area (TP), with their social partners. Individuals’ autistic traits would (Wang et al., 2020) or would not (Kruppa et al., 2021) modulate IBC depending on the tasks that were involved. Overall, these studies provide an important perspective for the study of neural functions beyond one person in ASD populations.
To date, no study has examined IBC in school-aged children with elevated autistic traits during peer interactions. The insights gained from the aforementioned research about social interactions among autistic individuals with their parents, well-trained experimenters, or confederates may not directly translate to peer interaction among school-aged children. Parents of autistic children and well-trained researchers have more experience with ASD than the average population. The limited knowledge of autism among TD children may cause unintended or intended hostility when interacting with autistic children, resulting in their social withdrawal or mental-health issues. Further, school-aged children may not be mature or capable of resolving conflicts independently, which are unavoidable during social interactions. There is still a notable gap in comprehensive information regarding how school-aged children with varying autistic traits interact with peers in normative social cultures.
The primary goal was to examine how autistic traits of school-aged children would modulate their IBC with their peers in different types of interaction using functional near-infrared spectroscopy (fNIRS) hyperscanning. fNIRS is child-friendly, motion-tolerant, and acceptable for autistic individuals. IBC was specifically examined in the bilateral frontaltemporoparietal network, which has been reported to consistently show IBC in dyadic fNIRS hyperscanning studies (for a review, see Czeszumski et al., 2022). In three experiments that established a hierarchy of nonverbal communicative settings (i.e., copresence without explicit interaction, joint activity, and turn-taking activity with explicit interaction), we tested how varying degrees of autistic traits of children modulated IBC.
The mere presence of another individual will increase arousal and social facilitation (Guerin, 1986; Platania & Moran, 2001), with information exchanging between brains through neural oscillation (Holroyd, 2022), resulting in altered IBC. This phenomenon was observed in a previous fNIRS hyperscanning study (Miura & Noguchi, 2022) that reported increased IBC in the left FC (lFC) between dyads of adults when they performed self-paced addition tasks together compared with when they performed these tasks alone. The mere presence effect on the IBC between dyads of children with varying autistic traits remains unknown. To bridge this gap, Experiment 1 tested the hypothesis of the mere presence effect on IBC by asking children to watch a video together or alone in separate sessions, which ruled out neural entrainment to visual stimuli (Spaak et al., 2014). We further hypothesized that the increase in IBC would be modulated by the autistic traits of children, such that the mere presence of a peer may have a minimum influence on children exhibiting severe autistic traits compared with children with milder traits (Burnside et al., 2017).
Statement of Relevance
Peer interaction is the primary social interaction for schoolers. This study examined the neural mechanisms underlying peer interaction among school-aged children between 9 and 11 years old with elevated autistic traits. Peer interaction was quantified by monitoring the brain activity of two children in a dyad simultaneously to calculate their interbrain connections. In three experiments, we demonstrated the effect of copresence, joint activity, and a tangible goal during cooperative interactions on interbrain and intrabrain connections in the attention networks between dyads of children. Results from a convergence of three experiments also provide the best evidence to date that interbrain connection is modulated by child autistic traits, which was supported by the altered intrabrain connections in individuals.
Experiment 2 tested how joint activity affected IBC by asking children in the dyad to read a book and vocalize in one session but subvocalize in another session. Aligned joint reading could be achieved through the continuous allocation of attention to the partner and self-adjustment of own paces (Smallwood & Schooler, 2015), or leader-follower harmony (Jiang et al., 2015; Sänger et al., 2012), with the responder following the lead of the initiator. We hypothesized that either mechanism above during joint reading would increase the IBC and intrabrain coherence (IntraBC) in the frontotemporoparietal network compared with the subvocalization session when children read at their own pace and may neglect others’ progress. However, the difficulty in reading and comprehension among children who were more autistic (Nally et al., 2018) may hinder their ability to align with their peers, hence resulting in smaller differences in IBC between the two reading conditions, compared with children who were less autistic.
Experiment 3 examined how autistic traits affect IBC during nonverbal turn-taking activities by asking children to pretend to play a Jenga game or play for real. Playing a Jenga game requires cooperative activities and theory of mind and has been shown to synchronize two TD adults (Liu et al., 2016) in the right FC (rFC). Pretend play requires imagination and the ability to conceive nonliteral situations, which has been challenging for autistic children (Jarrold, 2003). We tested the hypotheses that a tangible goal in the real-play session would better synchronize two brains, resulting in greater IBC compared with the pretend play, and that the increase in IBC would be modulated by children’s autistic traits.
The second goal was to examine whether social interactions across the three experiments would activate distinguished interbrain networks for children with lower or higher autistic traits in each dyad. To shed light on the specific autistic traits associated with interbrain connections, we further examined the relations between interbrain connections and varying subcategories of autistic traits, including attention to detail, imagination, mentalization, and social skills. In exploratory analyses we investigated whether the altered interbrain networks would correspond with activation in intrabrain networks in individuals. An improved understanding of this topic would enable the prediction of children with severe autistic traits using fNIRS hyperscanning and provide suggestions for peer-mediated intervention strategies that target specific subcategories for children with autistic traits but who have not been diagnosed.
Method
Participants
Forty-six children 9 to 11 years of age who were attending mainstream primary schools participated in this study in dyads (n = 23). According to a meta-analysis by Czeszumski et al. (2022), previous fNIRS hyperscanning studies that examined cooperative activations yielded a large effect size (Hedge’s g = 1.98). We determined that a sample size of 23 was sufficient to reveal significance (p < 0.05) with a power of 0.95. We included children who were (a) 9 to 11 years old at the time of testing and attending mainstream schools, (b) TD or diagnosed with ASD by a medical provider but without any other known neurodevelopmental conditions, and (c) with no observable difficulties that would prevent them from participating in the study (e.g., hostile/unfriendly to other peers with varying traits, unable to cooperate with others or concentrate to activities for a reasonable time, or incapable of book reading). Parents of all children completed an online survey and provided their basic demographic information. Thirty-two children (eight females) with a mean age of 9.9 years (SD = 1.0) were reported to be TD by their parents. Fourteen children (three females) with a mean age of 10.4 years (SD = 1.1) were reported to have been diagnosed with ASD. Parents also prefilled out a 50-item autism-spectrum quotient (AS-Q) questionnaire (Auyeung et al., 2008) that was developed to detect autistic traits among children between 4 and 11 years of age. Because our study did not concern ASD as a diagnosis but how autistic traits may modulate IBC, only autistic traits were assessed, and an ASD diagnosis was not ascertained beyond parental report. All dyads of children were matched in age; all but two dyads were matched in sex. One dyad among the two that were not matched in sex resulted from a mistake in the parent report of the child’s sex in the online survey. The other dyad was due to the researcher’s error during scheduling. On the day of testing, one parent of each child participant also filled out a 36-item Broad Autism Phenotype Questionnaire that was aimed to quantify the autistic traits of both parents (Hurley et al., 2007). The demographic information of child participants and their parents is shown in Table 1.
Demographic Information

Note: AS-Q scores were from an AS-Q questionnaire (child version) filled out by parents to quantify children’s autistic traits. BAPQ scores were from a 36-item BAPQ (adult version) filled out by one parent that was aimed to quantify parents’ autistic traits. TD = typically developing; ASD = autism-spectrum disorder; AS-Q = autism-spectrum quotient; BAPQ = Broad Autism Phenotype Questionnaire; HKD = Hong Kong dollars.
Experimental conditions
Children completed three experiments (six sessions in total in a counterbalanced order across dyads) in one visit in a sound-treated booth (Fig. 1). In Experiment 1, two children sat on rolling chairs side by side and watched a silent video (3 min long) about a junior Jenga competition (see the Supplemental Material available online). Children in a dyad either watched this video together (VoTo) or alone (VoAl) with their counterparts leaving the room. To keep their attention, children were told to role-play one of the players and think about whether each move was smart, risky, or inappropriate and to avoid articulation. Twenty-two dyads of children completed Experiment 1; one dyad (Dyad 12) completed only the VoTo session—hence, their data in Experiment 1 were not included in further analyses. To test the mere presence effect, we complied with the rule by asking children to perform the same tasks in the VoAl and VoTo sessions by not involving evaluation of the other part and by not involving the presence of an experimenter (Guerin, 1986). By recruiting children who had never met each other, this design represented real-world social-interaction situations and avoided the confound of closeness between two parts in a dyad on IBC (Balters et al., 2023; Bevilacqua et al., 2019; Bizzego et al., 2020).

Paradigms of the testing settings. Panel (a) shows a schematic paradigm for Experiment 1, in which two children watched a video of two teenage professionals playing a Jenga game alone or together. Panel (b) shows a schematic paradigm for Experiment 2, in which two children jointly read a book and were instructed to subvocalize and vocalize in two separate sessions. Panel (c) shows a schematic paradigm for Experiment 3, which involved two children playing or pretending to play a Jenga game in two sessions. All the cartoon characters underwent modifications using resources purchased online for noncommercial use.
In Experiment 2, children were asked to read a storybook about idioms together in their native language (i.e., Cantonese). The storybooks were written in traditional Chinese corresponding to the intermediate language in their school textbooks and are recommended for students in middle and high school. Children were told to read a story of their choice and subvocalize (SubVo) in one session (4.5 min long) and read a different story and vocalize (Vocal) in the other session. Communication between the dyad was encouraged in the Vocal session if they had difficulties in recognizing some Chinese characters or understanding the stories; in the SubVo session, children were also told to use gestures and eye gaze if communication was needed with their counterparts. Twenty-one dyads of children completed Experiment 2; two dyads (Dyads 16 and 18) did not participate or completed only one session in Experiment 2—hence, their data in Experiment 2 were not included in further analyses.
In Experiment 3, children were told to play Jenga with a goal of not letting the tower fall (Play) in one session (4.5 min long) and to pretend to play a Jenga game and using the same gestures but without physical contact with the wooden sticks (Pret) in the other session. By comparing the real and pretend play of Jenga, this design avoided the confounding motor synchrony because both sessions involved Jenga-related turn-taking movements of the two players (Holroyd, 2022) and allowed the test of the hypothesis that a tangible and shared goal could better synchronize two brains compared with an intangible goal. Twenty-two dyads of children completed Experiment 3; Dyad 16 completed only the Play session—hence, their data in this experiment were not included for further analyses. More detailed information about the experimental setup is available in the Supplemental Material.
fNIRS data acquisition and analyses
Data were collected in a sound-treated booth using a continuous-wave NIRScout system (NIRx Medical Technologies, LLC, Orlando, FL) with a sampling frequency of 7.81 Hz. During hyperscanning, 16 light-emitting diodes that emitted near-infrared light of two different wavelengths (760 and 850 nm) and 24 avalanche photodiode (APD) detectors were split into two sets and arranged to cover the frontotemporoparietal areas on both hemispheres of the two participants (Fig. 2). To reduce the noise in fNIRS data such as systemic responses from the extracerebral tissue (Tachtsidis & Scholkmann, 2016), short channels were included that have been shown to greatly improve fNIRS signal quality in adults (Zhou et al., 2020) and young children (Zhou et al., 2021, 2023). The last APD detectors for each participant in a dyad were connected to a bundle of eight short-channel detectors that were then connected to eight light sources to produce 8-mm short channels. The optodes were held by an elastic NIRScap of predetermined size based on each child’s head circumference. Among all children, the cap sizes used included 54, 56, and 58 cm; we sized up by 1 to 2 cm to avoid the snug fit of the blunt tips of NIRx optodes that makes children uncomfortable. The locations of light sources and detectors were arranged to the standard 10–10 system (Fig. 2). In total, 19 regular channels (approximately 30 mm long) were used per participant for fNIRS signal recording, which were then divided into four brain regions of interest (ROIs), that is, the lFC, rFC, left TP (lTP), and right TP (rTP).
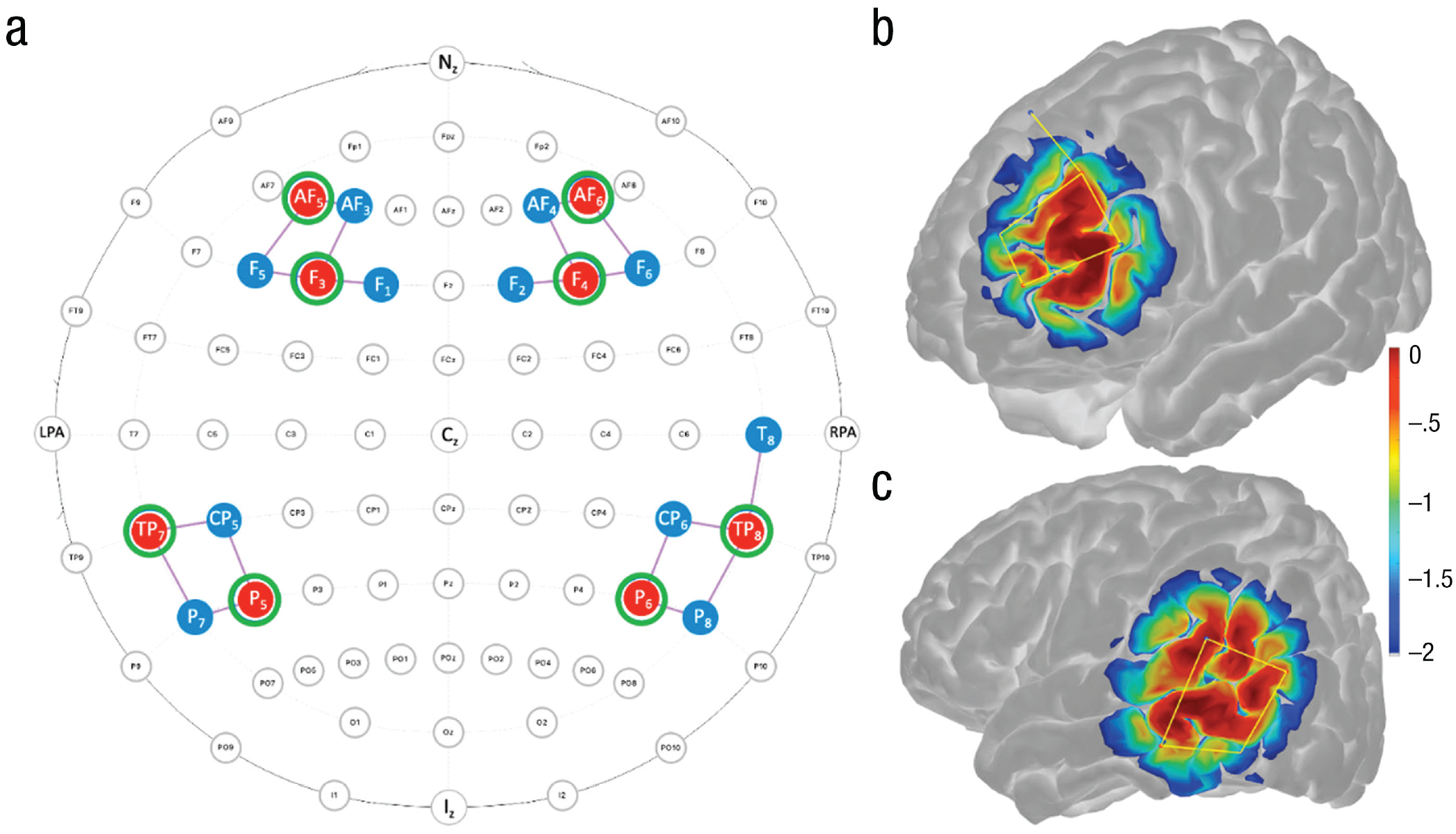
fNIRS montages. Panel (a) shows the montage, that is, the locations of and connections between fNIRS light sources (red; n = 8), regular light detectors (blue; n = 11), and short-channel detectors (green; n = 8), which was created with NIRSite Version 2.0. Panels (b) and (c) show the sensitivity maps (with a log10 unit of mm−1) of near-infrared light in measuring local chromophores in the regions of interest in the left frontal cortex and left temporoparietal area, respectively. The more reddish color refers to better sensitivity; the map was generated using the AtlasViewer toolbox (Aasted et al., 2015). fNIRS = functional near-infrared spectroscopy.
The fNIRS data were imported into and analyzed in MATLAB (The MathWorks, Natick, MA) with scripts from HOMER2 software (Huppert et al., 2009) and written by the authors to preprocess data, exclude channels of poor data quality, and denoise (Zhou et al., 2020). To reject channels of poor quality, the correlation between heartbeat signals recorded from the two wavelengths for each fNIRS channel, that is, the scalp coupling index (SCI), was calculated (Pollonini et al., 2016). Channels with an SCI lower than 0.25 were excluded from further analyses. This cutoff threshold was set to ensure that at least four short channels were included for denoising to obtain robust results for short-channel subtraction analyses, as recommended by Sato et al. (2016). To avoid step-like noises and spikes in signals that would result in high SCI values, we included one step to denoise before calculating the SCI for each channel (for details, see Zhou et al., 2020). Among a total of 19 regular channels across all the participants, the mean number of channels included for further analyses was 16.6 (SD = 2.2). Among a total of eight short channels, the mean number included for further analyses was 6.7 (SD = 1.3). For the numbers of regular and short fNIRS channels that were included for signal processing in individuals in each session, see Figs. S1 and Fig. S2 in the Supplemental Material.
After excluding channels of poorer signal quality, the preprocessing pipeline on the regular channels included for further analyses consisted of (a) converting light intensity to optical density, (b) reducing motion artifacts using a wavelet analysis method, (c) calculating the concentration changes in the hemoglobin with children’ ages taken into account (Scholkmann & Wolf, 2013), (d) filtering the low and high noise with a bandpass (0.01–0.5 Hz) filter, and (e) reducing the noise from the extracerebral tissue using a general linear model and principal component analysis combined method (for details, see Zhou et al., 2020). We selected a bandpass filter that excluded signals above 0.5 Hz that mainly correspond to heartbeats because the study did not concern such physiological signals.
After preprocessing, the IBC in each dyad between the child with higher and lower AS-Q scores was measured by calculating the wavelet transform coherence (WTC) for the change in oxygenated hemoglobin signals, following recommendations from a previous study (Nguyen et al., 2021). IBC was calculated because we were interested in how well changes in brain activity in one person in a dyad align with changes in brain activity in the other person when they were exposed to the same environmental settings or performing the same tasks. To mark interbrain ROI connections, we used ROIH-ROIL to stand for the connection between the ROI in the child with higher (ChildH) and lower (ChildL) total AS-Q scores (Fig. 2). The WTC measures cross-correlations between two signals in the frequency and time domains that uncover locally phase-locked components between two signals and has been widely used to reveal IBC (Cui et al., 2012; Nguyen et al., 2021). To identify the frequency of interest for calculating IBC, we did a band-by-band analysis (Pan et al., 2018) in the frequency domain between 0.015 and 0.5Hz, with a bandwidth of 0.016 Hz and a total of 60 frequency bands. Between each pair of channels for each frequency band, the coherence values across the time domain were then averaged. To calculate the IBC for each pair of ROI connections for each frequency band, for instance, ROIH-ROIL, the IBC values between the channels of good data quality within ROIH of ChildH and the channels within ROIL of ChildL were calculated for that frequency band and then averaged. For each dyad of children with higher and lower AS-Q scores each with four ROIs, there were 4 × 4 × 60 IBC values. One-tailed paired t-tests were conducted on the IBC values (n = 960) between the two sessions in each experiment across all dyads. For each pair of interbrain ROI connections in each experiment, frequency ranges of interest were identified as no less than three consecutive frequency bands among 60 that showed significance from the t-tests (p < .05) to control for false positives. For pairs of ROIs that frequency ranges of interest were identified in any of the three experiments, the WTC values were recalculated. Similar to the calculation of IBC, the IntraBC for the ROIs in all the individuals was calculated to understand how social interactions and autistic traits affect intrabrain connections in the frontotemporoparietal networks.
Statistical analysis
We hypothesized that copresence, joint activity, or a tangible goal could increase the IBC in dyads of school-aged children and that the increases in the IBC in each experiment would be correlated with the autistic traits of children in each dyad. A few steps were involved to test these hypotheses. First, one-tailed paired t-tests were conducted on the IBC values between the two sessions in that experiment for each pair of ROIs that showed a significant frequency band of interest in any of the three experiments. Second, the Benjamini-Hochberg method (Benjamini & Hochberg, 1995) was used to control the false discovery rate (FDR) for the p values among a maximum of 4 × 4 ROI connections that showed significant frequency bands in each experiment. Third, among the ROIs that showed significance, correlational analyses were conducted to examine how autistic traits of children would modulate the effect of each of the three types of social interactions on IBC. Correlations were examined between the increases in the IBC between the two experimental conditions (i.e., ΔIBC) and the higher and lower AS-Q total scores and the differences in the AS-Q total scores between two children in each dyad. These correlations were unadjusted because corrections for multiple comparisons were applied in the second step to identify the ROIs.
To test our second-tier hypothesis that children with higher or lower autistic traits in each dyad would activate distinct brain networks during peer interactions, we built separate linear regression models using iterative reweighted least squares (Holland & Welsch, 1977) among interbrain ROI connections that showed significant increases in the IBC to predict the higher and lower child AS-Q scores. To avoid overfitting the model and collinearity between variables, the differences in the IBC between two conditions in ROI connections identified from Experiment 1 were first entered into the model because copresence established the ground of social interaction. After identifying the combinations of ROI connections in Experiment 1 that best predicted the AS-Q scores, ROI connections identified in Experiments 2 and 3 were entered at the same time. The best-fit models to predict the higher and lower AS-Q scores were determined on the basis of the estimated error and adjusted R2 values. Similar to the regression analysis on the IBC results, linear regression models on the IntraBC results for AS-Q scores in individuals were built to understand how child autistic traits affect their intrabrain connections during peer interactions.
Results
Parent-report child AS-Q scores
The autistic traits of children were assessed on the basis of parent-reported AS-Q scores completed pretesting, with higher scores suggesting more autistic (Auyeung et al., 2008). With a cutoff threshold of 76 within the range of 0 to 150, AS-Q total scores have been reported to have high sensitivity (95%) and high specificity (95%) in determining children with ASD. Figure 3 plots parent-reported AS-Q scores of TD children (blue; n = 32) and children with ASD (red; n = 14) diagnosed by a medical provider. The group mean of the total AS-Q scores of TD children was significantly greater (t = 7.49, p < .001) than that in the TD group in Auyeung et al. (2008). Eleven of 14 children with ASD had AS-Q scores above 74 (i.e., near the cutoff threshold of 76). However, the group mean of parent-reported AS-Q scores of TD children, 75.23 (SD = 10.82), in the current study was neither significantly lower than 76 nor lower than the ASD group, t(44) = −1.23, p = .23). Half were above the ASD cutoff threshold, suggesting high autistic traits of these children (that parents may not be aware of and hence not seeking medical diagnosis). We adopted the assessment of autistic traits in the four subcategories for the AS-Q parent-report questionnaire as described in Auyeung et al. (2008), which are attention to detail, imagination, mind reading, and social skills; each subcategory has demonstrated high test–retest reliability. The ASD group showed higher scores in the categories of imagination, t(44) = 1.70, p = .048, and social skills, t(44) = 3.58, p < .001, but not in the categories of attention to detail, t(44) = −0.92, p = .36, or mind reading, t(44) = −0.87, p = .39, compared with the parent-reported TD group. Overall, because of our mere interest in the range of autistic traits as a group when examining how these traits modulated IBC during peer interactions and the lack of differences in AS-Q total scores, we no longer use the terms “ASD” and “TD” to refer to these children from here on (see also the Supplemental Material).

Parent-reported AS-Q scores of children. The top row plots the total AS-Q scores of TD children (blue) and children with medically diagnosed ASD (red). The cyan vertical dashed line (AS-Q = 76) indicates the ASD cutoff (Auyeung et al., 2008). Higher scores suggest more severe autistic traits. The bottom row plots AS-Q scores for the four subcategories (i.e., attention to detail, imagination, mind reading, and social skills). AS-Q = autism-spectrum quotient; TD = typically developing; ASD = autism-spectrum disorder.
We further examined the AS-Q total scores and scores in four subcategories of children in dyads. Figure 4 (left) plots the higher and lower AS-Q total scores of children in each dyad. Across dyads, children with higher and lower AS-Q total scores in each dyad, denoted as AS-QH and AS-QL, respectively, showed significant differences as a group, t(22) = 4.91, p < .001. Across dyads, children with higher and lower AS-Q total scores also showed significant differences in the subcategories of imagination, t(22) = 2.12, p = .023, mind reading, t(22) = 1.74, p = .048, and social skills, t(22) = 3.53, p < .001, with no significant differences in the subcategory of attention to detail, t(22) = 0.43, p = .33.
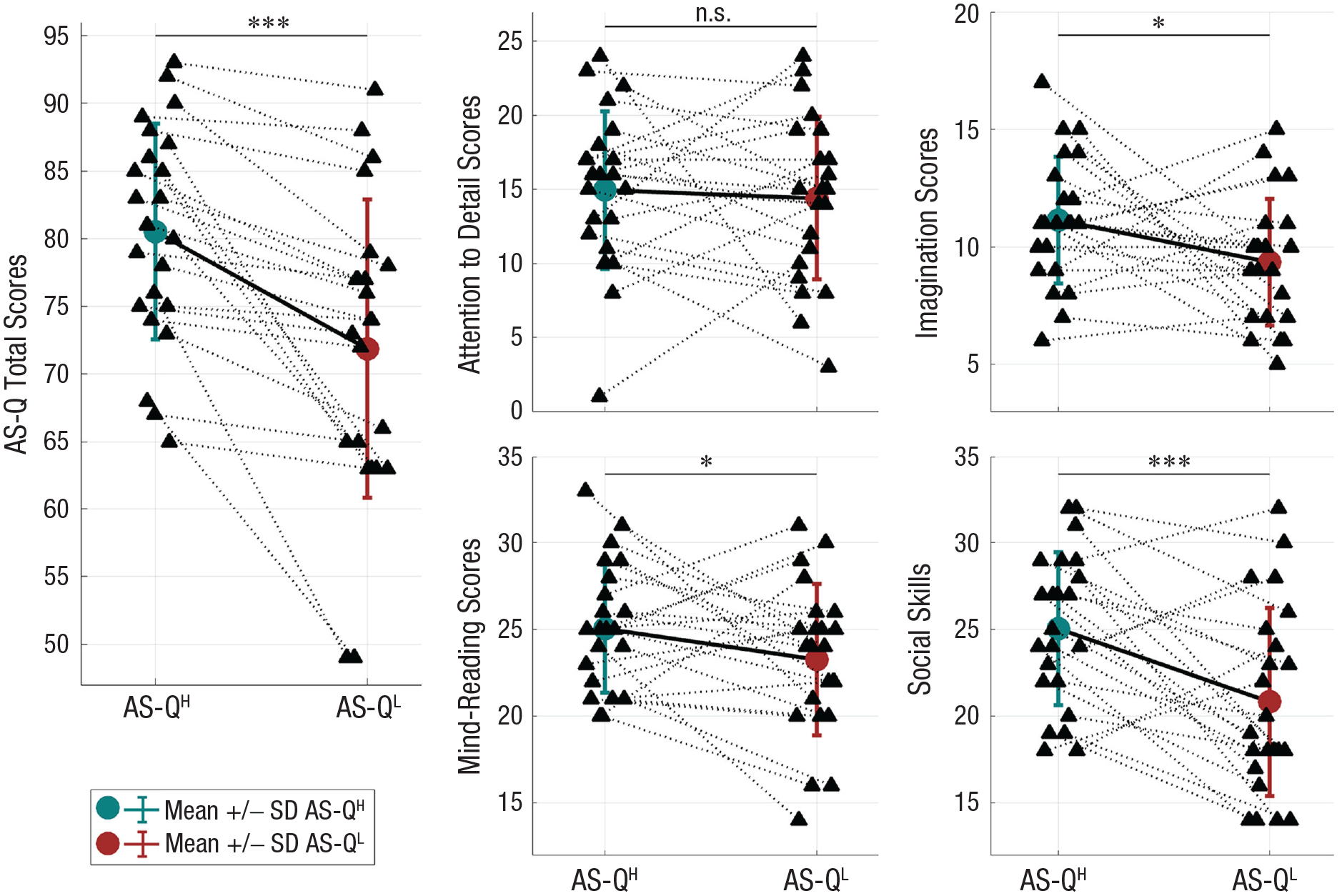
The higher and lower AS-Q scores of children in each dyad. In each dyad, we note the child with higher (teal) and lower (brown) AS-Q total scores (i.e., AS-QH and AS-QL, respectively). For children with lower and higher AS-Q total scores, we further examined their scores in four subcategories: attention to detail, imagination, mind reading, and social skills. Each panel plots the means (dots) and standard deviations (bars) of the higher and lower scores. Asterisks indicate significant differences between the two groups (*p < .05. ***p < .001). AS-Q = autism-spectrum quotient; n.s. = not significant.
ROI connections that showed increased IBC in each experiment
For each dyad, we examined the IBC in four ROIs: the lFC, rFC, lTP, and rTP. For fNIRS measures, we expected increased IBC because of three mechanisms. The first was the IBC in the low-frequency range because of slow changes in cortical hemodynamic responses, which are associated with neuronal activities through neurovascular coupling. The second was increased IBC in the middle-frequency range (approximately 0.2–0.3 Hz) because of the synchronized respiration (12–18 beats) between two participants in a dyad. The copresence of their peer (during video watching) and/or joint vocalization (during book reading) may alter their physiology to achieve such behavioral synchrony. Third, during Jenga playing, we expected increased IBC in the relatively higher frequency range because of synchronized motor activities, stemming from their cooperative turn taking, mirroring their partners’ movement, and engagement in the task. Table 2 summarizes the interbrain ROI connections that showed significant increases in IBC after controlling the FDR for multiple comparisons.
Summary of ROI Connections That Showed Significant IBC Results in Three Experiments

Note: From left to right, the columns show the experimental conditions, the ROI connections that showed significant differences in IBC across dyads between the two conditions, the frequency ranges that revealed the differences in IBC, the adjusted p values for multiple comparisons corrections using the FDR method, and correlations with AS-QH and AS-QL and differences between AS-QH and AS-QL (i.e., ΔAS-Q scores). ROI = region of interest; IBC = interbrain coherence; AS-Q = autism-spectrum quotient; VoTo = video together; VoAl = video alone; Vocal = vocalize; SubVo = subvocalize; Play = real play; Pret = pretend play; lFC = left frontal cortex; rFC = right frontal cortex; lTP = left temporoparietal area; rTP = right temporoparietal area; FDR = false discovery rate; H and L = higher and lower scores, respectively.
In Experiment 1 (video watching), across dyads of children, increased IBC from watching a video alone (VoAl) to together (VoTo) were found primarily in the following interbrain ROI connections: lFCH-rFCL, lFCH-lTPL, lTPH-rFCL, rFCH-lFCL, rTPH-lFCL, and between the rTPH and all the four ROIs of ChildL. The increased IBC was identified in the low frequencies (0.018–0.13 Hz) that likely correspond to the coherence of changes in neuronal responses and in the middle frequencies that may correspond to synchronized respiration in the VoTo session (0.154–0.276 Hz). Among the ROI connections that showed significance, the increases in IBC resulting from copresence in some were also correlated with the AS-QH, AS-QL, or ΔAS-Q total scores (Table 2). It is worth noting that the ΔIBC in the lTPH-rFCL showed positive correlations with the AS-QH and ΔAS-Q total scores, suggesting that when the AS-Q scores of the child with higher scores (more autistic) in each dyad were higher, and when the differences in the AS-Q total scores between two children were greater, the increases in the IBC resulting from the copresence between dyads of children were greater.
In Experiment 2 (book reading), increased IBC from self-paced subvocalization (SubVo) to joint vocalization (Vocal) sessions were shown between the lFCH and the four ROIs of ChildL, between the rFCH and rTPH and bilateral TPL, and the lTPH-rFCL. The increased IBC was mostly identified in the middle-frequency range (0.13–0.311 Hz), suggesting synchronized respiration between children in dyads during joint and vocalized reading compared with during subvocalized and self-paced reading. Among the ROI connections that showed significance, the increases in IBC resulting from joint book reading in some were also correlated with the AS-QH, AS-QL, or ΔAS-Q total scores (Table 2). The ΔIBC in the lTPH-rFCL in the low and middle frequencies showed significant and negative correlations with the AS-QH and AS-QL scores and a positive correlation with the ΔAS-Q scores, suggesting that when the AS-Q scores of either child were higher or when the differences in AS-Q scores between two children were smaller in each dyad, the increases in IBC resulting from joint activity between dyads of children were smaller.
In Experiment 3 (Jenga playing), increased IBC from pretend-play and play sessions was shown in these ROI connections: lFCH-rTPL, lTPH-rTPL, and rTPH-lFCL. The increased IBC was observed in the middle-frequency range (0.154–0.219 Hz), which could again be due to respiration, and in a relatively higher frequency band (0.29–0.35 Hz), which could be related to increased motor synchrony in a real game compared with a pretend-play session. The correlational analyses revealed no significant correlations between the ΔIBC and AS-Q scores, with a nonsignificant correlation between the ΔIBC in the lTPH-rTPL connection and the AS-QL scores (p = .08).
To summarize, all three social interactions increased IBC between dyads of children. Further, varying autistic traits of children were associated with the increases in IBC resulting from copresence during video watching and joint activity during book reading both in the lTPH-rTPL connection. However, no such correlation was found between the increases in IBC resulting from a tangible goal while playing Jenga games and child autistic traits.
IBC regression models predicting child AS-Q scores
We tested our second-tier hypothesis that children with higher or lower autistic traits in each dyad would activate distinct brain networks during peer interactions. Among the interbrain ROI connections that showed significant increases in IBC because of social interactions in each experiment (Table 2), regression analyses on the differences in the ΔIBC across experiments to predict the higher and lower AS-Q total scores of children in each dyad revealed two models, that is, AS-QH ~ IBC Model 1 and AS-QL ~ IBC Model 2. AS-QH ~ IBC Model 1 (Fig. 5c) predicts the higher AS-Q total scores (more autistic) of children in each dyad (Fig. 5a; R2 = .6, p = .006) along with the AS-Q scores in the subcategory of social skills (Fig. 5b). AS-QL ~ IBC Model 2 (Fig. 5f) predicts the lower AS-Q total scores (less autistic) of children in each dyad (Fig. 5d; R2 = .84, p < .001) along with the AS-Q scores in the subcategory of attention to detail (Fig. 5e). AS-QL ~ IBC Model 2 also predicts the AS-Q scores in the subcategories of imagination (R2 = .34, p = .078) and mind reading (R2 = .35, p = .073), although the results were not significant. These results suggest that between children who had higher or lower AS-Q total scores, indicating more or less autistic, the ΔIBC values, although interacting with each other, were associated with distinct subcategories of autistic traits. Finally, we explored whether the increases in IBC resulting from social interactions would predict the differences in AS-Q scores between two children in each dyad. This analysis revealed the third model, that is, ΔAS-Q ~ IBC Model 3, which predicts differences in AS-Q total scores (Fig. 5g; R2 = .69, p = .002) and differences in the subcategory of imagination (Fig. 5h) between ChildH and ChildL in each dyad.
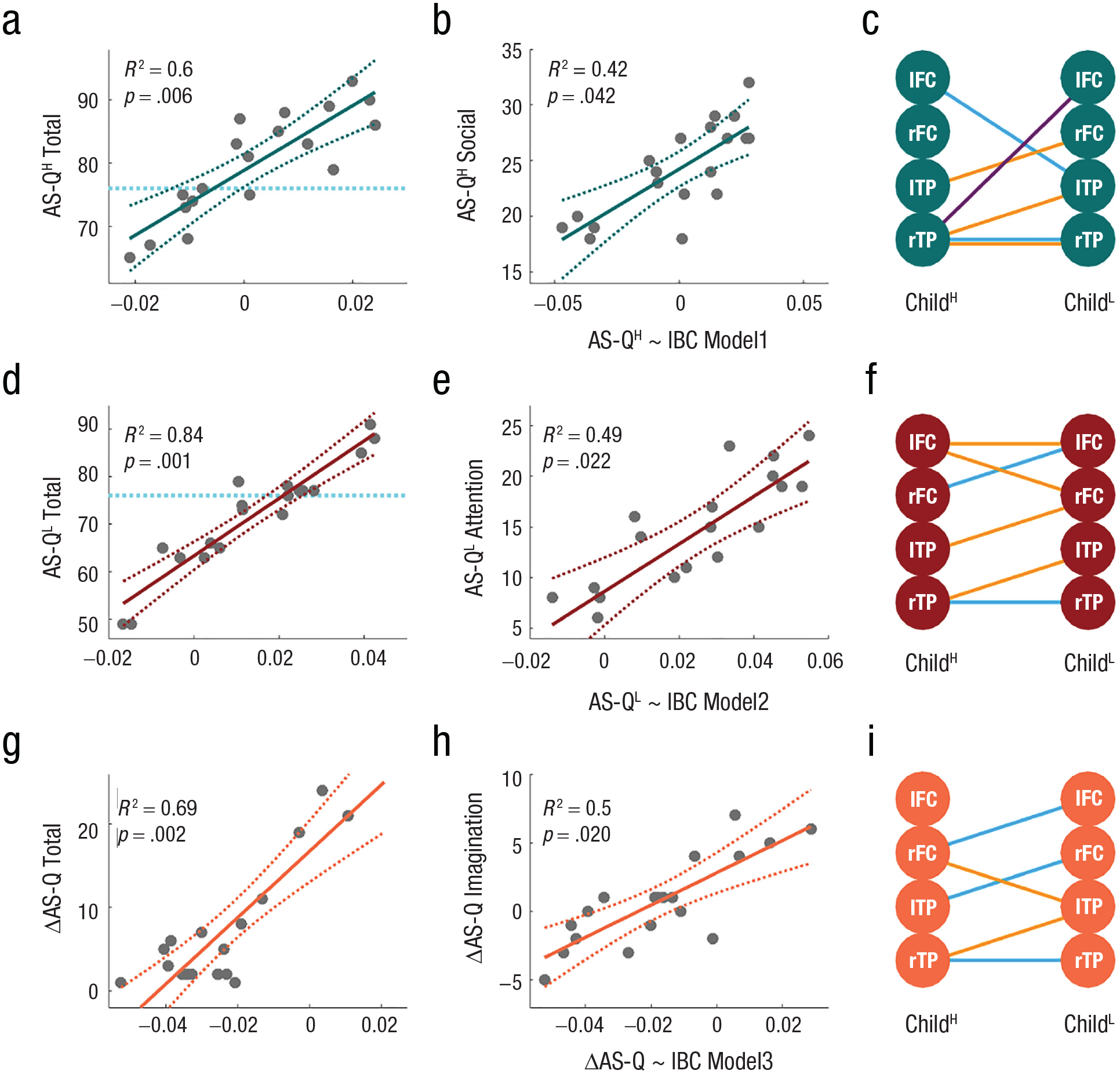
Regression models of IBC for child AS-Q scores. Panels (a) and (b) plot the regression models of differences in IBC between the two conditions in each experiment (i.e., ΔIBC) for AS-Q total scores and scores in the subcategories of social abilities for children with higher scores (more autistic) in each dyad (i.e., AS-QH ~ IBC Model 1; teal). Panel (c) plots the ROI connections between children with higher and lower AS-Q scores (i.e., ChildH and ChildL, respectively), identified in the video watching (cyan), Jenga playing (purple), and book reading (orange), which consisted of AS-QH ~ IBC Model 1. Panels (d) and (e) plot the regression models of the ΔIBC for AS-Q total scores and the scores in the subcategory of attention to detail for ChildL (less autistic) in each dyad (i.e., AS-QL ~ IBC Model 2). Panel (f) plots the ROI connections that consisted of AS-QL ~ IBC Model 2. Panels (g) through (i) plot the regression models of the ΔIBC for the differences in AS-Q scores between ChildH and ChildL in each dyad. IBC = interbrain coherence; AS-Q = autism-spectrum quotient; ROI = region of interest.
Further comparisons of AS-QH ~ IBC Model 1 and AS-QL ~ IBC Model 2 found unique (Figs. 6a and c) and shared interbrain ROI connections (Fig. 6b) that contributed to the models. The shared components consisted of the lTPH-rFCL and rTPH-lTPL connections during book reading in the middle-frequency ranges and the rTPH-rTPL connection during video watching and book reading in the low- to middle-frequency ranges (Fig. 6 and Table 2). The unique ROI connections in AS-QH ~ IBC Model 1 consist of the lFCH-lTPL connection during video watching and the rTPH-lFCL connection during Jenga playing, with stronger IBC in the middle-frequency range (0.15–0.22 Hz), whereas the unique ROI connections in AS-QL ~ IBC Model 2 consist of connections between the lFCH and the bilateral FCL in the middle-frequency range (0.15–0.31 Hz) during book reading and the rFCH-lFCL connection in the low-frequency range (0.025–0.031 Hz) during video watching. Overall, these results suggest that children with higher or lower AS-Q scores (i.e., being more or less autistic, respectively) may contribute to IBC differently, with different networks being involved for varying types of peer interactions.
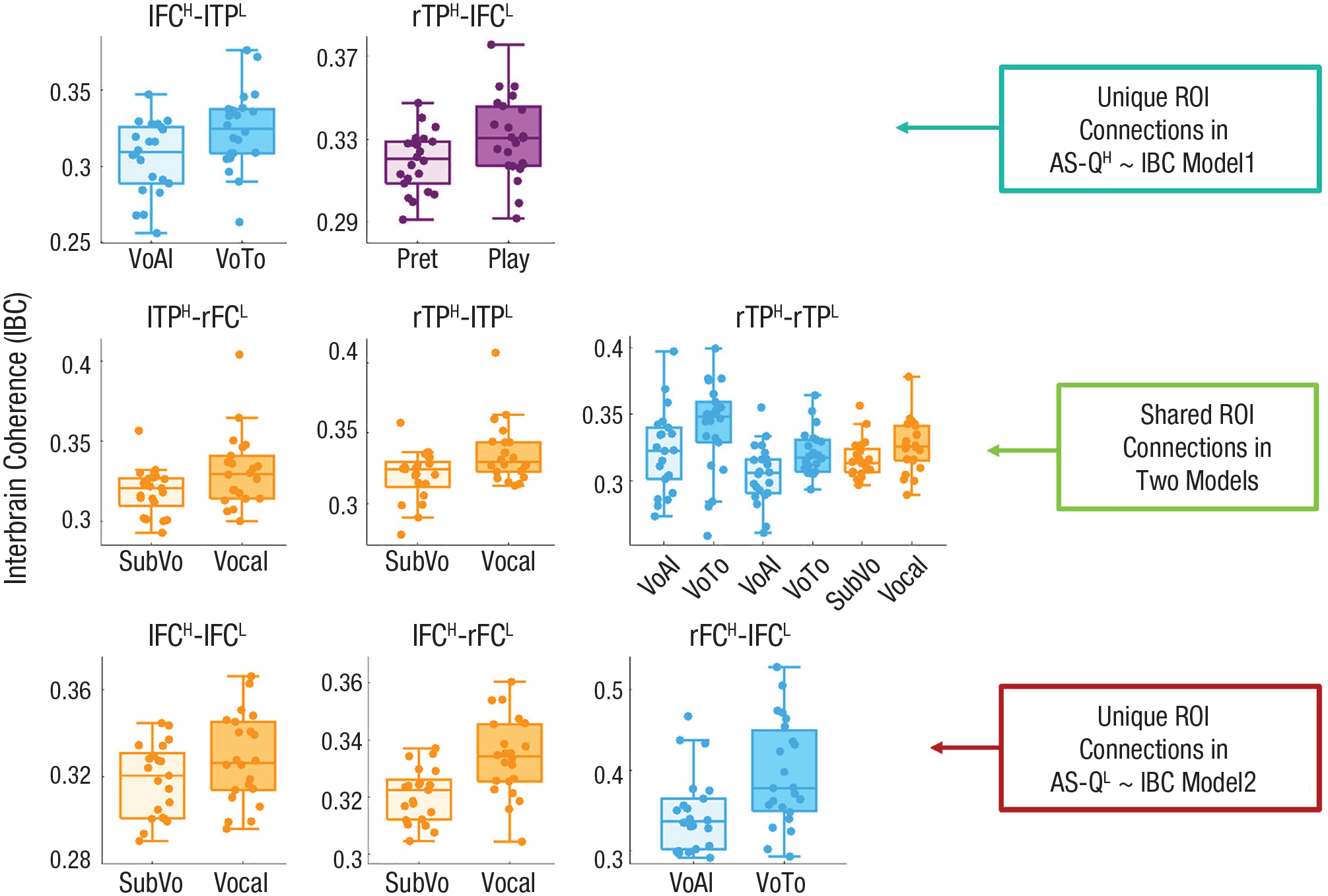
IBC between ROIs that contributed to the two regression models. The interbrain connections between ROIH-ROIL stand for the connection between the ROI in children with higher AS-Q scores (i.e., AS-QH) in each dyad and the ROI in children with lower AS-Q scores (AS-QL) in each dyad. The four ROIs were the lFC, rFC, lTP, and rTP. The conditions were watching video alone (VoAl) and together (VoTo) in Experiment 1 (cyan), self-paced subvocalized (SubVo) and joint and vocalized (Vocal) book reading in Experiment 2 (orange), and pretend Jenga play (Pret) and Jenga play for real (Play) in Experiment 3 (purple). The three rows represent the unique and shared ROI connections in the two models (i.e., AS-QH ~ IBC Model 1 and AS-QL ~ IBC Model 2). IBC = interbrain coherence; ROI = region of interest; lFC = left frontal cortex; rFC = right frontal cortex; lTP = left temporoparietal area; rTP = right temporoparietal area.
Effects of autistic traits on intrabrain coherence
We further attended to how autistic traits of children affect their own brain connections during peer interactions. Similar to the calculation of IBC, the IntraBC between four ROIs for individuals (n = 46) was calculated in each experiment. During video watching (Fig. 7), the connection between the lFC and rTP across individuals showed stronger IntraBC when children watched the video together than alone. During book reading, the following connections across individuals showed stronger IntraBC during joint reading compared with the self-paced subvocalization session: lFC-rFC, lFC-lTP, rFC-lTP, rFC-rTP, and rTP-rTP (Fig. 7). During Jenga playing, the lTP-rTP and rTP-rTP connections showed greater IntraBC when children played Jenga for the real-play compared with pretend-play session. The ROI connections that showed significant increases in IntraBC for interaction in the three experiments are summarized in Table 3. Similar to the regression model of IBC for the AS-Q scores above, regression analyses of IntraBC were done for individuals’ AS-Q scores. The results showed that the combination of rFC-lTP and rFC-rTP connections during book reading and the lTP-rTP during Jenga playing predicted the AS-Q scores in the subcategories of attention to detail and mind reading. The combination of rFC-lTP and rTP-rTP connections during book reading and the lTP-rTP connection during Jenga playing predicted the AS-Q scores in the subcategory of social skills. Overall, these results suggest that the autistic traits of children modulate their intrabrain connections, which may support the strength of their interbrain connections with their peers.

Regression models of IntraBC for child AS-Q scores. The lines connecting the ROIs and circles around the ROIs indicate intrabrain connections that showed a significant increase in IntraBC because of the interaction in each experiment after multiple comparisons corrections. The IntraBC entered in the regression models was the difference in IntraBC between the two conditions in each experiment. IntraBC = intrabrain coherence; AS-Q = autism-spectrum quotient; ROI = region of interest; lFC = left frontal cortex; rFC = right frontal cortex; lTP = left temporoparietal area; rTP = right temporoparietal area.
Summary of ROI Connections That Showed Significant IntraBC Results in Three Experiments

Note: From left to right, the columns show the two conditions in each experiment, the ROI connections that showed significant differences in IntraBC across individuals, the frequency ranges of interest that revealed the differences in IntraBC between the two conditions, and the adjusted p values for multiple comparisons corrections. ROI = region of interest; IntraBC = intrabrain coherence; VoTo = video together; VoAl = video alone; Vocal = vocalize; SubVo = subvocalize; Play = real play; Pret = pretend play; lFC = left frontal cortex; rFC = right frontal cortex; lTP = left temporoparietal area; rTP = right temporoparietal area.
Discussion
From copresence without interaction (video watching) to implicit joint activity (book reading) and to explicit turn-taking activity (Jenga play), the current study examined IBC among school-aged children with varying autistic traits.
Social synchronization between dyads of children
We found increased IBC in the frontotemporoparietal network (Table 2) in the low-frequency range and increased IntraBC between lFC and rTP (Table 3) among children watching video alone to together. Both the FC and TP are parts of the mentalization network; the ventromedial FC is involved in enduring trait inferences and characteristics, and the TP is responsible for the judgment of temporary beliefs and intentions (Van Overwalle & Vandekerckhove, 2013). Azhari et al. (2020) reported stronger IBC in the lFC between couples when they listened to infant/adult laughter together than alone, although with no differences in the two conditions when they listened to infant/adult cries, consistent with our results. Results in these two studies suggest that the mere presence effect on IBC may depend on the environment and experimental design.
From self-paced to joint book reading, we found increased IBC (lFCH-lFCL, lFCH-rFCL, and lTPH-rFCL) in the low- to middle-frequency ranges and increased IntraBC (lFC-rFC, lFC-lTP, and rFC-lTP) in individuals. These results suggest that increased IBC between peers was supported by stronger IntraBC in individuals to align their behaviors with their partners, including synchronized respiration in joint reading. Further, the increases in IBC in the lTPH-rFCL were negatively correlated with child AS-Q scores. These results suggest that when either child is more autistic, the increase in dyadic IBC during joint reading would be smaller. The involvement of the bilateral FC of ChildL and the lFC and lTP of ChildH in each dyad suggests their different roles (Sänger et al., 2013) and contributions to the aligned behaviors through the processes of continuous attention allocation and metacognition (Gvirts & Perlmutter, 2020). Previous studies have reported increased IBC in the lFC between TD individuals in joint versus solo singing (Osaka et al., 2015) and in coordinated versus uncoordinated musical plays (Lindenberger et al., 2009), consistent with our results. The rhythmic behaviors and synchronized motor activities in these tasks are influenced by a shared goal, individuals’ knowledge about the materials, and continuous allocation of attention to facilitate coordination, which contributed to the increase in IBC compared with when no coordination was available.
From pretend to real Jenga play, we found stronger IBC (lFCH-rTPL, lTPH-rTPL, and rTPH-lFCL) in the middle to high frequencies and stronger IntraBC within the bilateral TP. Explicit interaction during real Jenga play requires the perception and continuous evaluation of self and others’ mental states, behaviors, and intentions and cooperation to achieve the shared goal, whereas pretend play involves conceiving and engaging in a nonliteral situation, which is challenging for autistic individuals. The greater involvement of the rTP in interbrain and intrabrain connections in real Jenga play is consistent with the functional role of the rTP (Yang et al., 2015), which has been related to attention monitoring and supporting social cooperation. Previous studies have also found greater IBC when individuals cooperated to perform a task compared with when they performed a task separately (Fishburn et al., 2018; Wang et al., 2020) and compared with when individuals competed in a task in the FC (Kruppa et al., 2021; Reindl et al., 2018). Cooperative interactions increased IBC in a team compared with separate activities that entailed no interaction or motor synchrony and competitions that entailed interactions among all participants and mental states that are aligned with individuals but not shared interests. Overall, results in the current study and in previous studies suggest that a tangible/shared goal synchronizes two individuals in the frontotemporoparietal network.
Autistic traits modulate IBC during peer interactions
Results from our regression models of IBC that predicted the higher and lower child AS-Q scores in each dyad (i.e., AS-QH ~ IBC and AS-QL ~ IBC) show shared and unique interbrain connections during peer interaction (Fig. 6). The shared components between the two models consisted of IBC involving the rTP of both ChildL and ChildH with other ROIs during video watching and book reading. In addition to IBC, the rTP in individuals also showed stronger IntraBC in the middle frequencies during book reading and Jenga playing that did not contribute to the prediction of autistic traits. These results suggest that stronger intrabrain and interbrain connections in the bilateral TP of individuals may support attention control and behavior monitoring such as aligned respiration during peer interaction and that these interbrain connections were not distinctive among children with varying autistic traits.
The unique interbrain connections within the AS-QH ~ IBC model consisted of the lFCH-lTPL during video watching and rTPH-lFCL during Jenga playing. The unique interbrain connections within the AS-QL ~ IBC model consisted of the connections between the bilateral FC during video watching and book reading, suggesting the unique contribution of the less autistic child in each dyad during peer interaction. Consistent with our results, cortical activity and IBC in the FC have also been reported to support the execution and observation of nonverbal communicative gestures and emotion regulations (Reindl et al., 2018) and eye gaze (Leong et al., 2017), which are impaired in individuals with ASD. Between autistic children and their parents, IBC in the FC during a cooperative key-press task was modulated by children’s autistic symptoms (Wang et al., 2020). The unique interbrain connections involving the bilateral FC of ChildL (less autistic) during reading could also be related to their comprehension of the content, which drives aligned reading rhythms in the joint reading. Future research could test this hypothesis and examine how varying levels of comprehension affect IBC among children with different autistic traits.
Implications and future directions
The current study revealed findings about how IBC in peer-to-peer interactions in school-aged children was modulated by autistic traits. These findings demonstrate how autistic traits are tied not only to the brain of the individual but also how the individual interacts with others. Because social interaction is an important feature of autism, the current study provides for the first time the biological underpinnings of differences in social interaction associated with autism that are beyond the individuals themselves. On the basis of our findings, future research should consider whether peer-to-peer IBC is plastic in relation to support programs, and if so, what goals and strategies could modify IBC patterns. Future research should also consider social interaction beyond dyads because multiperson interactions are not only ecologically realistic (Dikker et al., 2017) but also introduce a whole host of variables that are theoretically interesting. As the field moves forward to better understand autism both as a diagnostic category and a representation of neurodiversity (Pellicano & den Houting, 2022), future research could investigate individuals not only on the basis of autistic traits as a continuous variable but also autism as a categorical variable. Future research could also investigate how studies of hyperscanning may be applied to developing interventions for individuals with different baseline communication characteristics (Ingvalson et al., 2013; Wong et al., 2017).
Supplemental Material
sj-docx-1-pss-10.1177_09567976241237699 – Supplemental material for Autistic Traits Modulate Social Synchronizations Between School-Aged Children: Insights From Three fNIRS Hyperscanning Experiments
Supplemental material, sj-docx-1-pss-10.1177_09567976241237699 for Autistic Traits Modulate Social Synchronizations Between School-Aged Children: Insights From Three fNIRS Hyperscanning Experiments by Xin Zhou, Xuancu Hong and Patrick C. M. Wong in Psychological Science
Footnotes
Acknowledgements
We thank all the parents and children for participating in this research.
Transparency
Action Editor: Karen Rodrigue
Editor: Patricia J. Bauer
Author Contributions
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.