
Abstract
A multiproxy sedimentary record from Lake Igaliku in southern Greenland documents 1450 years of human impacts on the landscape. Diatoms, scaled chrysophytes, and C and N geochemistry show perturbations consistent with recent agricultural activities (post-
Introduction
Arctic ecosystems are subject to an increasing number of stressors. The effects of climate change are becoming increasingly evident in the circumpolar North in the form of accelerating glacier melt (Zwally et al., 2002), reduced Arctic Ocean summer sea ice extent (Serreze et al., 2009), and marked biological shifts in both aquatic and terrestrial ecosystems (Post et al., 2009; Smol et al., 2005). Operating at the same time is a range of anthropogenic stressors (e.g. atmospheric deposition of nutrients, persistant organic pollutants (POPs), mercury) whose ecological implications are still largely unknown (Arctic Monitoring and Assessment Programme (AMAP), 2009). Adaptation to current and future climate change requires understanding not only of the range of natural climate variability and scenarios for the future, but also the response of ecosystems and civilizations to multiple stressors.
The south coastal region of Greenland is situated within a key geographic area for understanding North Atlantic climate variability, and occupies an important place in human history. The Norse settled southern Greenland ~
The establishment of the Norse farms in Greenland is thought to been aided by the favourable climatic conditions of the Medieval period. The cause of the collapse of their colony however, has been the source of much speculation (McGovern, 2000). One reason is thought to be largely climate-driven: As the climate cooled with the beginning of the LIA (
The paleoenvironment of Norse sites in the North Atlantic has been studied extensively using pollen (e.g. Edwards et al., 2008; Fredskild, 1988; Gauthier et al., 2010; Schofield et al., 2008, 2010) and sedimentological analyses (Sandgren and Fredskild, 1991). While a few studies have investigated the impact of Norse activities via changes in-lake ecology using the remains of chironomids elsewhere in the North Atlantic (e.g. Faroes: Gathorne-Hardy et al., 2007; Iceland: Lawson et al., 2007), to date, no studies have used either sedimentary diatoms or carbon and nitrogen stable isotopes in lake sediments to investigate the environmental impacts of the Norse. This is despite the wide use of these paleolimnological proxies in other paleoenvironmental applications (Smol and Stoermer, 2010). Diatoms, in particular, have been successfully used to trace the nature of human impacts on the landscape as early as the Neolithic and Bronze Age (Bradshaw et al., 2005; Fritz, 1989) and for disentangling the relative impacts of early human settlement patterns in boreal and temperate regions (Anderson et al., 1995; Ekdahl et al., 2004). In the Arctic, diatoms are commonly used to trace environmental responses to Holocene climate variability as well as recent warming (Smol et al., 2005; Wolfe, 2002). They have also been used to investigate other prehistoric and historic cultural impacts in the Arctic (i.e. from whaling cultures (Douglas et al., 2004) and sewage treatment (Douglas and Smol, 2000)) but have not yet been used to investigate agricultural impacts.
Despite the fact that Greenland Norse colonies are commonly thought to have been influenced by late-Holocene climate variability, to date not a single continuous, high-resolution, multiproxy paleoenvironmental record has been generated within immediate proximity of a Norse site in Greenland. Therefore, the patterns of late-Holocene natural variability are unknown at these sites, as is the extent to which Norse farming caused the purported environmental destruction often claimed. Here we present a high-resolution multiproxy biological and geochemical record from Lake Igaliku that explicitly tests the magnitude of Norse landscape impacts relative to the natural variability of the last 1450 years, and compares the period of Norse tenure (
Methods
Study area
Lake Igaliku (61°00'N, 45°26'W, 30 m a.s.l.) occupies a low valley between the head of Igalikup Kangerlua (Igaliku Fjord) and Tunulliarfik fjord (Erik’s Fjord) in southern Greenland (Figure 1). The catchment area of the lake (3.1 km2) is of relatively low relief with a modern farm and pastures close to the lake and bedrock hills reaching 300 m a.s.l. to the south. The bedrock of the region is composed of Ketilidian Proterozoic crystalline granite overlain by the volcano-sedimentary rocks of the Eriksfjord formation, which outcrops in the hills to the northwest. This region of southern Greenland is classified as low Arctic, where permafrost is discontinuous, and birch and alder copses grow locally in the innermost sheltered fjords. Mean annual temperature is 0.9°C, with July and January means of 10.3°C and −6.8°C, respectively. The lake is 34 ha in area and 26 m deep. It has one outflow, but no major inflow.

Map showing (a) the location of Lake Igaliku on Greenland, (b) the region around Lake Igaliku including roads (dashed lines), buildings (black rectangles) and current (post-1960) hay fields (grey shaded areas) as well as the archaeological site of Gardar, and (c) the bathymetry of the lake and coring location.
From archaeological and written accounts, Igaliku was one of Norse Greenland’s most prosperous farm regions. Originally settled by Erik the Red’s daughter, Freydis, at the beginning of the Norse Landnám, it was given to the bishop in 1126 and became the site of the cathedral as well as the Thing (assembly) for the Eastern Settlement of Greenland (Jones, 1986; Sanmark, 2009). The farms are estimated to have housed between 75 and 100 cattle, in addition to sheep, and thus would have required extensive use of the surrounding land for grazing as well as hay production (Nørlund, 1936). The catchment of Lake Igaliku was never the site of a Norse farm but would have been used for grazing livestock. The modern community consists of 60 permanent inhabitants and was founded in the late 1700s. Agricultural practices resumed in the 1920s, at the same time that the climate of southern Greenland reached its recent maximum (Box et al., 2009). Current sheep farming in the catchment is limited to one farm, established in the early 1960s, which has a barn for wintering sheep and summer hay production on a 30 ha field. A small ditch currently drains effluent from the barn into the nearby lake. The farm currently deploys 750–900 kg N fertilizer per year within the lake catchment to boost yields for winter fodder (Mikki Egede, personal communication, 2011).
Pollen and non-pollen palynomorph (NPP) studies from Lake Igaliku sediments reflect agricultural impacts on the landscape during Norse occupation. A decline in arboreal birch pollen c.
Cores and chronology
A suite of sediment cores was retrieved using piston and gravity cores from the deepest part of Lake Igaliku in June 2007 (see Figure 1). The 4 m composite sequence captures the entire Holocene (c. 10,000 years), including the marine to lacustrine transition c. 8500 yr BP; however, only the upper core is discussed here (the last ~1450 years). The chronology for the core is based on 13 birch macrofossil 14C dates (twigs and leaves), a 137Cs peak (
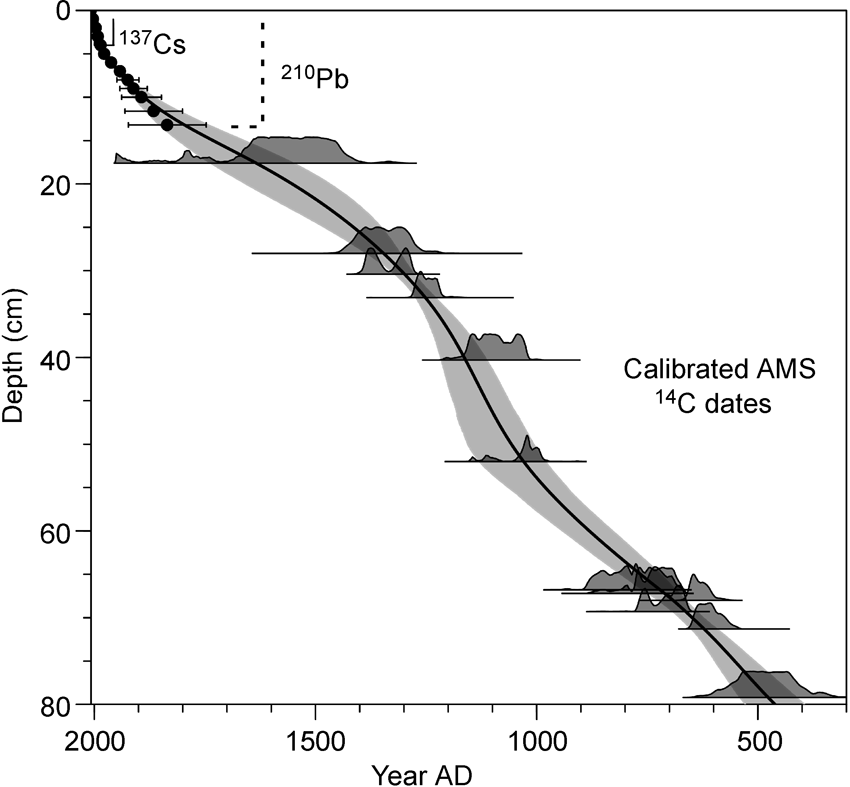
Age–depth model showing the dates, probabilities of radiocarbon dates and the 95% confidence interval for the age–depth model (grey band).
Diatoms and chrysophytes
Diatoms and chrysophytes were prepared from wet sediments using a standard protocol for large sample numbers (Renberg, 1990). 5–10 ml 30% hydrogen peroxide was added to the ~0.3 g samples and heated until the reaction subsided, followed by the addition of several milliliters of 10% HCl. Samples were then rinsed with distilled water and a known quantity of Divinyl Benzene (DVB) microspheres was added to the slurries (Battarbee and Kneen, 1982), which were then dried on coverslips and mounted using Naphrax mounting medium. Using transmitted light microscopy under oil immersion at least 350 diatom valves were identified and enumerated from the slides as well as chrysophyte cysts and scales. Identification of diatoms was aided by reference literature from Greenland (Foged, 1953, 1972, 1977) and Labrador (Fallu et al., 2000).
Stable Isotopes and geochemistry
Core samples for stable isotope and geochemistry measurements were dried and ground to fine powder prior to analysis. The total carbon and total nitrogen contents were measured by total combustion in a Vario Max CNS elemental analyser (Elementar, Germany). Mass loss of the samples was negligible after combustion at 950°C, and thus the total carbon measured is interpreted to be organic. The δ13C and δ15N isotopic analyses were performed in triplicate on an elemental analyser NA 1500 NCS (Carlo Erba Instruments, Milan, Italy) coupled in continuous flow mode to a stable isotope ratio mass spectrometer VG Isochrom (Micromass, Manchester, UK). The results are presented in standard delta notation. Standard errors for δ13C and δ15N measurements are < 0.2‰ and < 0.5‰, respectively. δ13C measurements since
Numerical analysis
Principle components analysis (PCA) was performed on a covariance matrix of diatom taxa with relative frequencies >1%. PCA was chosen because the species assemblages form a short primary gradient established by detrended correspondence analysis (DCA; 1.67 SD).
A second PCA was performed on a correlation matrix of selected diatom taxa from the initial PCA (i.e. those with the highest scores: Cyclotella stelligera, Fragilaria tenera, Cyclotella rossii complex), chrysophyte scales, and geochemical data (C, N, C/N, δ13C, δ15N). These proxies were analysed together in order to generate an overall ecological response to landscape disturbance and natural variability over the last 1450 years.
Results
Diatoms and chrysophytes
Over 140 species of diatoms from 25 genera were identified from the sediments of Lake Igaliku. Scanning electron microscope images of several of the key taxa are shown in Figure 3. The flora is typical of dimictic, circumneutral, oligotrophic West Greenlandic lakes (Perren, 2007). Cyclotella stelligera (=Discostella stelligera) and the complex belonging to variants of Cyclotella rossi, C. comensis, C. tripartita, and C. ocellata (henceforth called Cyclotella rossii complex) dominate the planktonic component and are identical to those found along the west coast of Greenland (Perren et al., 2009) as well as those described from northeast Greenland (Cyclotella sp. A: Cremer and Wagner, 2004). Brachysira vitrea, small Achnanthes and Achnanthidium spp., and Fragilaria sensu lato (=Staurosira, Staurosirella, Pseudostaurosira, Fragilariaforma spp.) are the dominant benthic taxa. Lesser quantities of representatives of the genera Cymbella, Eunotia, Navicula, and Nitzschia are also present.

Scanning electron micrographs of key diatom and chrysophyte taxa from Lake Igaliku sediments. (a) Inner valve view of Cyclotella bodanica (=Puncticulata bodanica) showing central and marginal fultoportulae and two rimoportulae; (b) Cyclotella stelligera (=Discostella stelligera) showing the stellate arrangement of alveoli in the colliculate central area of the valve; (c) chrysophyte scale belonging to Mallomonas crassisquama; external (d) and internal (e) views of Fragilaria tenera showing the apical pore field and single rimportula, respectively.
The stratigraphy of dominant diatom taxa is shown in Figure 4. The diatom assemblages are remarkably stable over the last 1450 years. The most notable exception is in the last 30 years, when Cyclotella stelligera and Fragilaria tenera increase dramatically (to maxima of 60% and 37%, respectively) at the expense of the Cyclotella rossii complex. Chrysophyte scales, belonging principally to the genera Synura and Mallomonas, also increase to over 109 scales/g sediment at the surface against a steady background concentration of between 0–103 scales/g sediment.

Stratigraphic diagram showing the relative frequency of important diatom taxa, the concentration of scaled chrysophytes, the first 2 PCA axes and the Arctic temperature anomaly reconstruction from Kaufman et al. (2009). The periods of Norse Landnám and 20th century agriculture are shown.
Recent changes (post-
The PCA of diatom results show two major features: a major shift in lake ecology ~
Geochemistry
Stable carbon and nitrogen isotopes and geochemical stratigraphies are shown alongside the principal diatom and chrysophyte stratigraphies in Figure 5. Nitrogen content values oscillate between 0.4% and 0.7% dry mass with the lowest values between

Stratigraphic diagram showing the proxies used in the synthesis PCA: chrysophyte scales, key diatom taxa, and geochemistry parameters. The periods of Norse settlement and modern agriculture are shown.
The enrichment of 15N in last 30 years in Igaliku is the inverse of observed recent trends in δ15N. In Greenland Ice Sheet snow and in Arctic lake sediments, δ15N values decline since
Synthesis of multiple proxies
The PCA integrating diatoms, chrysophytes, and N and C geochemistry demonstrates the coherence of the individual proxies with each other (Figure 6a). The first and second axes explain, respectively, 36% and 20% of the variance of the data set. The first axis is strongly associated with N, Cyclotella stelligera, the concentration of scaled chrysophytes, Fragilaria tenera, and δ15N. The diatom complex of Cyclotella rossii dominates the negative end of the first axis. The second axis is dominated by the δ13C values on the positive side, and by C/N ratio on the negative end.

Synthesis PCA results. (a) PCA biplot showing the scores of the proxies used in the analysis (arrows); (b) PCA biplot of the downcore sample scores from the analysis (circles).
The PCA biplot of samples in Figure 6b demonstrates the degree to which recent changes in Lake Igaliku are unprecedented. Norse samples cluster within the range of natural background values (
Discussion
The biogeochemical response of Lake Igaliku documents subtle environmental changes over the last 1450 years, until c.
Scant environmental impact from Norse farming
At Igaliku, the Norse period is registered in the sediments as slightly higher relative frequencies of the diatom taxon Cyclotella stelligera, slightly higher δ15N values (both a proxy for increased production), high C/N ratios, depleted δ13C values, as well as the lowest values of C and N in the last 1450 years. Sedimentological analyses (Massa et al., 2012) demonstrate a higher flux of minerogenic materials during this time, which is consistent with the C and N values shown here and with a possible soil erosion source. The peak of this signal is constrained to the period c.
It is an unexpected result that nearly 500 years of Norse agricultural activities are barely registered in the ecology of adjacent lake sediments. Elsewhere in the Northern Hemisphere, diatoms document nutrient enrichment in response to early agriculture by Anderson et al. (1995); Bradshaw et al. (2005); Ekdahl et al. (2004); Fritz (1989). In southern Greenland, widespread evidence from a number of nearby peat, soil, and lake sediment paleoenvironmental studies documents abrupt vegetation change (Fredskild, 1988; Schofield et al., 2008, 2010), as well as erosion and soil degradation (Jakobsen, 1991; Sandgren and Fredskild, 1991) consistent with the timing of the Norse Landnám. Entomological and δ15N isotopic evidence from infields also document extensive manuring and irrigation to maximize hay production (Buckland et al., 2009; Commisso and Nelson, 2008), which would be consistent with large landscape-level disturbance and the potential for soil erosion.
The extent to which the Norse caused the systematic regional degradation of the landscape, can be questioned. Our findings echo those from the Sandøy, on the Faroe Islands, which show limited paleoenvironmental evidence of Norse landscape degradation (Gathorne-Hardy et al., 2007). Although there is little doubt from coprophilous fungal and sedimentological evidence (Gauthier et al., 2010; Massa et al., 2011) that the catchment and immediate shore of Lake Igaliku were used for grazing and therefore subject to some degree of erosion; the overall ecological impact of the Norse settlement was probably very localized.
The minimal role of climate at Lake Igaliku
Paleoclimatic archives from the circumpolar North suggest that the last 2000 years encapsulates both the warmest and coldest climates of the late Holocene (Bradley, 2000). Although warmth during the Medieval time (~
The LIA is expressed along the southwestern Greenland margin and the inland ice as a period of extreme cold, dustiness, and drought (Andersen et al., 2006; Dahl-Jensen et al., 1998; Kaplan et al., 2002), with local glaciers achieving their late-Holocene maxima (Weidick et al., 2004). The LIA at Igaliku is marked by greater proportions of large planktonic Cyclotella species (e.g. C. rossii complex) and by Fragilaria virescens v. exigua. These shifts are consistent with a temperature-driven phytoplankton size-fraction response found by Winder et al. (2009), where warm conditions favour small phytoplankton (i.e. Cyclotella stelligera) and vice versa. The geochemical data, on the other hand, show highly variable values with no clear trend during the LIA.
Medieval versus modern farming impacts: An historical analogue?
A good historical analogue for the impact of Norse sheep farming exists: From 1920 to 1976, sheep farming in southern Greenland used methods not entirely dissimilar from the Norse. Sheep were left to forage in winter and minimal amounts of fodder were produced. The period prior to 1976 is unremarkable in the biogeochemical proxies at Lake Igaliku despite large sheep populations (upwards of 50,000 in S Greenland) on the land. However, beginning in 1976, the method of farming shifted towards fodder production and higher yields at slaughter which introduced fertilizers (250–300 kg/ha per yr) and effluent from winter sheep stables into the local landscape and lake (Figure 7: agricultural phase II; Greenland Agriculture Advisory Board, 2009). After 1976, sediments from Igaliku show a rise in planktonic diatoms (Cyclotella stelligera, Fragilaria tenera), as well as chrysophyte scales, δ15N, and N, reflecting increased nutrient additions and the beginning of industrialized agriculture. These effects supercede all changes associated with the Norse Landnám.

Stratigraphic diagram showing variations in δ15N, Fragilaria tenera, Cyclotella stelligera, and chrysophyte scales, the synthesis PCA axes 1 and 2, the mean annual temperature anomaly from the Narsarsuaq meteorological station, June, July and August (JJA) temperature from Narsarsuaq, and the total population of sheep in Greenland for the last 150 years. Grey bands mark the recent documented sheep population crises.
The Norse-modern farming analogue also demonstrates the sensitivity of Arctic agriculture. In good years, between 1920 and 1976, the sheep yields in southern Greenland were high. In bad years, such as the cold, snowy spring of 1967, nearly 60% of the sheep population perished from starvation (Greenland Agriculture Advisory Board, 2009), with a similar situation again in 1971 and 1976 (Figure 7; Austrheim et al., 2008). The mean annual temperature for these years is 1–2°C below the 1873–2005 average (Figure 7; Carstensen and Jørgensen, 2010), yet had catastrophic consequences for sheep farming in Greenland. Although the Norse sheep populations were likely far smaller than those of today, two or three consecutive years with late snowy springs, as are known to have occurred throughout the last several millennia (Barlow et al., 1997; Ogilvie, 1991), could have easily crippled subsistence farming in Greenland, putting added reliance on marine resources and decreasing the resilience of the Norse in Greenland.
The last 30 years at Igaliku: Multiple stressors (double trouble?)
Determining the relative influence of climatic forcing versus land use changes on aquatic ecosystems is a difficult task, especially when the two phases of human settlement on the land coincide with the warmest periods of the last 2000 years. This is especially difficult in the Arctic, where enhanced warming will likely alter nutrient cycling dynamics. Recent climate warming since ~1960 is widely documented from a number of sites in the Arctic (Arctic Climate Impact Assessment (ACIA), 2005) and Arctic temperature trends exceed global averages two- to threefold over the last century (Trenberth et al., 2007). Despite this, warming is a relatively recent phenomenon in southern Greenland (Box et al., 2009). Meteorological records from the nearest weather station at Narsarsuaq, Greenland (1873–present; Figure 7), show that only the most recent years, since 2005, attain the mean annual temperature maxima of the 1920–1950s, which were the warmest period of the instrumental record (Box et al., 2009; Carstensen and Jorgensen, 2010). Summer temperatures (JJA), however, show steady warming over the instrumental period with a larger increase since the 1980s (Figure 7). The fact that the well-dated upper sequence at Igaliku does not show a shift in the decades following the 1920s, but at
Conclusions
The microfossil and geochemical proxy records from the last 1450 years at Lake Igaliku do not support the hypothesis that the Norse strongly impacted their environment. The five centuries of Norse farming in southern Greenland are registered in the sedimentary biological and chemical proxies as subtle deviations from long-term means, as opposed to the marked shifts documented since the advent of mechanized farming in ~1976. Similarly, the natural environmental variability of the last 1450 years is eclipsed by the recent changes seen in Lake Igaliku.
Despite subtle historical impacts, recent changes in Lake Igaliku, evidenced by marked shifts in diatoms, scaled chrysophytes, and δ15N, suggest that the lake is sensitive to changing land use. Recent farming activities have outpaced the geochemical and biological resilience of the lake and have likely heightened its sensitivity to future stressors. Under climate change scenarios, South Greenland is expected to warm 3–6°C over the next 100 years (ACIA, 2005). With additional pressure from the agricultural sector, these stressors will likely result in unanticipated new ecological states in the future. These findings confirm the sensitivity of Arctic lakes to external anthropogenic forcing and contribute to the understanding of the relative differences between the historic and modern impacts of agriculture in the North and their sensitivity to future stressors.
Footnotes
Acknowledgements
The authors are grateful to M Campy, H Grisey, and B Vannière for technical help in Greenland and to the farmer Mikki Egede at Igaliku and Aqqalooraq Frederiksen of the Greenland Agricultural Advisory Service in Qaqortoq, for providing information about current agricultural practices. We also thank AP Wolfe and two anonymous reviewers for providing comments on the manuscript.
Funding
Financial support for this study was provided by the University of Franche-Comté, the University of Burgundy and the French Polar Institute (IPEV).