
Abstract
Over the last 200 years Australia has suffered the greatest rate of mammal species extinction of any continent. This demands extensive biodiversity research, but unfortunately has been hampered by poor documentation of Australia’s native species at the time of European colonization. Late-Holocene fossil mammal assemblages preserved in caves, rockshelters and surface lag deposits from deflated sand dunes can provide a more complete understanding of pre-European ecological conditions than can be developed from our knowledge of present biodiversity. In South Australia, few regions have experienced greater landscape modification and biodiversity loss than Yorke Peninsula. We investigate the composition, richness, evenness and age of two owl accumulations from southeastern and southwestern Yorke Peninsula and contrast them with a surface lag deposit assemblage probably accumulated by humans. We then examine the pre-European biogeography of the fauna recovered. The three assemblages have similar species richness, but differ dramatically in composition and evenness. The biases imposed by differing accumulation agents can explain compositional differences between owl and human assemblages, but not the differences between the respective owl accumulations. We argue that key substrate differences – one area is dominated by sand and the other by calcrete – have favoured distinct vegetation communities that fostered distinctly different mammal assemblages from which raptors accumulated prey. The ecological requirements of the extant mammals appear to be reflected in the fossil assemblages, providing support for the application of uniformitarian principles and confidence in the relevance of late-Holocene fossil assemblages to modern conservation and natural resource management.
Keywords
Introduction
European colonization has had major environmental repercussions that have fundamentally transformed Australia’s species, ecosystems and landscapes, causing widespread declines in biodiversity (e.g. Bickford and Gell, 2005; McDowell and Medlin, 2010). Faced with environmental change, organisms may remain unaffected, or may respond with dispersal, population size or density change, clinal shift, evolution, extirpation or extinction (Barnosky, 2008; Barnosky and Bell, 2003; Hadly, 1996). Therefore, it is of fundamental importance for natural resource management that we understand the impacts of past environmental change as a key to predicting and preparing for future changes in biodiversity. Such predictions rely on a detailed understanding of the impacts of past environmental variation, yet few data sets used in biodiversity assessments span more than one full generation of the targeted organism (Froyd and Willis, 2008; Willis et al., 2005, 2007).
When historical records lack adequate temporal depth, conservation, natural resource and landscape managers can turn to the Holocene palaeoecological record to develop a better understanding of the ecology of extant organisms and their response to past environmental changes (Birks, 1996; Delcourt and Delcourt, 1998; Froyd and Willis, 2008; Jackson and Erwin, 2006; Lyman, 1996; Swetnam et al., 1999). Species composition, diversity, relative abundance, richness and evenness of Holocene fossil assemblages can be compared with those of modern ecosystems to assess the degree of modification or degradation that an ecosystem has experienced (Hadly and Barnosky, 2009).
Holocene fossil deposits usually consist of unstratified surficial raptor assemblages and lag deposits (e.g. dune blowouts). Because of the dynamic nature of such settings, they are typically assumed to be geologically very young. It might even be argued that the age of such accumulations is not relevant because of the high potential for time averaging, a taphonomic process whereby specimens accumulated at different times becoming mixed together. Some researchers (Barnosky and Shabel, 2005; Barnosky et al., 2003; Hadly, 1999) have suggested that because of an increased potential for the inclusion of rare taxa, time averaging may actually be beneficial under these circumstances. However, with no knowledge of the extent of time that has elapsed over the depositional period, this concept is difficult to assess. Further, without radiometric dating, comparison between fossil accumulations erroneously assumed to be of similar age may be invalid.
In Australia to date efforts have been made to determine the pre-European vertebrate fauna for various regions (e.g. Baynes, 1984, 1987; Baynes and Baird, 1992; Baynes and Johnson, 1996; Baynes and McDowell, 2010; Copley et al., 1989; Medlin, 1993; Morton and Baynes, 1985; Start et al., 2012; Wakefield, 1972), but only one study of a late-Holocene mammal assemblage (McDowell, 1997) has investigated paleoecological change within a tight chronological framework.
No region of South Australia has been more impacted by European colonization than Yorke Peninsula (Figure 1). Owing to its gentle topography and relatively predictable climate, Yorke Peninsula has been subjected to extensive vegetation clearance for agriculture, habitat modification, introduction of invasive species, and altered environmental water flows and fire regimes (Department for Environment and Heritage (DEH), 2007). Consequently, little is known of the pre-European mammalian fauna. Recent biological surveys (Neagle, 2008) have provided information about Yorke Peninsula’s current biodiversity, but the potential of the Holocene fossil record remains untapped. This study seeks to test key assumptions frequently made about Holocene fossil deposits, which include their age, the effects of time averaging, their utility for determining species distribution baselines and the validity of uniformitarianism. We use AMS radiocarbon dating to assess the age and extent of time averaging, and analyze composition, relative abundance, richness and evenness of mammal species in two large unstratified owl accumulations (INPA and TRHA, Figure 1) and one surface lag deposit (BALA, Figure 1). The assemblages are compared and contrasted to assess their relevance to natural resource and conservation management and the degree of time averaging they have experienced.
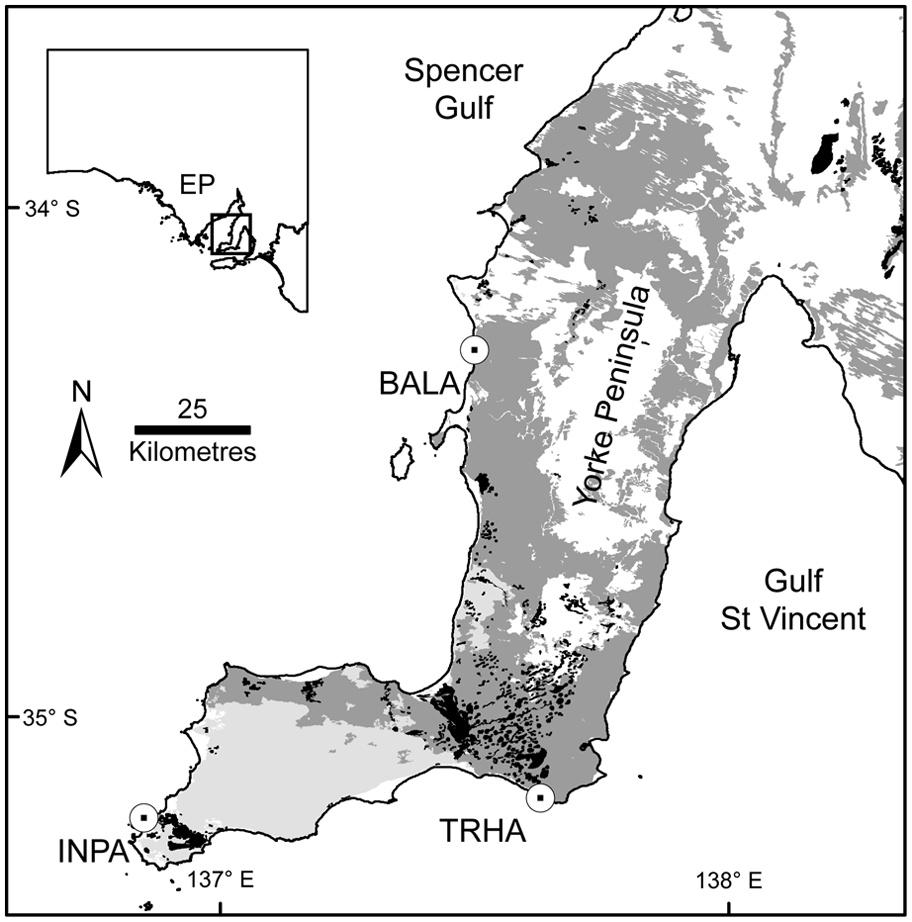
Location of fossil assemblages, pre-European vegetation and geomorphology of Yorke Peninsula, South Australia (after Department of Water, Land and Biodiversity Conservation (DWLBC), 2005). EP: Eyre Peninsula; BALA: Balgowan assemblage; INPA: Innes National Park assemblage; TRHA: Troubridge Hill assemblage; black: ephemeral salt lakes; dark grey: Dryland Tea-tree–Drooping Sheoak low open woodland on calcrete; light grey: Mallee on sand; white: other.
Study area
Yorke Peninsula experiences a Mediterranean climate with mild wet winters and hot dry summers (Neagle, 2008). Southwestern Yorke Peninsula receives a mean annual precipitation of 478.3 mm (average of 48 years of data collected intermittently between 1893 and 2010 from Marion Bay, Innes National Park and Stenhouse Bay weather stations combined; Bureau of Meteorology, 2011), whereas southeastern Yorke Peninsula receives a mean annual precipitation of 411.4 mm (average of 122 years of data collected between 1875 and 2001 from Edithburgh Post Office weather station; Bureau of Meteorology, 2011).
Southern Yorke Peninsula has two distinct surface substrates derived from different sedimentary processes and the underlying Bridgewater Formation (Milnes and Ludbrook, 1986). The two substrates support different vegetation associations (Figure 1; DWLBC, 2005). Mobile calcareous sands derived from coastal dunes dominate the western part of southern Yorke Peninsula where the Innes National Park assemblage (INPA) was found (DWLBC, 2005). Prior to European colonization, this substrate supported Kingscote Mallee–Coastal White Mallee (Eucalyptus rugosa–E. diversifolia) associations with patches of vegetation dominated by White Mallee (E. dumosa–E. phenax) (Croft, 2008). Remnants of this vegetation indicate a shrubby understorey that includes Dryland Tea-tree (Melaleuca lanceolata), Mallee Wreath Wattle (Acacia triquetra), Rock Wattle (A. rupicola) and Wedge-leaved Pomaderris (Pomaderris obcordata). By contrast, the surface geology of the eastern part of southern Yorke Peninsula, where the Troubridge Hill assemblage (TRHA) was found, is dominated by a discontinuous cover of shallow loamy calcareous soil overlying calcrete (DWLBC, 2005). Before European colonization, this region supported Dryland Tea-tree–Drooping Sheoak (M. lanceolata–Allocasuarina verticillata) low open woodland (Croft, 2008) around and within the numerous ephemeral salt lakes and associated gypsum deposits that occur in the area (DWLBC, 2005). The understorey was probably open and grassy with sparse shrubs or understorey trees (Croft, 2008). This vegetation association occurs on the majority of the west coast of Yorke Peninsula, including the area where the Balgowan assemblage (BALA) was found (Figure 1).
Yorke Peninsula was officially settled by European farmers in 1846 (Carmichael and Mudie, 1973). It has been subjected to such extensive clearing and landscape modification, that Aitken (1973: 156) observed that ‘Few regions of South Australia have such a depleted fauna of native mammals as Yorke Peninsula’. He listed only eight non-volant native terrestrial mammal species: one monotreme Tachyglossus aculeatus, and seven marsupials Sminthopsis crassicaudata, Lasiorhinus latifrons, Trichosurus vulpecula, Macropus eugenii, Macropus fuliginosus, Macropus robustus and Cercartetus concinnus, collected from the Peninsula following European settlement, but did not specify localities. Of these M. eugenii have since been lost, though L. latifrons and T. vulpecula may still be present (Robinson et al., 2000a). Only three native non-volant terrestrial mammals (T. aculeatus, M. fuliginosus and C. concinnus) remain extant in Innes National Park (DEH, 2003), although M. eugenii has recently been re-introduced (DEH, 2004). The absence of Rattus fuscipes from the modern Yorke Peninsula fauna is probably due to the presence of introduced Rattus rattus (Brandle, 2008), which can competitively exclude R. fuscipes (Cox et al., 2000).
Methods
AMS radiocarbon dating
Three complete long bones of Perameles each weighing approximately 1 g were selected from both the INPA and TRHA and submitted to the Waikato radiocarbon laboratory for AMS dating. Samples were processed using standard ultrafiltration AMS radiocarbon techniques (see Beaumont et al., 2010). Fossils from BALA were not radiocarbon dated because all specimens are highly weathered and considered unlikely to contain datable collagen. Ages were calibrated to the SHCal04 Southern Hemisphere calibration curve (McCormac et al., 2004) using OxCal v4.1.7 (Bronk Ramsey et al., 2010).
Collection and analysis
Fossils were collected from unstratified, calcareous sediments found in shallow coastal caves and a deflated sand dune. The INPA (Figure 1) was collected by C Kemper, P Rismiller, L Jansen and M Ayre in March 1988. The initial sample of the TRHA (Figure 1) was collected by T Worthy and A Camens in November 2008. A second sample was collected from this site by one of the authors (MCM) in April 2011. The BALA (Figure 1) was collected from a lag deposit on a deflated sand dune by M Fuller over a number of visits between 1986 and 1988. The BALA collection is held in the South Australian Museum (SAM) Palaeontology collection and the INPA and TRHA are held in the SAM Mammal collection.
Fossils from INPA and TRHA were initially handpicked from the cave floor. When the absence of stratigraphy became evident the top 10 cm of sediment from a small area of the floor was excavated and gently sieved through 1.5 mm mesh. Concentrated samples were then wet sieved and sorted to separate diagnostic bones which were picked out with forceps and stored in labeled containers. Fossils from BALA were handpicked from the dune surface. Diagnostic bones (whole and part skulls, maxillae, dentaries and/or teeth) were identified using published descriptions and comparative material held by the SAM. All specimens were identified to the lowest taxonomic level possible (usually species). The minimum number of individuals (MNI) was determined by counting the number of the most common diagnostic element of each species in each assemblage. To facilitate comparisons between samples, MNI was converted to relative abundance (Ri%), an expression of the MNI of a given species as a proportion of the total MNI for that collection. Richness (S) and Buzas-Gibson Evenness (E) indices were calculated to compare structural differences between each fossil assemblage. Prey mass was used to characterize accumulation agents. Mean adult body masses for all species were derived from van Dyck and Strahan (2008).
Results
AMS radiocarbon dating
AMS radiocarbon ages obtained from INPA and TRHA (Table 1) indicate that both assemblages were deposited within the last millennium. The INPA had a maximum age span of between 228 and 142 cal. BP, and appears to have accumulated very quickly. By contrast, TRHA had a maximum age span of between 921 and 661 cal. BP years. Because it accumulated more slowly, the TRHA may have experienced greater time averaging and may therefore have incorporated a greater number of rare species. However, as over half of the specimens recovered were attributable to a single species (Table 2), time averaging may have had minimal affect. The absence of the introduced house mouse, Mus musculus, from INPA and the occurrence of only a single specimen in TRHA are consistent with the radiocarbon ages.
AMS radiocarbon ages of bones from Innes National Park assemblage (INPA) and Troubridge Hill assemblage (TRHA), Yorke Peninsula, South Australia.

The late-Holocene non-volant mammal fauna of Yorke Peninsula, South Australia: minimum numbers of individuals (MNI) and relative abundances (Ri%) of species recorded from three sites, Balgowan assemblage (BALA), Innes National Park assemblage (INPA) and Troubridge Hill assemblage (TRHA).
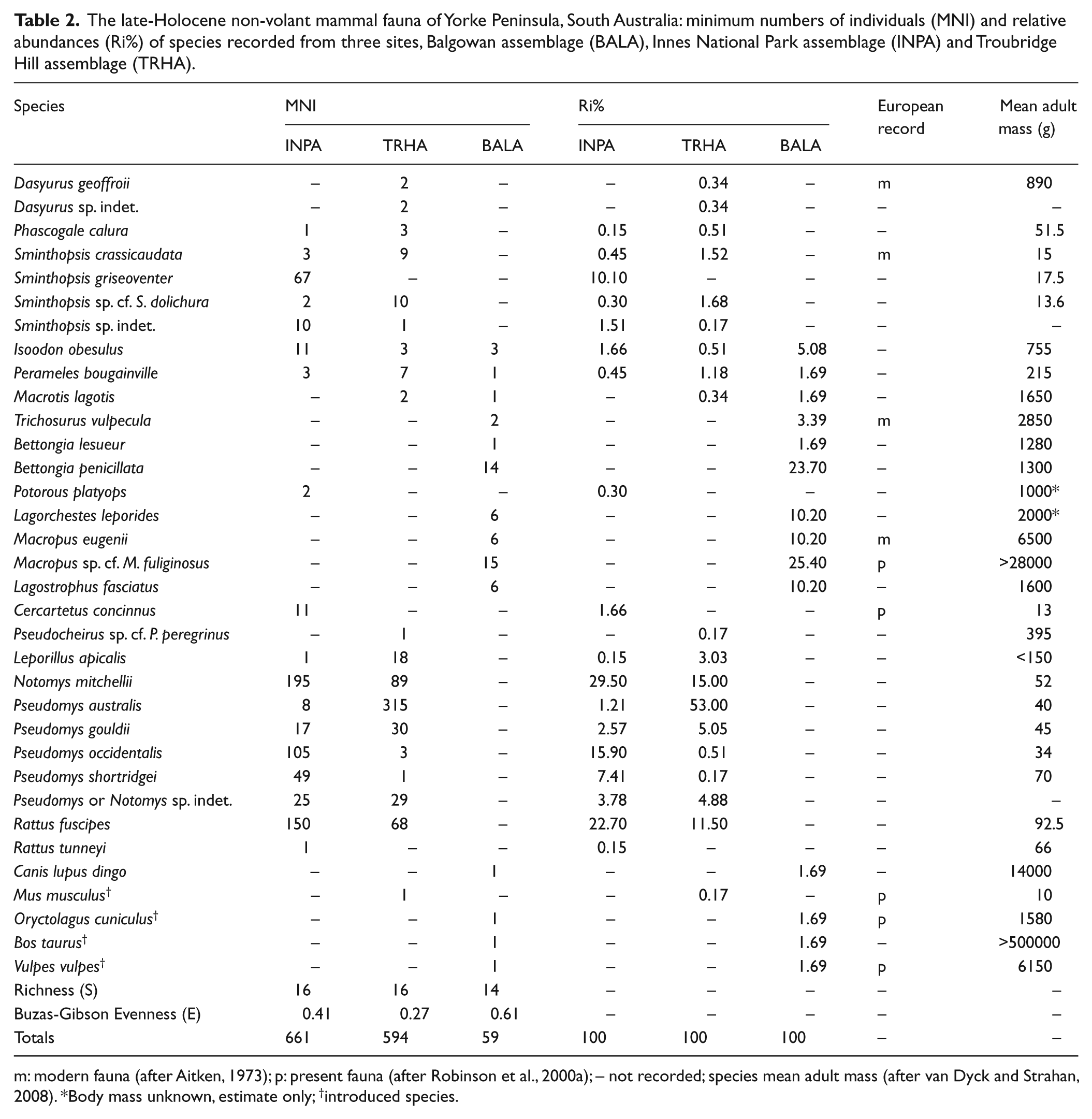
m: modern fauna (after Aitken, 1973); p: present fauna (after Robinson et al., 2000a); – not recorded; species mean adult mass (after van Dyck and Strahan, 2008). *Body mass unknown, estimate only; †introduced species.
Faunal analysis
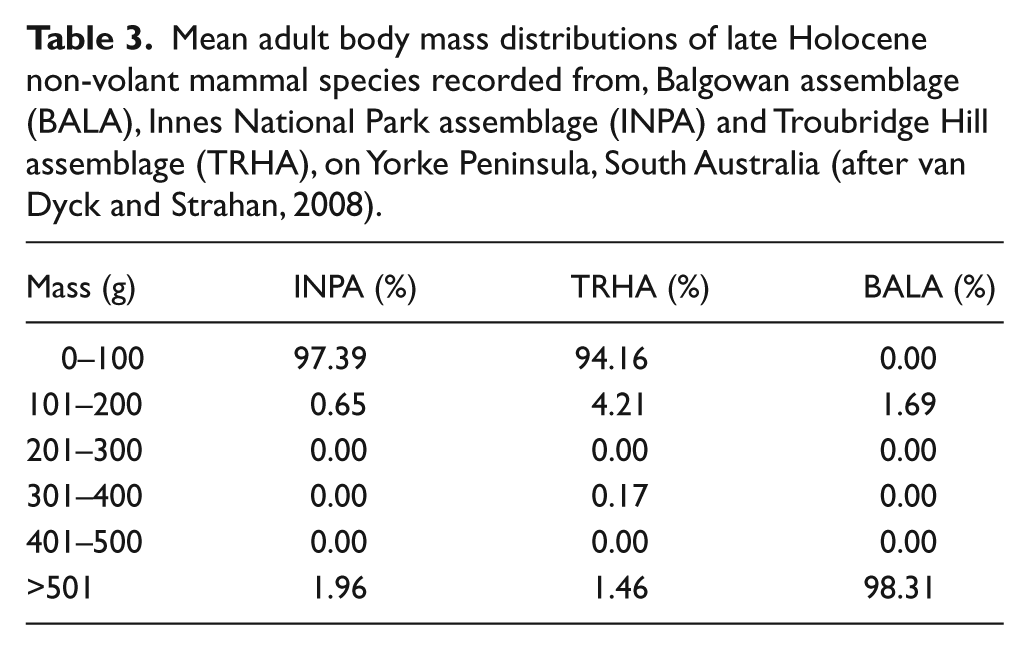
Thirty-one mammal species represented by a minimum of 1314 individuals were recovered from the three Holocene fossil assemblages examined (Table 2). Collections from both INPA and TRHA were composed predominately (>98%) of mammals weighing 200 g or less (Table 3). Mammals that weigh more than 200 g were rare and all represented by juveniles. In contrast, the collection from BALA was composed almost entirely of mammals that weighed more than 500 g (>98%; Table 3). The species compositions from the three assemblages also differed dramatically. Only two species (Isoodon obesulus and Perameles bougainville) were recorded from all three assemblages and only one additional species (Macrotis lagotis) occurred in both the BALA and TRHA. No other species occurred in both INPA and BALA. Many species occur in both the INPA and TRHA, but with the exception of Notomys mitchellii, these species are typically rare in either one or both of the assemblages. INPA is dominated by Sminthopsis griseoventer, N. mitchellii, Pseudomys occidentalis and R. fuscipes, TRHA is dominated by Pseudomys australis and to a lesser extent N. mitchellii and R. fuscipes, and BALA is dominated by macropodids (kangaroos and kin). This is reflected in the richness and evenness of the assemblages (Table 2). BALA was slightly less rich than the other two assemblages but richness did not differ greatly. However, Buzas-Gibson Evenness indicated that BALA had the most evenly distributed species abundance. TRHA had the least evenly distributed species whilst species from the INPA were moderately evenly distributed.
Mean adult body mass distributions of late Holocene non-volant mammal species recorded from, Balgowan assemblage (BALA), Innes National Park assemblage (INPA) and Troubridge Hill assemblage (TRHA), on Yorke Peninsula, South Australia (after van Dyck and Strahan, 2008).
Discussion
Systematics
Of the 31 species identified in this study, three were assigned to species with caution. Specimens identified as Sminthopsis sp. cf. S. dolichura (Table 2) were distinguished from S. griseoventer and S. aitkeni by the following characters: M3 metacristid shorter or subequal to, rather than longer than its crown length; dP3 has three major cusps rather than two; Cl crown taller rather than subequal or shorter than that of P1; very small entoconids on M2 and M3 (Kitchener et al., 1984). Differentiating between fragmentary specimens of S. dolichura and S. murina is much more difficult. Maxillae appear morphologically indistinguishable and lower molars of both species have very small or absent entoconids. In addition, fossil specimens rarely retain all of their teeth, so key diagnostic features are not always preserved. As S. griseoventer occurs sympatrically with S. dolichura on Eyre Peninsula today (Kemper et al., 2010) and S. murina has never been recorded west of the Flinders Lofty Block, these specimens probably represent S. dolichura rather than S. murina, but cannot be diagnosed with absolute confidence.
Yorke Peninsula lies well outside the known distribution of M. giganteus, but fragmentary remains of M. fuliginosus appear morphologically indistinguishable from M. giganteus. Therefore, specimens from Yorke Peninsula are referred to Macropus sp. cf. M. fuliginosus reflecting the species that presently occurs in the area. For similar reasons the specific identity of the Pseudocheirus specimen is also unclear. Fragmentary crania of Pseudocheirus occidentalis and Pseudocheirus peregrinus appear to be indistinguishable (Baverstock et al., 1990). On the basis of then-unpublished research, Ride (1970) synonymized several named forms of Pseudocheirus, including Pseudocheirus occidentalis, with P. peregrinus. Although early molecular systematic data indicated considerable complexity within Ride’s concept of P. peregrinus, McKay (1988) maintained the synonymy. However, Pseudocheirus occidentalis and P. peregrinus are now considered separate species (Burbidge and de Tores, 1998; de Tores, 2008). As the study area is closer to the known range of P. peregrinus than that of Pseudocheirus occidentalis, the taxon is referred to here as Pseudocheirus sp. cf. peregrinus.
If the tentatively made species identifications (Table 2) are correct, then, with the exceptions of L. latifrons, Lagorchestes leporides and P. australis, the native non-volant mammals recorded from Yorke Peninsula all consist of species that are either currently considered to be conspecific with populations recorded from southwestern Australia, or have been considered so in the past. The Australian arid zone cuts the late-Holocene southern coast where the limestone karst Nullarbor Plain reaches the coast at the Head of the Great Australian Bight in far western South Australia. This has long been identified as a potential barrier between eastern and western biotas (e.g. Main et al., 1958; Parsons, 1969). The extinction of many and in some cases all of the relevant populations of many mammal species since European colonization has rendered investigation of these east–west relationships much more difficult.
Accumulation agents
The INPA and TRHA were both found in shallow caves in coastal cliffs and were predominately composed of small mammals that weighed <200 g (Table 3), characteristics that are indicative of tytonid owl accumulations (Andrews, 1990; Baird, 1991; Higgins, 1999). However, the BALA was collected from a surface lag deposit on a deflated sand dune. Its fauna is composed entirely of species that weighed >200 g (Table 3), the remains of which were associated with charcoal indicative of an Aboriginal midden (Coutts, 1966). As different accumulation agents necessarily impose different biases on the assemblages they produce, quantitative palaeoecological comparison between owl assemblages (INPA; TRHA) and Aboriginal midden assemblages (BALA) cannot be made. However, contrasting assemblages deposited by different accumulation agents may provide a more complete appreciation of Yorke Peninsula’s Holocene mammal fauna. Differing accumulation agents may explain compositional differences between BALA and the other two assemblages, but not differences in the composition of INPA and TRHA. Avenant (2005) showed that frequencies of barn owl prey in South Africa closely matched the frequencies of prey available in the surrounding small animal community. In addition, studies have shown that Australian barn owls, which are typically nomadic and follow rodent irruptions, can have considerable dietary flexibility and are capable of consuming a diet consisting primarily of marsupials (Heywood and Pavey, 2002) or even reptiles (McDowell and Medlin, 2009). Therefore, the differences of species presence and frequency observed between INPA and TRHA are likely to represent differences in the proximal living communities at the time(s) of predation.
Assemblage composition and diversity
Our analysis of the INPA reveals that several now locally extinct mammal species persisted on southern Yorke Peninsula until European settlement (Table 2). It is also possible that species recorded from TRHA persisted until European settlement. Species which had not previously been reported from the study area include Phascogale calura, S. griseoventer, Potorous platyops, Leporillus apicalis, Pseudocheirus sp. cf. P. peregrinus and Lagostrophus fasciatus. The last two represent new records for the entire Yorke Peninsula.
Whilst the INPA and TRHA share many species in common (Table 2), S. griseoventer, I. obesulus, C. concinnus, P. platyops, N. mitchellii, Pseudomys occidentalis, R. fuscipes and Rattus tunneyi are either unique to, or more abundant in, the INPA. In contrast, Dasyurus geoffroii, M. lagotis, Pseudocheirus sp. cf. P. peregrinus, L. apicalis and P. australis are either unique to, or much more abundant in the TRHA.
The species that characterize the INPA either require higher precipitation, sandy substrates or prefer mallee with heath understorey vegetation. Brandle (2010) found that S. griseoventer occurred in a range of mallee communities with a shrubby understorey of Melaleuca species on a variety of soils with light calcareous strew cover. This species has also been detected on sandy substrates (Crowther et al., 1999) and in heath woodland on coastal plains (Dickman, 1988). Isoodon obesulus is known to occupy a number of vegetation communities. However, they generally include a combination of sandy soil and dense heath (Paull, 2008). The nest sites of C. concinnus have been reported to be associated with sandy substrates (Morrant and Petit, 2011) and E. diversifolia, E. rugosa, and Melaleuca spp. are considered important sources of dietary nectar and pollen (Pestell, 2005; Pestell and Petit, 2007). The occurrence of C. concinnus in the fossil record on southern Yorke Peninsula suggests that, unlike southern Eyre Peninsula (McDowell and Medlin, 2010), mallee communities were well-established components of the pre-European vegetation rather than the result of European disturbance. Notomys mitchellii is also typically associated with sandy habitats (Brandle, 2010; Watts and Aslin, 1981) and mallee areas (Robinson et al., 2000b). Rattus fuscipes, though best considered an ecological generalist, is frequently found in coastal heathlands or forests with a heath understorey, but has also been detected on limestone and sand islands that support a chenopod shrubland (Robinson et al., 2000b).
Pseudomys occidentalis is a poorly understood species that was widely distributed across southern Australia prior to European colonization (Whisson, 2008), but never collected alive in South Australia. Some researchers have suggested that the species was in decline before the arrival of Europeans (Lee, 1995; Watts and Aslin, 1981), but its abundant occurrence in an assemblage radiometrically dated to the approximate time of European colonization of Australia shows otherwise. Pseudomys occidentalis appears to be highly sensitive to fire and apparently prefers patches of extremely dense, long unburnt (between 30 and 50 yr) vegetation on sandy clay loam or sandy loam (Whisson, 2008). It is typically assumed that prior to European colonization fires were managed by Aborigines. The abundance of Pseudomys occidentalis in the INPA suggests that the area was infrequently burnt or burnt in patches. Near absence of the species from the TRHA probably reflects its preference for dense vegetation, rather than more frequent fire.
Pseudomys australis is by far the most abundant species in the TRHA, in which it contributes >50% of the relative abundance (Table 2). The fossil distribution of this species indicates that it was much more widely distributed in the past but is now restricted to gibber with cracking clay areas in the Lake Eyre Basin (Brandle et al., 1999; Robinson et al., 2000b). These environmental conditions were probably replicated on southern Yorke Peninsula by areas of exposed calcrete associated with ephemeral lakes that would provide the gypsum-rich cracking clays that the species uses for shelter and nesting. However, the coastal form attributed to this species in the Yorke Peninsula assemblages is somewhat larger than the extant desert form and may represent either an example of Bergmann’s rule or potentially a hitherto unrecognized distinct species.
Other species that distinguish the fauna of TRHA from INPA include D. geoffroii, M. lagotis, Pseudocheirus sp. cf. P. peregrinus and L. apicalis. With the exception of Pseudocheirus sp. these species could once be found throughout the more arid areas of Australia but unlike D. geoffroii and M. lagotis, L. apicalis is not thought to have occurred on Yorke Peninsula post European settlement (van Dyck and Strahan, 2008). Leporillus apicalis are thought to have built their nests in hollow tree limbs, caves and rock overhangs (Baynes, 1987; Copley, 1999 and references therein). Leporillus apicalis occurred primarily on calcareous but occasionally gypseous and/or saline soils that supported perennial shrublands composed primarily of chenopods (e.g. Atriplex spp. and Maireana spp.) nitre-bush (Nitraria billardierei) and pigfaces (e.g. Carpobrotus spp.) (Copley, 1999; Medlin, 1993). These succulent plants are thought to have made up a large proportion of the diet of both Leporillus species (Copley, 1999). Therefore, L. apicalis may have been associated with the ephemeral salt lakes that occur in the study area (Croft, 2008). However, Douglas (1980) identified sandalwood and quandong (both Santalum spp.) seeds as an important dietary component in more arid parts of this rodent’s range.
Viewed together, the three assemblages investigated in this study yielded 27 species of native mammal, suggesting that the diversity of Yorke Peninsula’s pre-European fauna was comparable with the mammalian diversity found in other studies (e.g. Baynes, 1987; Copley et al., 1989; McDowell and Medlin, 2010; Watts and Ling, 1985). However, if considered individually, the richness of mammals yielded by each individual assemblage is considerably lower (Table 2). Whilst these assemblages show similar richness, their evenness varies considerably. The evenness values obtained from the INPA and BALA indicate that a larger number of species shared greater abundance. The very low evenness of TRHA is congruent with its extremely high abundance of P. australis. The very high relative abundance of P. australis may result from its propensity to irrupt during resource pulses following high rainfall, a phenomenon that barn owls are quick to exploit (McDowell and Medlin, 2009; Pavey et al., 2008). However, it is unlikely that the TRHA roost was used only during rodent irruptions and even though irruptions may periodically elevate the relative abundance of P. australis, it cannot explain why the species dominates the TRHA but is almost absent from the INPA. Pavey et al. (2008) demonstrated that barn owls are capable of selective predation and have a preference for larger rodents. However, assuming all owls are equally selective, the differences in the evenness of the TRHA and INPA cannot be explained by this phenomenon.
Factors that could influence the mammal richness, evenness and composition in a fossil assemblage include the accumulation agent (each with their inherent sampling biases), the prevailing climatic conditions experienced at the time of deposition and differences in local ecological constraints such as substrate, vegetation and fire frequency. However, it is most likely that differences in evenness reflect the levels of homogeneity of the ecosystems surrounding the assemblage locations.
Climatic conditions
Climate change is a driving force for faunal change and evolution (Barnosky, 2008; Barnosky and Bell, 2003; Hadly, 1996) and may affect the diversity and distribution of mammals on a century and perhaps even decadal scale. Dodson (2001) found little change in the terrestrial vegetation of Australia’s Mediterranean-type climate regions during the Holocene, implying that conditions have been relatively stable for much of the epoch. Numerous climate proxies indicate that 900 years ago, when the TRHA began accumulating, the Northern Hemisphere was experiencing the ‘Medieval Warm Period’, during which surface temperatures were equal to or greater than present (e.g. Jones et al., 2009; Mann et al., 2008; Nunn, 2007; Tyson and Lindesay, 1992). However, 200 years ago when the INPA was accumulating, the Northern Hemisphere was experiencing the tail end of the ‘Little Ice Age’, during which surface temperatures were cooler than present (Jones et al., 2009; Mann et al., 2008; Nunn, 2007; Tyson and Lindesay, 1992). Mann and Jones (2003) argued that even though fewer climate proxy records are available, surface temperatures in the Southern Hemisphere expressed a similar trend to the better-studied Northern Hemisphere record. While these periods correspond with the depositional times of TRHA and INPA respectively, the warmer and wetter period (Cobb et al., 2003) correlates with the more arid-adapted fauna of TRHA, suggesting little or no relationship between these global climate trends and local faunal composition.
The compositional differences between the INPA and TRHA correlate well with Yorke Peninsula’s historical rainfall distribution. The mean annual rainfall of western Yorke Peninsula, where the wetter-adapted fauna occurred, is 14% greater than that of eastern Yorke Peninsula, where the drier-adapted fauna occurred and may have influenced local ecological constraints enough to explain local differences in faunal composition.
Local ecological constraints
Small mammals typically have specific environmental requirements that dictate their distributions (e.g. Hadly, 1999). Many require a particular precipitation level and vegetation community for feeding, nesting or to provide protection from predation. Others, particularly burrowing species, are restricted to specific substrates (e.g. Robinson et al., 2000b). The clear species compositional differences between the INPA and TRHA might best be explained by the distinctly different substrates and associated vegetation that occurred on southwestern and southeastern Yorke Peninsula (Figure 1). As these vegetation communities represent pre-European conditions (Croft, 2008), relationships between fauna, flora and substrate can be inferred with confidence. TRHA and BALA share the same substrate and vegetation association (Dryland Tea-tree–Drooping Sheoak low open woodland on calcrete, see Figure 1). However, marked differences in faunal composition between the two assemblages add further support to the conclusion that they are the product of different accumulation agents.
Biogeography
Yorke Peninsula’s late-Holocene mammal fauna can be separated into four groups: (1) taxa that retain naturally occurring populations in both southeastern and southwestern Australia; (2) taxa that retain naturally occurring populations in both southeastern and southwestern Australia but whose taxonomic status has been unclear; (3) taxa that are locally extinct in southeastern Australia but still have naturally occurring populations in southwestern Australia; and (4) taxa that are extinct in both southeastern and southwestern Australia. Distribution maps in van Dyck and Strahan (2008) are utilized in the following discussion.
Sminthopsis dolichura and M. fuliginosus are examples of members of the Yorke Peninsula fauna that retain extant mainland populations both southwest and southeast of the Nullarbor. Tachyglossus aculeatus, S. crassicaudata, S. griseoventer, I. obesulus, T. vulpecula, C. concinnus, N. mitchellii, P. shortridgei and R. fuscipes also share this distribution pattern. Because of conservation concerns the distribution of P. shortridgei has been well studied. Cooper et al. (2003) found little genetic difference between southwestern and southeastern populations, but Salinas et al. (2009) found that Victorian and southwestern Australian specimens showed differentiation comparable with different species. They inferred that the two populations were isolated about 1.4 million years ago by aridity on the Nullarbor Plain. Although P. shortridgei was not recorded from surfaces of southern Nullarbor caves east of the western end of the Great Australian Bight by Baynes (1987), its remains are present in mid-Holocene levels of Allen’s Cave and Warbla Cave in the far west of South Australia (Baynes, 1994, personal observations; Kemper et al., 2010). During glacials continuous east–west populations may have existed in coastal dunes across what is now the Bight (Cooper et al., 2003). Little is known of the South Australian distribution of P. shortridgei between the mid-west and southeast (Kemper et al., 2010). It is possible that the major genetic discontinuity occurred across the Murray River valley and estuary where suitable heath habitats probably do not exist, rather than across the Nullarbor, although major genetic discontinuities may have occurred in both areas. Neaves et al. (2009) have demonstrated that such genetic discontinuities are unexpectedly common even in large vagile Australian mammals such as kangaroos. Remains of P. shortridgei are plentiful in South Australian assemblages, such as those on Yorke Peninsula, so this question should be resolvable using ancient DNA techniques. It would also be interesting to explore the affinities of the Kangaroo Island population, particularly if it is still extant (Kemper et al., 2010).
The genus Pseudocheirus also retains southwestern and southeastern populations that were once considered conspecific but have since been recognized as separate species. Pseudocheirus remains were recorded in surface material from three caves along the southern edge of the western Nullarbor, which Baynes (1987) reported under the then-current name of P. peregrinus. Pseudocheirus has also been recorded from late-Holocene strata in Allen’s Cave in the far west of South Australia (Baynes, 1994), and from a Holocene site on southern Eyre Peninsula (McDowell and Medlin, 2010; Table 4).
Native non-volant mammal faunas of Yorke and Eyre Peninsulas, South Australia. Eyre Peninsula is divided into a southwestern zone (south and west of the 400 mm mean annual rainfall isohyet, Schwerdtfeger, 1985), comparable in area and rainfall range with Yorke Peninsula, and a northern and eastern zone of lower rainfall. Late-Holocene and historic Eyre Peninsula mammal records from Watts and Ling (1985) and Baynes (1987) with modifications, McNamara (1997), McDowell and Medlin (2010) (southwestern zone only) and Kemper et al. (2010). Vernacular names from van Dyck and Strahan (2008). a
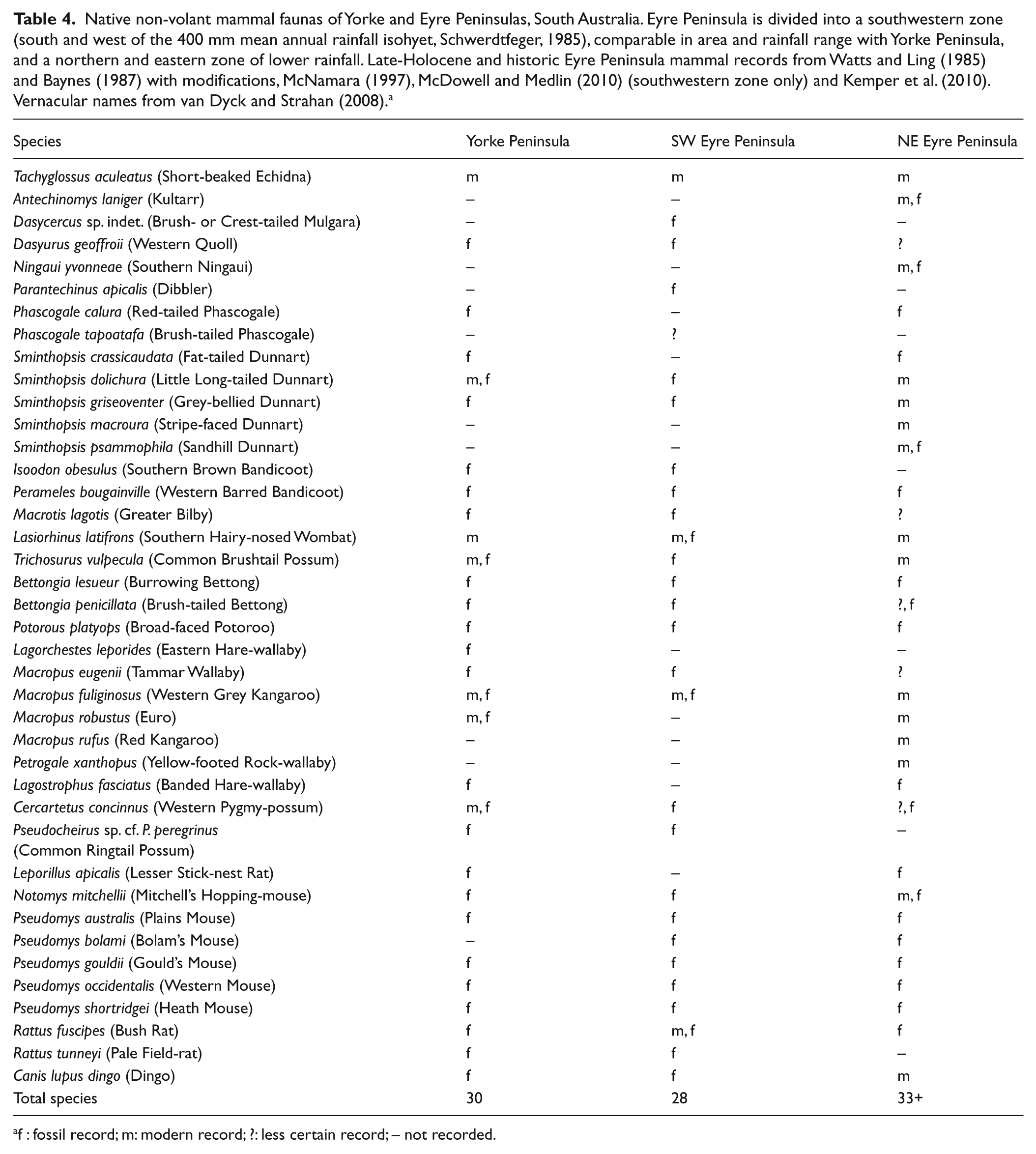
f : fossil record; m: modern record; ?: less certain record; – not recorded.
Dasyurus geoffroii, P. calura, Bettongia penicillata and Pseudomys occidentalis comprise a third species group whose naturally occurring mainland populations persist only in the southwest, having become extinct in southeastern Australia. Macropus eugenii shows a variation on this pattern, surviving on Kangaroo Island in South Australia.
The final group consists of species in which both southeastern and southwestern mainland populations are extinct (though some survive on islands or in northern Australia). These species include P. bougainville, M. lagotis, Bettongia lesueur, P. platyops, L. fasciatus, L. apicalis, P. gouldii (encompassing P. fieldi) and R. tunneyi. Of these, L. fasciatus has long been known from South Australian mid-Holocene deposits (e.g. McNamara, 1997) and was recently shown to have been extant and widespread, though subspecifically distinct, in South Australia when colonized by Europeans (Helgen and Flannery, 2003).
The pre-European fauna of Yorke Peninsula is highly comparable in both species composition and diversity with that recorded from southwestern Eyre Peninsula (Table 4). Both regions are similar in area and climate (DEH, 2003; Schwerdtfeger, 1985) and following Holocene sea-level rise would have formed virtual islands for terrestrial fauna, isolated by the Southern Ocean to the south and aridity to the north. The 27 species recorded from southwestern Eyre Peninsula include two dasyurids not recorded from Yorke Peninsula: Dasycercus sp. indet., widespread in sandy habitats in the arid zone, and Parantechinus apicalis, at its most easterly known occurrence. Pseudomys bolami is also recorded from Eyre Peninsula but not Yorke Peninsula. Mammals recorded from Yorke Peninsula but not southwestern Eyre Peninsula include P. calura, L. leporides, L. fasciatus and L. apicalis. This is the most westerly occurrence of L. leporides known; the other three species have been recorded from northern Eyre Peninsula (Table 4). An additional six species were recorded from northern Eyre Peninsula only (Table 4), though all were once widespread in the arid zone.
Conclusions
The faunal compositions of the three Holocene fossil mammal assemblages we investigated are distinctly different. The differences between the BALA fauna and those of the more southerly assemblages are most likely due to differing accumulation agents. Evidence suggests that BALA was accumulated by humans, but that INPA and TRHA were accumulated by owls. Whilst assemblages accumulated by different accumulation agents may not be quantitatively comparable, they form an ensemble that provides a more complete representation of a region’s fauna than can be gained from any single site.
The segregation of small mammal species on southern Yorke Peninsula was probably driven by local ecological constraints. Species that show preferences for mallee, sandy substrates and dense vegetation occur predominately or exclusively in the INPA. In contrast, many of the species that occur predominately or exclusively in the TRHA show preferences for stony, loamy and cracking clay substrates and more open woodland with a shrubland and grassland understorey. The high degree of habitat fidelity to known ecological constraints or preferences shown by the Yorke Peninsula fossil faunas provides support for the assumption of uniformitarianism that palaeoecologists are typically forced to make. However, given the close proximity of the INPA and TRHA and their extensive composition differences, this study also demonstrates the risk of geographically over-extrapolating palaeoecological data, particularly in regions of heterogeneous habitat.
It is clear that before the arrival of Europeans the native non-volant mammal fauna of Yorke Peninsula was much more diverse than that of today. By establishing the age of the analyzed assemblages we demonstrate that climate change played little or no part in the extirpation or extinction of these mammals. Europeans are probably responsible through habitat destruction, introduction of exotic competitors, predators and/or hunting. However, more data are needed before we can determine the relative impacts of each mechanism.
This research highlights the potential Holocene fossil assemblages have for providing baseline data to natural resource managers, informing on the composition, richness, evenness and heterogeneity of pre-European faunas. In addition, studies of this type are valuable to conservation managers because they identify species that may be re-introduced to a site as part of ecosystem restoration programmes. Investigation of the biogeography of species identified in this study show strong faunal affinities between Yorke Peninsula and southwestern Eyre Peninsula. This suggests that prior to Holocene sea-level rise mammal populations were continuous between the peninsulas and across the intervening lower-lying area that is now Spencer Gulf.
AMS radiocarbon dating established that the INPA and TRHA were of late-Holocene age and suffered little from the effects of time averaging. Whilst this research supports the frequently made assumption that surface and shallow unstratified deposits are typically of late-Holocene origin, it is highly desirable that future research includes radiometric dating to avoid comparison of differently aged assemblages. The Bridgewater Formation undoubtedly preserves numerous yet-to-be-discovered Holocene fossil vertebrate assemblages. As more are recovered, analyzed and dated, the ensuing composite fossil record may reveal the pressures that dictated the biogeography of Australia’s pre-European fauna, and may provide insight into the processes that must be managed to preserve our remaining biodiversity.
Footnotes
Acknowledgements
We thank C Kemper, P Rismiller, L Jansen and M Ayre for collecting the INPA; A Camens and T Worthy for collecting the initial TRHA sample; C Kemper and D Stemmer for providing access to specimens in the SAM Mammal and Subfossil Collections; C Kemper for advice on mammal distributions; M Binnie for providing access to specimens in the SAM Palaeontology Collection and R Correll and A Macken for useful discussion of draft manuscripts. The quality of the paper was further enhanced by the constructive comments of two anonymous referees.
Funding
This work was supported by Nature Foundation SA.